
Molecular and Serological Detection of Vector-Borne Pathogens Responsible for Equine Piroplasmosis in Europe between 2008 and 2021

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
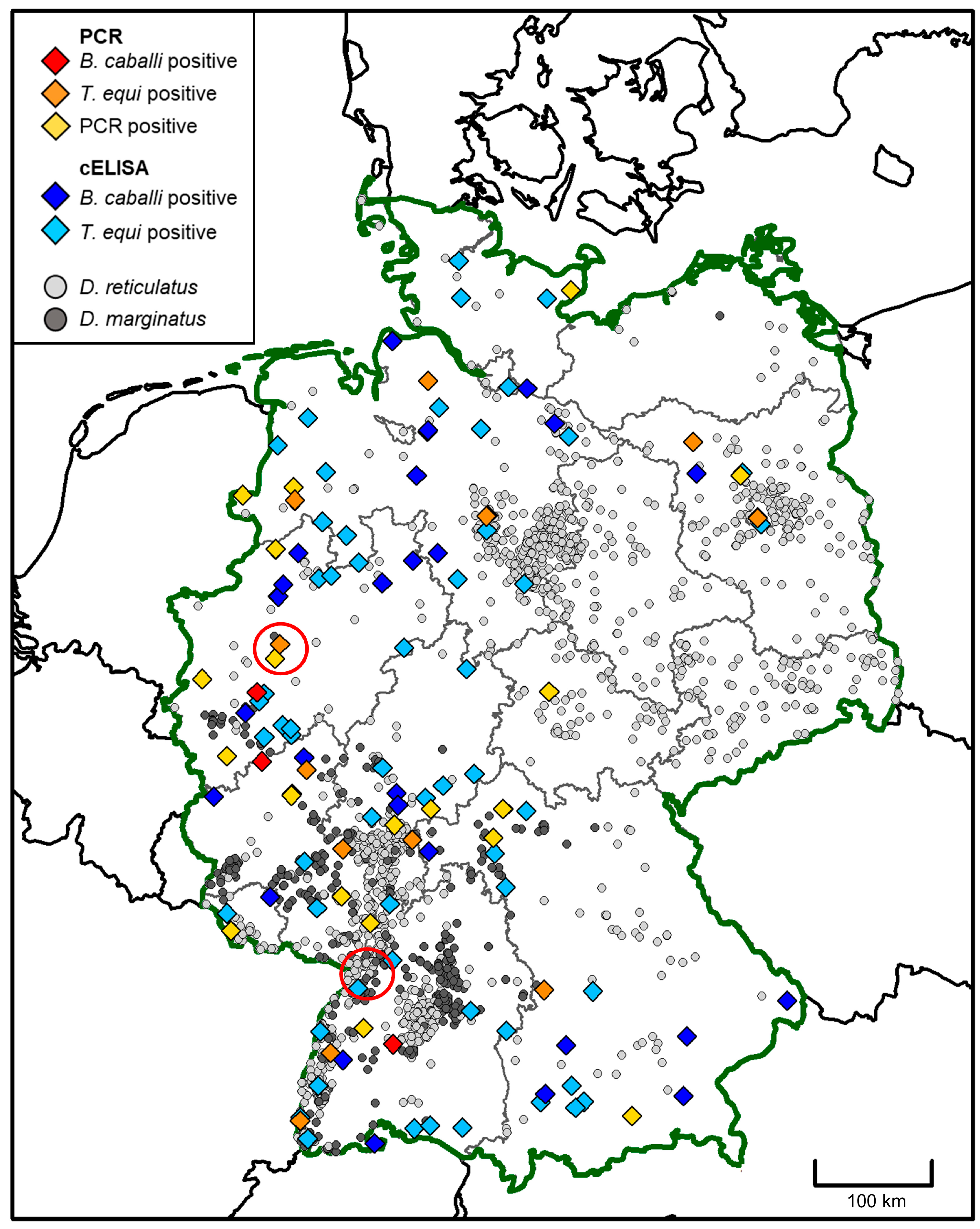
3. Results
3.1. Molecular and Serological Testing for EP
3.2. Signalement
3.3. Regionality, Seasonality, and Years of Testing
3.4. Geographic Origin of Affected Horses in Germany from 2008 to 2021
4. Discussion
4.1. Analysis of Risk Factors
4.1.1. Regionality
4.1.2. Age
4.1.3. Sex
4.1.4. Breed
4.2. Endemisation in Germany
4.3. Diagnostic Assays
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qablan, M.A.; Obornik, M.; Petrzelkova, K.J.; Sloboda, M.; Shudiefat, M.F.; Horin, P.; Lukes, J.; Modry, D. Infections by Babesia caballi and Theileria equi in Jordanian equids: Epidemiology and genetic diversity. Parasitology 2013, 140, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- De Waal, D.T. Equine piroplasmosis: A review. Br. Vet. J. 1992, 148, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Bartolome Del Pino, L.E.; Meana, A.; Zini, M.; Cersini, A. Evidence of transplacental transmission of equine piroplasms Theileria equi and Babesia caballi in an Italian breed mare. Folia Parasitol. 2023, 70, 005. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, M.T.; Lewis, B.D.; Penzhorn, B.L. Molecular evidence for transplacental transmission of Theileria equi from carrier mares to their apparently healthy foals. Vet. Parasitol. 2007, 148, 130–136. [Google Scholar] [CrossRef] [PubMed]
- De Waal, D.T.; van Heerden, J. Infectious Diseases of Lifestock; Oxford University Press: Cape Town, South Africa, 2004. [Google Scholar]
- Sant, C.; d’Abadie, R.; Pargass, I.; Basu, A.K.; Asgarali, Z.; Charles, R.A.; Georges, K.C. Prospective study investigating transplacental transmission of equine piroplasmosis in thoroughbred foals in Trinidad. Vet. Parasitol. 2016, 226, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Levy, S.; Gottlieb, Y.; Fry, L.M.; Knowles, D.P.; Steinman, A. Twenty years of Equine Piroplasmosis research: Global distribution, molecular diagnosis, and phylogeny. Pathogens 2020, 9, 926. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Pelzel-McCluskey, A.M.; Mealey, R.H.; Knowles, D.P. Equine piroplasmosis. Vet. Clin. N. Am. Equine Pract. 2014, 30, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Sumbria, D.; Moudgil, A.D.; Singla, L.D. Equine piroplasmosis: Current status. Veterinaria 2014, 1, 9–14. [Google Scholar]
- Stiller, D.; Coan, M.E. Recent developments in elucidating tick vector relationships for anaplasmosis and equine piroplasmosis. Vet. Parasitol. 1995, 57, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Ehrmann, C.; Lembcke, M.; Roscher, K.; Strube, C. Theileria equi-infection in 2 German horses returning from a trail ride in southern France. Tierarztl. Prax. Ausg. G Grosstiere Nutztiere 2020, 48, 124–129. (In German) [Google Scholar] [CrossRef] [PubMed]
- Scoles, G.A.; Ueti, M.W. Vector ecology of equine piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Salim, B.; Bakheit, M.A.; Kamau, J.; Sugimoto, C. Current status of equine piroplasmosis in the Sudan. Infect. Genet. Evol. 2013, 16, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, C.M. Equine piroplasmosis. J. Equine Vet. Sci. 2013, 33, 497–508. [Google Scholar] [CrossRef]
- Brüning, A. Equine piroplasmosis an update on diagnosis, treatment and prevention. Br. Vet. J. 1996, 152, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.M.; Sloet van Oldruitenborgh-Oosterbaan, M.M.; Stout, T.A.; van der Kolk, J.H.; Wollenberg, L.; Nielen, M.; Jongejan, F.; Werners, A.H.; Houwers, D.J. Prevalence of the causative agents of equine piroplasmosis in the South West of The Netherlands and the identification of two autochthonous clinical Theileria equi infections. Vet. J. 2012, 193, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Teodorowski, O.; Kalinowski, M.; Winiarczyk, D.; Janecki, R.; Winiarczyk, S.; Adaszek, L. Molecular surveillance of tick-borne diseases affecting horses in Poland-Own observations. Vet. Med. Sci. 2021, 7, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Tanczos, B.; Gyurkovszky, M.; Foldvari, G.; Solymosi, N.; Edelhofer, R.; Hornok, S. Serological and molecular detection of Theileria equi infection in horses in Hungary. Vet. Parasitol. 2013, 192, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Gallusova, M.; Qablan, M.A.; D’Amico, G.; Obornik, M.; Petrzelkova, K.J.; Mihalca, A.D.; Modry, D. Piroplasms in feral and domestic equines in rural areas of the Danube Delta, Romania, with survey of dogs as a possible reservoir. Vet. Parasitol. 2014, 206, 287–292. [Google Scholar] [CrossRef]
- Davitkov, D.; Vucicevic, M.; Stevanovic, J.; Krstic, V.; Slijepcevic, D.; Glavinic, U.; Stanimirovic, Z. Molecular detection and prevalence of Theileria equi and Babesia caballi in horses of central Balkan. Acta Parasitol. 2016, 61, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Gotić, J. Clinical and Serological Diagnostics and Molecular Typing of Equine Piroplasmosis Aetiological Agents in the Republic of Croatia. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 2015. [Google Scholar]
- Bartolome Del Pino, L.E.; Nardini, R.; Veneziano, V.; Iacoponi, F.; Cersini, A.; Autorino, G.L.; Buono, F.; Scicluna, M. Babesia caballi and Theileria equi infections in horses in Central-Southern Italy: Sero-molecular survey and associated risk factors. Ticks Tick Borne Dis. 2016, 7, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Pikalo, J.; Sattler, T.; Eichinger, M.; Loitsch, A.; Schmoll, F.; Schusser, G.F. Seroprevalence of Babesia caballi and Theileria equi in horses in Central Germany. Pferdeheilkunde 2016, 32, 254–259. [Google Scholar] [CrossRef][Green Version]
- Sigg, L.; Gerber, V.; Gottstein, B.; Doherr, M.G.; Frey, C.F. Seroprevalence of Babesia caballi and Theileria equi in the Swiss horse population. Parasitol. Int. 2010, 59, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Graham, H.; van Kalsbeek, P.; van der Goot, J.; Koene, M.G.J. Low seroprevalence of equine piroplasmosis in horses exported from the Netherlands between 2015 and 2021. Front. Vet. Sci. 2022, 9, 954046. [Google Scholar] [CrossRef] [PubMed]
- Belkova, T.; Bartova, E.; Ricarova, D.; Jahn, P.; Jandova, V.; Modry, D.; Hrazdilova, K.; Sedlak, K. Theileria equi and Babesia caballi in horses in the Czech Republic. Acta Trop. 2021, 221, 105993. [Google Scholar] [CrossRef]
- Coultous, R.M.; Phipps, P.; Dalley, C.; Lewis, J.; Hammond, T.A.; Shiels, B.R.; Weir, W.; Sutton, D.G.M. Equine piroplasmosis status in the UK: An assessment of laboratory diagnostic submissions and techniques. Vet. Rec. 2019, 184, 95. [Google Scholar] [CrossRef] [PubMed]
- Coultous, R.M.; Leadon, D.P.; Shiels, B.R.; Sutton, D.; Weir, W. Investigating the presence of equine piroplasmosis in Ireland. Vet. Rec. 2020, 187, e97. [Google Scholar] [CrossRef] [PubMed]
- Kouam, M.K.; Kantzoura, V.; Gajadhar, A.A.; Theis, J.H.; Papadopoulos, E.; Theodoropoulos, G. Seroprevalence of equine piroplasms and host-related factors associated with infection in Greece. Vet. Parasitol. 2010, 169, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Montes Cortes, M.G.; Fernandez-Garcia, J.L.; Habela Martinez-Estellez, M.A. Seroprevalence of Theileria equi and Babesia caballi in horses in Spain. Parasite 2017, 24, 14. [Google Scholar] [CrossRef] [PubMed]
- Giubega, S.; Ilie, M.S.; Luca, I.; Florea, T.; Dreghiciu, C.; Oprescu, I.; Morariu, S.; Darabus, G. Seroprevalence of anti-Theileria equi antibodies in horses from three geographically distinct areas of Romania. Pathogens 2022, 11, 669. [Google Scholar] [CrossRef] [PubMed]
- Nadal, C.; Marsot, M.; Le Metayer, G.; Boireau, P.; Guillot, J.; Bonnet, S.I. Spatial and temporal circulation of Babesia caballi and Theileria equi in France based on seven years of serological data. Pathogens 2022, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Dirks, E.; Werner, G.; Schwarz, B.; Trübenbach, L.; Schwendenwein, I.; Joachim, A.; Cavalleri, J. Equine Piroplasmosis in German-speaking countries–An underdiagnosed disease? Prakt. Tierarzt 2021, 102, 1078–1088. (In German) [Google Scholar] [CrossRef]
- Scheidemann, W.; Liebisch, G.; Liebisch, A.; Budde, K. Equine piroplasmosis-A case of an acute infection with Theileria equi (syn. Babesia equi) in Germany. Pferdeheilkunde 2003, 19, 16–20. (In German) [Google Scholar] [CrossRef][Green Version]
- Friedhoff, K.T.; Soule, C. An account on equine babesioses. Rev. Sci. Tech. 1996, 15, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. Int. J. Environ. Res. Public Health 2019, 16, 1736. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of equine piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Bonfini, B.; Semproni, G.; Savini, G. Use of an in vitro culture system to detect Theileria equi strains from infected equids and/or reservoirs. Vet. Ital. 2006, 42, 217–223, 209–215. [Google Scholar] [PubMed]
- Mans, B.J.; Pienaar, R.; Latif, A.A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Zweygarth, E.; Josemans, A.I. L-cysteine replaces microaerophilous culture conditions for the in vitro initiation of Theileria equi. Parasitol. Res. 2014, 113, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Zweygarth, E.; Lopez-Rebollar, L.M.; Nurton, J.; Guthrie, A.J. Culture, isolation and propagation of Babesia caballi from naturally infected horses. Parasitol. Res. 2002, 88, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Weiland, G. Species-specific serodiagnosis of equine piroplasma infections by means of complement fixation test (CFT), immunofluorescence (IIF), and enzyme-linked immunosorbent assay (ELISA). Vet. Parasitol. 1986, 20, 43–48. [Google Scholar] [CrossRef]
- Wise, L.N.; Kappmeyer, L.S.; Silva, M.G.; White, S.N.; Grause, J.F.; Knowles, D.P. Verification of post-chemotherapeutic clearance of Theileria equi through concordance of nested PCR and immunoblot. Ticks Tick Borne Dis. 2018, 9, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Fritz, C.L. Emerging tick-borne diseases. Vet. Clin. N. Am. Small Anim. Pract. 2009, 39, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Camino, E.; Pozo, P.; Dorrego, A.; Carvajal, K.A.; Buendia, A.; Gonzalez, S.; de Juan, L.; Dominguez, L.; Cruz-Lopez, F. Importance of equine piroplasmosis antibody presence in Spanish horses prior to export. Ticks Tick Borne Dis. 2020, 11, 101329. [Google Scholar] [CrossRef] [PubMed]
- Zahler, M.; Schein, E.; Rinder, H.; Gothe, R. Characteristic genotypes discriminate between Babesia canis isolates of differing vector specificity and pathogenicity to dogs. Parasitol. Res. 1998, 84, 544–548. [Google Scholar] [CrossRef] [PubMed]
- South, A. rworldmap: A New R package for Mapping Global Data. R J. 2011, 3, 35–43. [Google Scholar] [CrossRef]
- Drehmann, M.; Springer, A.; Lindau, A.; Fachet, K.; Mai, S.; Thoma, D.; Schneider, C.R.; Chitimia-Dobler, L.; Broker, M.; Dobler, G.; et al. The spatial distribution of Dermacentor ticks (Ixodidae) in Germany-Evidence of a continuing spread of Dermacentor reticulatus. Front. Vet. Sci. 2020, 7, 578220. [Google Scholar] [CrossRef]
- Springer, A.; Lindau, A.; Probst, J.; Drehmann, M.; Fachet, K.; Thoma, D.; Rose Vineer, H.; Noll, M.; Dobler, G.; Mackenstedt, U.; et al. Update and prognosis of Dermacentor distribution in Germany: Nationwide occurrence of Dermacentor reticulatus. Front. Vet. Sci. 2022, 9, 1044597. [Google Scholar] [CrossRef]
- Ruegg, S.R.; Torgerson, P.; Deplazes, P.; Mathis, A. Age-dependent dynamics of Theileria equi and Babesia caballi infections in southwest Mongolia based on IFAT and/or PCR prevalence data from domestic horses and ticks. Parasitology 2007, 134, 939–947. [Google Scholar] [CrossRef]
- Laus, F.; Spaterna, A.; Faillace, V.; Veronesi, F.; Ravagnan, S.; Beribe, F.; Cerquetella, M.; Meligrana, M.; Tesei, B. Clinical investigation on Theileria equi and Babesia caballi infections in Italian donkeys. BMC Vet. Res. 2015, 11, 100. [Google Scholar] [CrossRef]
- Laus, F.; Veronesi, F.; Passamonti, F.; Paggi, E.; Cerquetella, M.; Hyatt, D.; Tesei, B.; Fioretti, D.P. Prevalence of tick borne pathogens in horses from Italy. J. Vet. Med. Sci. 2013, 75, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Edelhofer, R.; Foldvari, G.; Joachim, A.; Farkas, R. Serological evidence for Babesia canis infection of horses and an endemic focus of B. caballi in Hungary. Acta Vet. Hung. 2007, 55, 491–500. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Idoko, I.S.; Edeh, R.E.; Adamu, A.M.; Machunga-Mambula, S.; Okubanjo, O.O.; Balogun, E.O.; Adamu, S.; Johnson, W.; Kappmeyer, L.; Mousel, M.; et al. Molecular and serological detection of piroplasms in horses from Nigeria. Pathogens 2021, 10, 508. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Mangili, V.; Salvatori, R.; Maresca, C.; Scoccia, E.; Torina, A.; Moretta, I.; Gabrielli, S.; Tampieri, M.P.; Pietrobelli, M. Prevalence and diagnosis of Babesia and Theileria infections in horses in Italy: A preliminary study. Vet. J. 2010, 184, 346–350. [Google Scholar] [CrossRef]
- Probst, J.; Springer, A.; Strube, C. Year-round tick exposure of dogs and cats in Germany and Austria: Results from a tick collection study. Parasites Vectors 2023, 16, 70. [Google Scholar] [CrossRef]
- Joachim, A.; Cavalleri, J.M.V.; Berger, S. Equine anaplasmosis and equine piroplasmosis in Germany, Austria and Switzerland–previously anecdotal, now relevant? Schweiz. Arch. Tierheilkd. 2022, 164, 35–50. (In German) [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.W.; Walker, W.; Alexander, J. Sex-associated hormones and immunity to protozoan parasites. Clin. Microbiol. Rev. 2001, 14, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, I.; Helm, C.S.; Von Samson-Himmelstjerna, G.; Krücken, J.; Kottmann, T.; Holtdirk, A.; Kohn, B.; Hendrickx, G.; Marsboom, C.; Müller, E. Molecular detection of Babesia spp. in dogs in Germany (2007-2020) and identification of potential risk factors for infection. Parasit. Vectors 2023, 16, 396. [Google Scholar] [CrossRef]
- Zuk, M.; McKean, K.A. Sex differences in parasite infections: Patterns and processes. Int. J. Parasitol. 1996, 26, 1009–1023. [Google Scholar] [CrossRef]
- Rubel, F.; Brugger, K.; Chitimia-Dobler, L.; Dautel, H.; Meyer-Kayser, E.; Kahl, O. Atlas of ticks (Acari: Argasidae, Ixodidae) in Germany. Exp. Appl. Acarol. 2021, 84, 183–214. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewska, E.J.; Estrada-Pena, A.; Alsarraf, M.; Kowalec, M.; Bajer, A. Mapping of Dermacentor reticulatus expansion in Poland in 2012–2014. Ticks Tick Borne Dis. 2016, 7, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Ringenier, M.; Putting, M.; Berger, L.; Burgers, S.; Kortekaas, R.; Lenssen, J.; van Roessel, M.; Wijnveld, M.; Madder, M. Novel foci of Dermacentor reticulatus ticks infected with Babesia canis and Babesia caballi in the Netherlands and in Belgium. Parasit. Vectors 2015, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Baneth, G. Babesiosis in dogs and cats—Expanding parasitological and clinical spectra. Vet. Parasitol. 2011, 181, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Davitkov, D.; Vucicevic, M.; Stevanovic, J.; Krstic, V.; Tomanovic, S.; Glavinic, U.; Stanimirovic, Z. Clinical babesiosis and molecular identification of Babesia canis and Babesia gibsoni infections in dogs from Serbia. Acta Vet. Hung. 2015, 63, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Ybanez, A.P.; Ybanez, R.H.D.; Talle, M.G.; Arreglo, R.M.T.; Geens, M.J.C.; Villas, J.G.I., 3rd; Villar, S.R.; Laruga, C.L.; Cao, S.; Moumouni, F.P.A.; et al. Serological and molecular detection of Theileria equi and Babesia caballi in Philippine horses. Ticks Tick Borne Dis. 2018, 9, 1125–1128. [Google Scholar] [CrossRef]
- Banerjee, D.P.; Singh, B.; Gautam, O.P.; Sarup, S. Cell-mediated immune response in equine babesiosis. Trop. Anim. Health Prod. 1977, 9, 153–158. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Categories | n/N Total (%) | Piroplasm Specific PCR | B. caballi cELISA | T. equi cELISA |
|---|---|---|---|---|
| Warmblood horses | 1299/1806 (71.9) | 43/477 (9.0 [6.8; 11.9]) | 49/872 (5.6 [4.3; 7.4]) | 146/878 (16.6 [14.3; 19.2]) |
| Heavy horses | 185/1806 (10.2) | 6/93 (6.5 [2.9; 13.4]) | 12/99 (12.1 [7.1; 20.0]) | 12/100 (12.0 [7.0; 19.8]) |
| Ponies | 185/1806 (10.2) | 5/75 (6.7 [2.9; 14.7]) | 9/112 (8.0 [4.3; 14.6]) | 7/113 (6.2 [3.0; 12.2]) |
| Thoroughbreds | 137/1806 (7.6) | 11/73 (15.1 [8.6; 25.0]) | 3/67 (4.5 [1.5; 12.4]) | 12/67 (17.9 [10.6; 28.7]) |
| Total | 1806/1806 (100) | 65/718 (9.1 [7.2; 11.4]) | 73/1150 (6.3 [5.1; 7.9]) | 177/1158 (15.3 [13.3; 17.5]) |
| Fisher’s exact test 1 | - | p = 0.375 | p = 0.079 | p = 0.013 * |
| Age (N = 1856) | p 1 | ||
|---|---|---|---|
| <9 Years | ≥9 Years | ||
| Piroplasm-specific PCR | 42/348 (12.1 [9.1; 15.9]) | 24/385 (6.2 [4.2; 9.1]) | 0.007 * |
| Theileria equi cELISA | 107/716 (14.9 [12.5; 17.7) | 56/481 (11.6 [9.1; 14.8]) | 0.122 |
| Babesia caballi cELISA | 44/707 (6.2 [4.7; 8.3]) | 26/479 (5.4 [3.7; 7.8]) | 0.617 |
| Sex (N = 2573) | p 1 | ||
| Female | Male | ||
| Piroplasm-specific PCR | 38/413 (9.2 [6.8; 12.4]) | 58/649 (8.9 [7.0; 11.4]) | 0.913 |
| Theileria equi cELISA | 79/577 (13.7 [11.1; 16.7]) | 155/1029 (15.1 [13.0; 17.4]) | 0.507 |
| Babesia caballi cELISA | 35/573 (6.1 [4.4; 8.4]) | 70/1023 (6.8 [5.5; 8.6]) | 0.600 |
| N | B | SE | Wald | p | Odds Ratio | 95%-CI for Odds Ratio | ||
|---|---|---|---|---|---|---|---|---|
| Lower Limit | Upper Limit | |||||||
| Piroplasm-specific PCR | ||||||||
| Age (<9 years) * | 733 | 0.725 | 0.267 | 7.348 | 0.007 * | 2.065 | 1.222 | 3.487 |
| Sex (male) | 1062 | −0.032 | 0.219 | 0.021 | 0.884 | 0.968 | 0.631 | 1.487 |
| Year (year) | 1589 | 0.053 | 0.028 | 3.650 | 0.056 | 1.055 | 0.999 | 1.114 |
| Breed (Warmblood) | 718 | −0.014 | 0.275 | 0.003 | 0.960 | 0.986 | 0.575 | 1.690 |
| Breed (Heavy horses) | 718 | −0.413 | 0.444 | 0.867 | 0.352 | 0.662 | 0.277 | 1.579 |
| Breed (Ponies) | 718 | −0.365 | 0.482 | 0.573 | 0.449 | 0.694 | 0.270 | 1.786 |
| Breed (Thoroughbreds) | 718 | 0.664 | 0.357 | 3.461 | 0.063 | 1.942 | 0.965 | 3.907 |
| Season (Spring) | 1589 | 0.029 | 0.201 | 0.021 | 0.884 | 1.030 | 0.695 | 1.525 |
| Season (Summer) | 1589 | 0.225 | 0.176 | 1.642 | 0.200 | 1.252 | 0.888 | 1.767 |
| Season (Autumn) | 1589 | −0.079 | 0.198 | 0.161 | 0.689 | 0.924 | 0.626 | 1.362 |
| Season (Winter) | 1589 | −0.296 | 0.236 | 1.573 | 0.210 | 0.744 | 0.469 | 1.181 |
| Region (North) | 1589 | −0.280 | 0.473 | 0.350 | 0.554 | 0.756 | 0.299 | 1.911 |
| Region (South) * | 1589 | 0.372 | 0.170 | 4.769 | 0.029 * | 1.451 | 1.039 | 2.026 |
| Region (Central) * | 1589 | −0.483 | 0.179 | 7.265 | 0.007 * | 0.617 | 0.434 | 0.876 |
| Region (West) | 1589 | −0.923 | 0.727 | 1.610 | 0.204 | 0.397 | 0.096 | 1.653 |
| Region (East) * | 1589 | 0.473 | 0.239 | 3.939 | 0.047 * | 1.605 | 1.006 | 2.562 |
| Babesia caballi cELISA | ||||||||
| Age (<9 years) | 1186 | 0.145 | 0.255 | 0.325 | 0.569 | 1.156 | 0.702 | 1.905 |
| Sex (male) | 1596 | 0.121 | 0.241 | 0.322 | 0.570 | 1.129 | 0.742 | 1.717 |
| Year (years) * | 2578 | 0.066 | 0.025 | 7.071 | 0.008 * | 1.068 | 1.017 | 1.121 |
| Breed (Warmblood) | 1150 | −0.462 | 0.259 | 3.173 | 0.075 | 0.630 | 0.379 | 1.047 |
| Breed (Heavy horses) * | 1150 | 0.806 | 0.335 | 5.786 | 0.016 * | 2.239 | 1.161 | 4.316 |
| Breed (Ponies) | 1150 | 0.285 | 0.371 | 0.591 | 0.442 | 1.330 | 0.643 | 2.750 |
| Breed (Thoroughbreds) | 1150 | −0.388 | 0.604 | 0.414 | 0.520 | 0.678 | 0.208 | 2.214 |
| Season (Spring) | 2578 | 0.251 | 0.187 | 1.800 | 0.180 | 1.286 | 0.891 | 1.857 |
| Season (Summer) | 2578 | −0.350 | 0.187 | 3.497 | 0.061 | 0.705 | 0.488 | 1.017 |
| Season (Autumn) | 2578 | 0.278 | 0.161 | 2.968 | 0.085 | 1.320 | 0.963 | 1.810 |
| Season (Winter) | 2578 | −0.263 | 0.212 | 1.527 | 0.217 | 0.769 | 0.507 | 1.166 |
| Region (North) | 2578 | −0.537 | 0.350 | 2.355 | 0.125 | 0.584 | 0.294 | 1.160 |
| Region (South) | 2578 | 0.313 | 0.160 | 3.841 | 0.050 | 1.367 | 1.000 | 1.869 |
| Region (Central) | 2578 | −0.222 | 0.162 | 1.876 | 0.171 | 0.801 | 0.583 | 1.101 |
| Region (West) | 2578 | 0.109 | 0.259 | 0.178 | 0.673 | 1.115 | 0.672 | 1.851 |
| Region (East) | 2578 | 0.059 | 0.310 | 0.036 | 0.850 | 1.060 | 0.577 | 1.948 |
| Theileria equi cELISA | ||||||||
| Age (<9 years) | 1197 | 0.288 | 0.177 | 2.654 | 0.103 | 1.333 | 0.943 | 1.885 |
| Sex (male) | 1606 | 0.111 | 0.149 | 0.558 | 0.455 | 1.118 | 0.834 | 1.498 |
| Year (years) * | 2591 | 0.091 | 0.018 | 26.684 | <0.001 * | 1.095 | 1.058 | 1.134 |
| Breed (Warmblood) * | 1158 | 0.471 | 0.211 | 4.993 | 0.025 * | 1.602 | 1.060 | 2.422 |
| Breed (Heavy horses) | 1158 | −0.304 | 0.319 | 0.906 | 0.341 | 0.738 | 0.395 | 1.380 |
| Breed (Ponies) * | 1158 | −1.079 | 0.399 | 7.309 | 0.007 * | 0.340 | 0.155 | 0.743 |
| Breed (Thoroughbreds) | 1158 | 0.203 | 0.330 | 0.377 | 0.539 | 1.224 | 0.642 | 2.336 |
| Season (Spring) | 2591 | 0.015 | 0.139 | 0.011 | 0.915 | 1.015 | 0.773 | 1.332 |
| Season (Summer) | 2591 | −0.139 | 0.124 | 1.254 | 0.263 | 0.870 | 0.683 | 1.110 |
| Season (Autumn) | 2591 | −0.024 | 0.117 | 0.043 | 0.836 | 0.976 | 0.776 | 1.228 |
| Season (Winter) | 2591 | 0.188 | 0.133 | 2.001 | 0.157 | 1.207 | 0.930 | 1.567 |
| Region (North) * | 2591 | −0.773 | 0.259 | 8.898 | 0.003 * | 0.462 | 0.278 | 0.767 |
| Region (South) * | 2591 | 0.925 | 0.111 | 69.229 | <0.001 * | 2.521 | 2.027 | 3.134 |
| Region (Central) | 2591 | −0.622 | 0.119 | 27.449 | <0.001 * | 0.537 | 0.426 | 0.678 |
| Region (West) | 2591 | −0.351 | 0.209 | 2.828 | 0.093 | 0.704 | 0.468 | 1.060 |
| Region (East) | 2591 | −0.082 | 0.228 | 0.131 | 0.717 | 0.921 | 0.590 | 1.438 |
| Region | n/N Total (%) | PCR | B. caballi cELISA | T. equi cELISA |
|---|---|---|---|---|
| Central Europe | 1695/4060 (41.7) | 51/690 (7.4 [5.7; 9.6]) | 64/1070 (6.0 [4.7; 7.6]) | 116/1079 (10.8 [9.0;12.7]) |
| Eastern Europe | 336/4060 (8.3) | 24/172 (14.0 [9.6; 19.9]) | 12/168 (7.1 [4.1; 12.1]) | 24/169 (14.2 [9.7; 20.3]) |
| Western Europe | 289/4060 (7.1) | 2/48 (4.2 [1.2; 14.0]) | 18/242 (7.4 [4.8; 11.4]) | 28/244 (11.5 [8.1; 16.1]) |
| Northern Europe | 267/4060 (6.6) | 5/66 (7.6 [3.3; 16.5]) | 9/213 (4.2 [2.0; 7.9]) | 17/213 (8.0 [5.0; 12.4]) |
| Southern Europe | 1473/4060 (36.3) | 72/613 (11.7 [9.4; 14.5]) | 72/885 (8.1 [6.5; 10.1]) | 208/886 (23.5 [20.8; 26.4]) |
| Total | 4060/4060 (100) | 154/1589 (9.7 [8.3; 11.2]) | 175/2578 (6.8 [5.9; 7.8]) | 393/2591 (15.2 [13.8; 16.6]) |
| Fisher’s exact test 1 | - | p = 0.014 * | p = 0.193 | p < 0.001 * |
| Timeframes | PCR | B. caballi cELISA | T. equi cELISA |
|---|---|---|---|
| 2008–2012 | 4/141 (2.8 [1.1; 7.1]) | 17/302 (5.6 [3.5; 8.8]) | 28/308 (9.1 [6.4; 12.8]) |
| 2013–2017 | 56/533 (10.5 [8.2; 13.4]) | 36/818 (4.4 [3.2; 6.0]) | 85/825 (10.3 [8.4; 12.6]) |
| 2018–2021 | 94/915 (10.3 [8.5; 12.4]) | 122/1458 (8.4 [7.1; 9,9]) | 280/1458 (19.2 [17.3; 21.3]) |
| Total | 154/1589 (9.7 [8.3; 11.2]) | 175/2578 (6.8 [5.9; 7.8]) | 393/2591 (15.2 [13.8; 16.6]) |
| Fisher’s exact test 1 | p = 0.007 * | p < 0.001 * | p < 0.001 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Axt, C.W.; Springer, A.; Strube, C.; Jung, C.; Naucke, T.J.; Müller, E.; Schäfer, I. Molecular and Serological Detection of Vector-Borne Pathogens Responsible for Equine Piroplasmosis in Europe between 2008 and 2021. Microorganisms 2024, 12, 816. https://doi.org/10.3390/microorganisms12040816
Axt CW, Springer A, Strube C, Jung C, Naucke TJ, Müller E, Schäfer I. Molecular and Serological Detection of Vector-Borne Pathogens Responsible for Equine Piroplasmosis in Europe between 2008 and 2021. Microorganisms. 2024; 12(4):816. https://doi.org/10.3390/microorganisms12040816
Chicago/Turabian StyleAxt, Carla Wiebke, Andrea Springer, Christina Strube, Clarissa Jung, Torsten J. Naucke, Elisabeth Müller, and Ingo Schäfer. 2024. "Molecular and Serological Detection of Vector-Borne Pathogens Responsible for Equine Piroplasmosis in Europe between 2008 and 2021" Microorganisms 12, no. 4: 816. https://doi.org/10.3390/microorganisms12040816
APA StyleAxt, C. W., Springer, A., Strube, C., Jung, C., Naucke, T. J., Müller, E., & Schäfer, I. (2024). Molecular and Serological Detection of Vector-Borne Pathogens Responsible for Equine Piroplasmosis in Europe between 2008 and 2021. Microorganisms, 12(4), 816. https://doi.org/10.3390/microorganisms12040816