
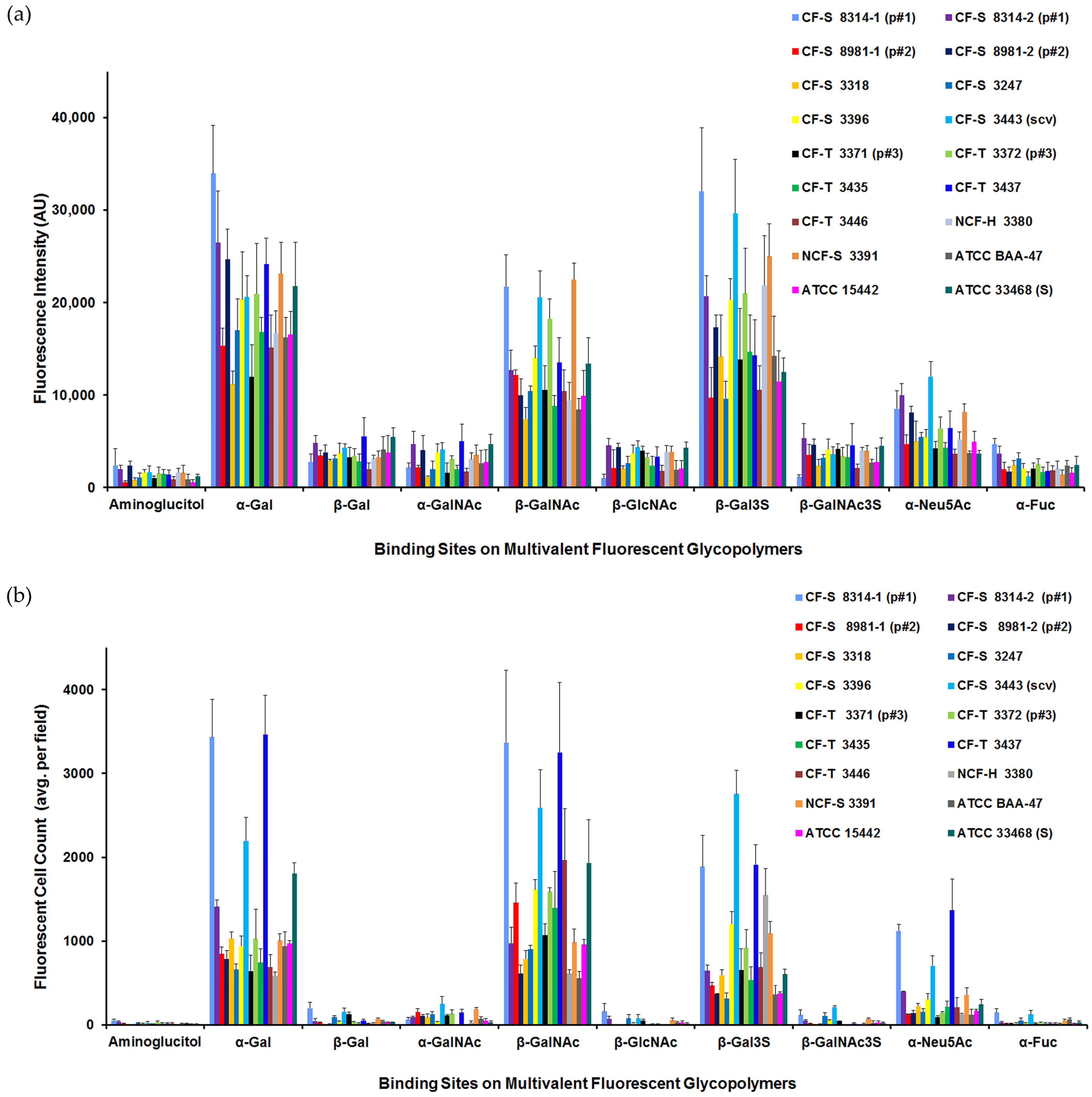
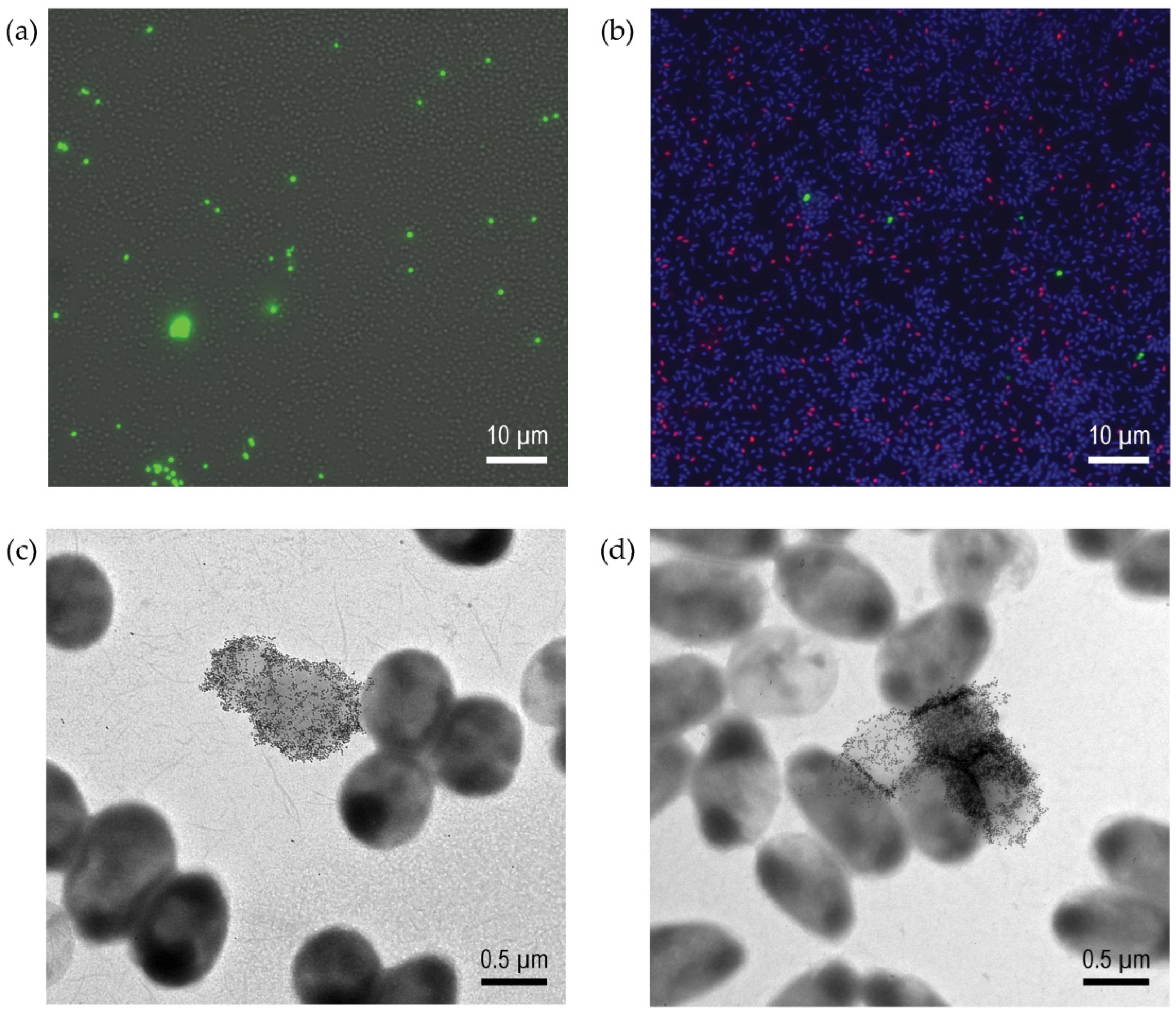
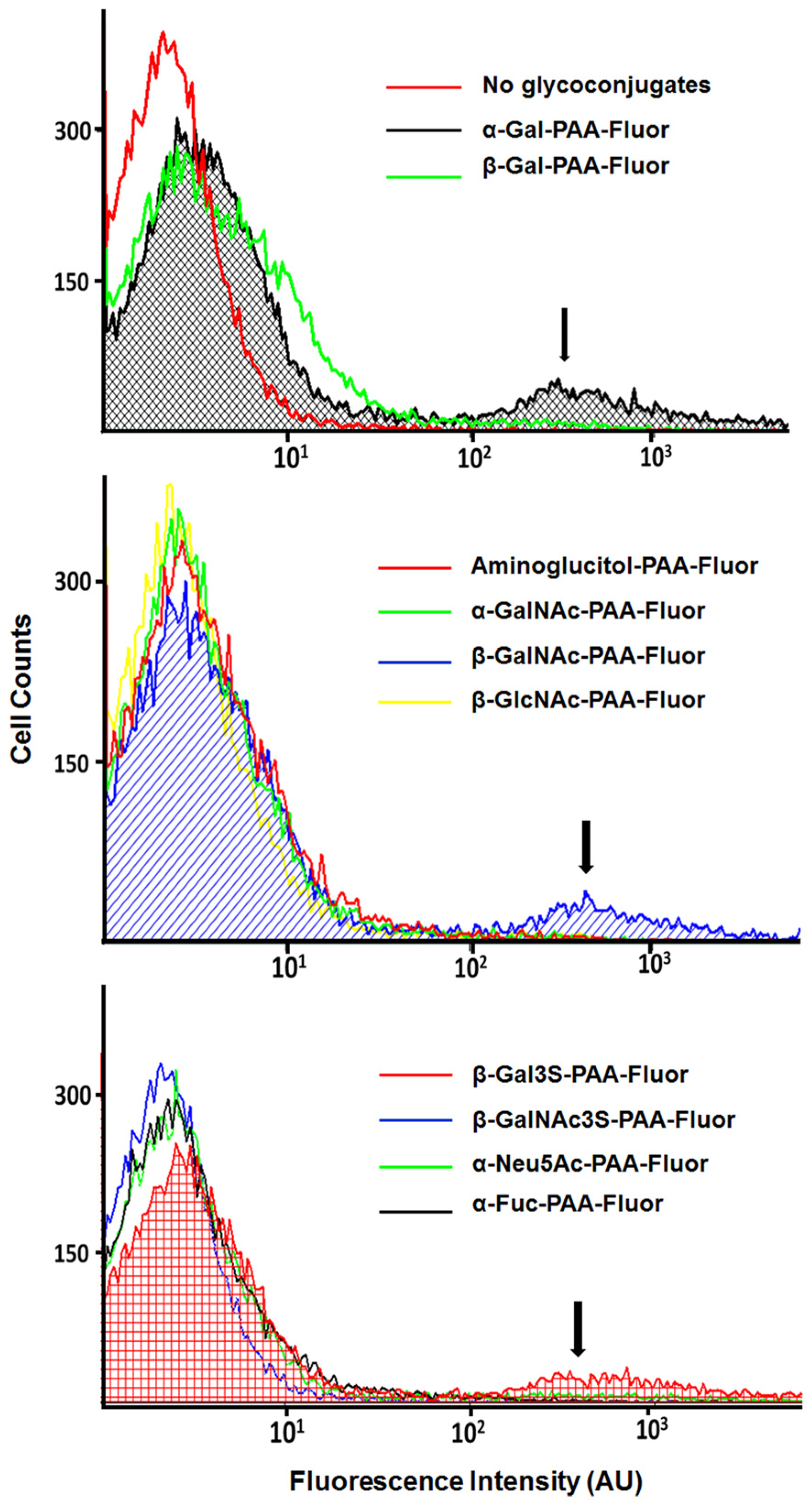
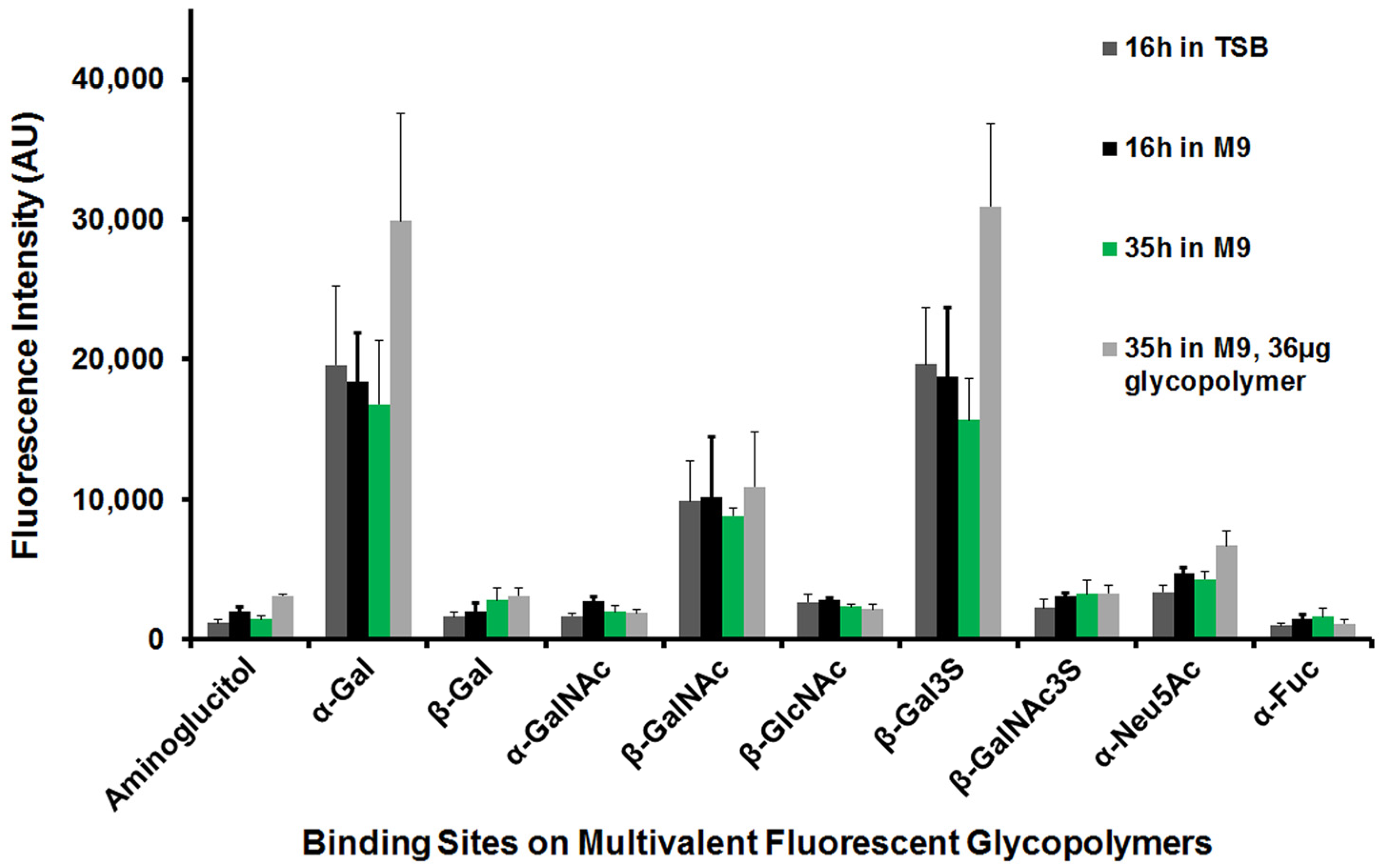
Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Chance, D.L.; Wang, W.; Waters, J.K.; Mawhinney, T.P. Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides. Microorganisms 2024, 12, 801. https://doi.org/10.3390/microorganisms12040801
Chance DL, Wang W, Waters JK, Mawhinney TP. Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides. Microorganisms. 2024; 12(4):801. https://doi.org/10.3390/microorganisms12040801
Chicago/Turabian StyleChance, Deborah L., Wei Wang, James K. Waters, and Thomas P. Mawhinney. 2024. "Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides" Microorganisms 12, no. 4: 801. https://doi.org/10.3390/microorganisms12040801
APA StyleChance, D. L., Wang, W., Waters, J. K., & Mawhinney, T. P. (2024). Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides. Microorganisms, 12(4), 801. https://doi.org/10.3390/microorganisms12040801