
Combined Effects of Lactic Acid Bacteria and Protease on the Fermentation Quality and Microbial Community during 50 Kg Soybean Meal Fermentation Simulating Actual Production Scale

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Microorganisms
2.2. Soybean Meal Fermentation Experiment
2.3. Fermentation Quality, Chemical Composition, and Microbiology Population Analysis
2.3.1. Evaluation of Sensory
2.3.2. Fermentation Quality
2.3.3. Chemical Composition
2.3.4. Microbiology Population
2.4. SDS-PAGE Profile Determination
2.5. Microbial Community Measurement
2.5.1. DNA Extraction
2.5.2. PCR Amplification
2.6. Statistical Analysis
3. Results
3.1. Sensory Evaluation
3.2. The Fermentation Quality
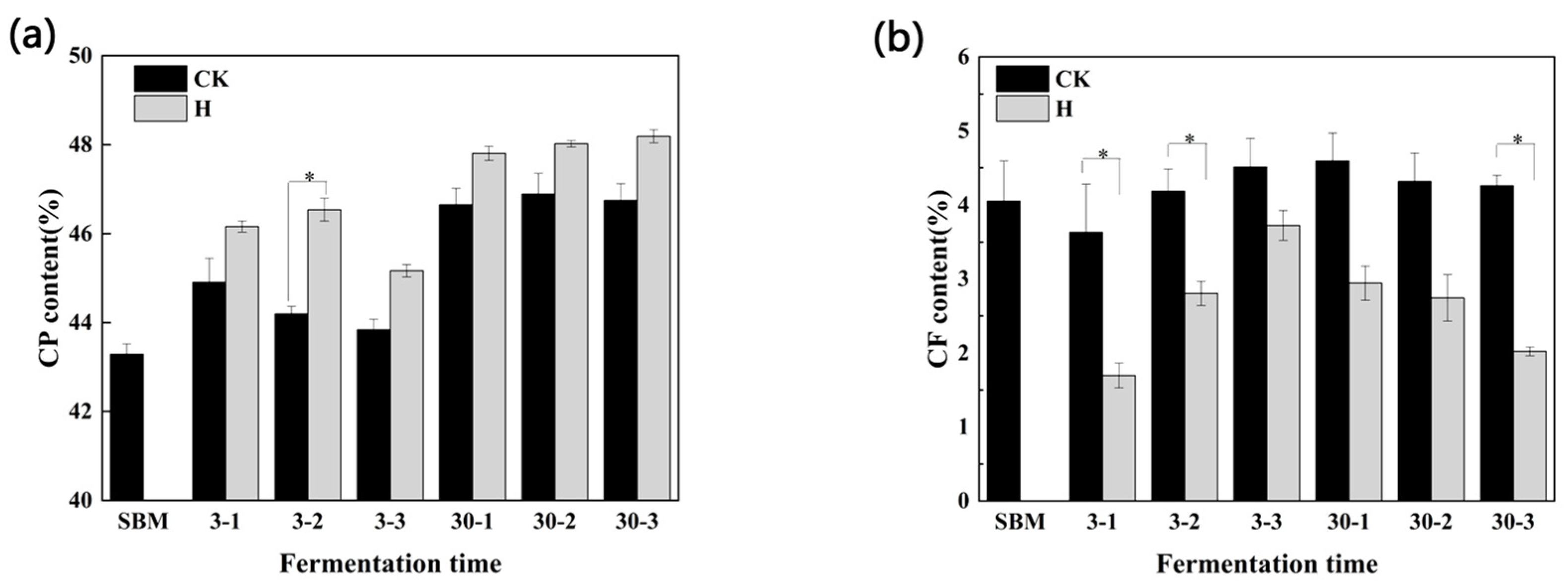
3.3. Chemical Composition Analysis
3.4. Microbial Population of FSBM
3.5. SDS-PAGE Profile
3.6. Microbial Community Dynamics
3.6.1. Alpha Index
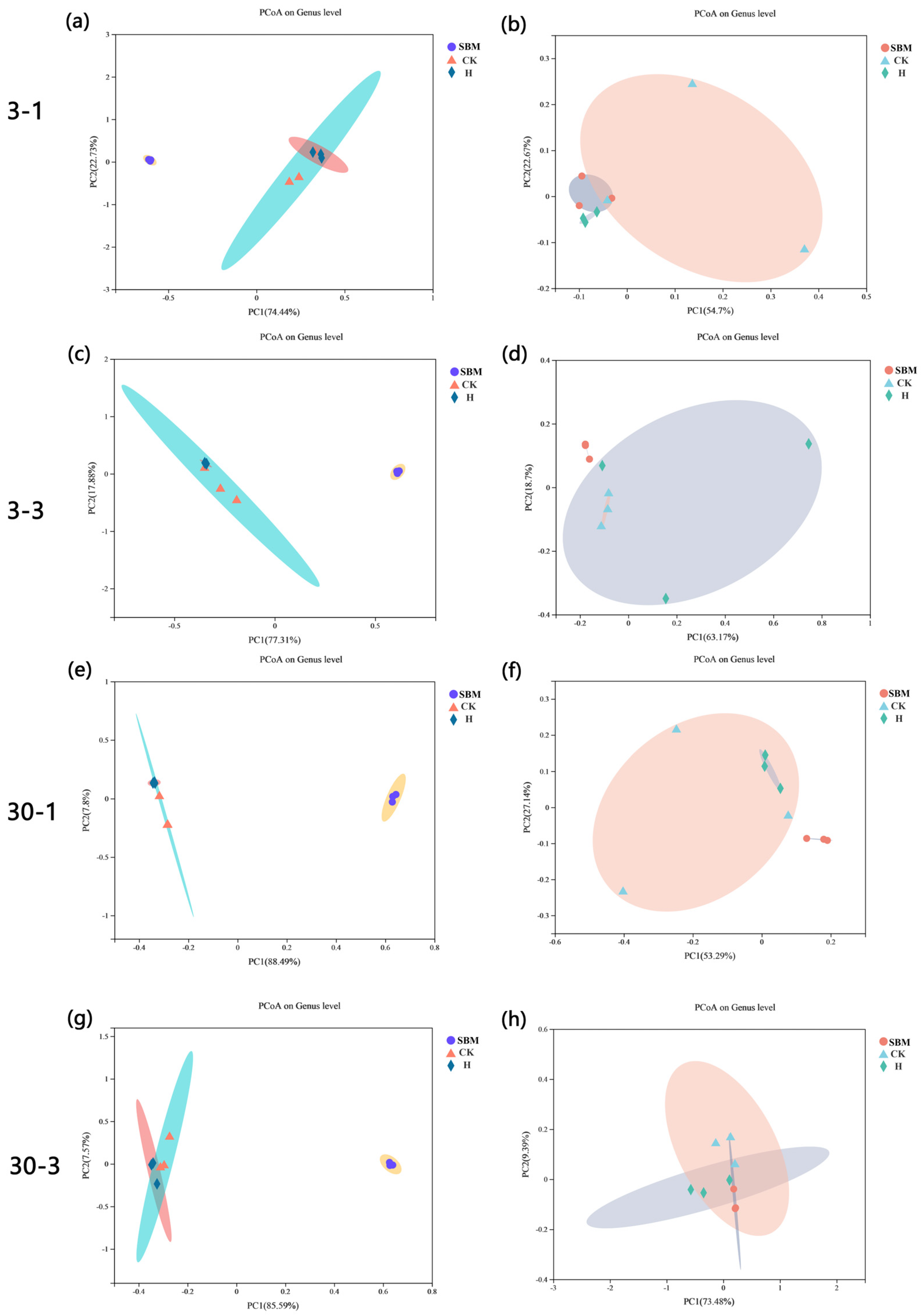
3.6.2. Principal Coordinate Analysis
3.6.3. Abundance of Microbial Community
3.7. Correlation Analysis between Fermentation Quality and Microbial Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zheng, L.; Li, D.; Li, Z.; Kang, L.; Jiang, Y.; Liu, X.; Chi, Y.; Li, Y.; Wang, J. Effects of Bacillus fermentation on the protein microstructure and anti-nutritional factors of soybean meal. Lett. Appl. Microbiol. 2017, 65, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Banaszkiewicz, T. Nutritional Value of Soybean Meal. In Soybean and Nutrition; El-Shemy, H., Ed.; IntechOpen: London, UK, 2011. [Google Scholar]
- Chatterjee, C.; Gleddie, S.; Xiao, C. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Yasar, S.; Tosun, R.; Sonmez, Z. Fungi fermentation inducing improved nutritional qualities associated with the altered secondary protein structure of soybean meal determined by FTIR spectroscopy. Measurement 2020, 161, 107895. [Google Scholar] [CrossRef]
- Kim, S.W.; Less, J.F.; Wang, L.; Yan, T.; Kiron, V.; Kaushik, S.J.; Lei, X. Meeting global feed protein demand: Challenge, opportunity, and strategy. Annu. Rev. Anim. Biosci. 2019, 7, 221–243. [Google Scholar] [CrossRef]
- Emerstorfer, F.; Hein, W.; Resch, R.; Poetsch, E.M.; Zitz, U.; Kneifel, W. Application of plant-based antimicrobials for the growth inhibition of clostridia in pressed beet pulp silage. J. Sci. Food Agric. 2011, 91, 2038–2044. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, C.; Zhu, Q.; Qian, G. Optimization of solid-state fermentation with Lactobacillus brevis and Aspergillus oryzae for trypsin inhibitor degradation in soybean meal. J. Integr. Agric. 2013, 12, 869–876. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, G.Z.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, K.; Maharachchikumbura, N.S.S.; Ekanayaka, H.A.; Qing, T.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Carbonetto, B.; Nidelet, T.; Guezenec, S.; Perez, M.; Segond, D.; Sicard, D. Interactions between Kazachstania humilis yeast species and lactic acid bacteria in sourdough. Microorganisms 2020, 8, 240. [Google Scholar] [CrossRef]
- Hong, K.J.; Lee, C.H.; Kim, S.W. Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. J. Med. Food 2004, 7, 430. [Google Scholar] [CrossRef]
- Feng, J.; Liu, X.; Xu, Z.; Liu, Y.; Lu, Y. Effects of Aspergillus oryzae 3.042 fermented soybean meal on growth performance and plasma biochemical parameters in broilers. Anim. Feed Sci. Technol. 2007, 134, 235–242. [Google Scholar] [CrossRef]
- Rubio, L.A.; Clemente, A. Molecular size distribution affects portal absorption rate of casein amino acids in rats. J. Anim. Physiol. Anim. Nutr. 2010, 94, 145–153. [Google Scholar] [CrossRef]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-origin lactic acid bacteria may exhibit probiotic properties: Review. BioMed Res. Int. 2018, 506, 3185. [Google Scholar] [CrossRef] [PubMed]
- Teneva-Angelova, T.; Hristova, I.; Pavlov, A.; Beshkova, D. Lactic Acid Bacteria—From Nature Through Food to Health. In Advances in Biotechnology for Food Industry; Holban, M.H., Grumezescu, M.A., Eds.; Academic Press: Amsterdam, The Netherlands, 2018; pp. 91–133. [Google Scholar]
- Padhi, S.; Sharma, S.; Sahoo, D.; Montet, D.; Rai, A.K. Potential of lactic acid bacteria as starter cultures for food fermentation and as producers of biochemicals for value addition. In Lactic Acid Bacteria in Food Biotechnology; Ramesh, C.R., Azevedo, C.A.V., Montet, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 281–304. [Google Scholar]
- Wilson, S.; Blaschek, K.; De Mejia, E.G. Allergenic proteins in soybean: Processing and reduction of P34 allergenicity. Nutr. Res. Rev. 2005, 63, 47–58. [Google Scholar] [CrossRef]
- Yang, A.; Zuo, L.; Cheng, Y.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Degradation of major allergens and allergenicity reduction of soybean meal through solid-state fermentation with microorganisms. Food Funct. 2018, 9, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Abduru, S.A. Nutritional Characteristics of Soybean Meal Fermented by Lactobacillus acidophilus and Its Feeding Effect on Yellow-Feathered Broilers. Master’s Thesis, Shihezi University, Xinjiang, China, 2014. (In Chinese). [Google Scholar]
- Morales-Mena, A.; Martínez-González, S.; Teague, K.D.; Graham, L.E.; Señas-Cuesta, R.; Vuong, C.N.; Lester, H.; Hernandez-Patlan, D.; Solis-Cruz, B.; Fuente-Martinez, B.; et al. Assessment of fermented soybean meal on Salmonella typhimurium infection in neonatal turkey poults. Animals 2020, 10, 1849. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, B.; Liu, C.; Zhou, H.; Wang, X.; Mai, K.; He, G. Effects of dietary raw or Enterococcus faecium fermented soybean meal on growth, antioxidant status, intestinal microbiota, morphology, and inflammatory responses in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2020, 100, 261–271. [Google Scholar] [CrossRef]
- Wang, H. Study on In Vitro Probiotics of Pediococcus pentosaccharides NCu301 and Its Application in Soybean Meal Fermentation. Master’s Thesis, Nanchang University, Jiangxi, China, 2018. (In Chinese). [Google Scholar]
- Cao, K. Screening and Identification of Lactic Acid Bacteria with High Bile Salt Tolerance and Its Application in Soybean Meal Fermentation. Master’s Thesis, Jiangxi Agricultural University, Jiangxi, China, 2015. (In Chinese). [Google Scholar]
- Zhao, Y.; Sun-Waterhouse, D.; Zhao, M.; Zhao, Q.; Qiu, C.; Su, G. Effects of solid-state fermentation and proteolytic hydrolysis on defatted soybean meal. LWT Food Sci. Technol. 2018, 97, 496–502. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Zhang, L. Optimization of lactic acid fermentation conditions for fermented tofu whey beverage with high-isoflavone aglycones. LWT Food Sci. Technol. 2019, 111, 211–217. [Google Scholar] [CrossRef]
- Singracha, P.; Niamsiri, N.; Visessanguan, W.; Lertsiri, S.; Assavanig, A. Application of lactic acid bacteria and yeasts as starter cultures for reduced salt soy sauce (moromi) fermentation. LWT Food Sci. Technol. 2017, 78, 181–188. [Google Scholar] [CrossRef]
- Fernandez, M.; Hudson, J.; Korpela, R.; de los Reyes-Gavilán, C.G. Impact on human health of microorganisms present in fermented dairy products: An overview. BioMed Res. Int. 2015, 2015, 412714. [Google Scholar] [CrossRef]
- Ma, H.; Wang, W.; Wang, Z.; Tan, Z.; Qin, G.; Wang, Y.; Pang, H. Microbial population succession and community diversity and its correlation with fermentation quality in soybean meal treated with Enterococcus faecalis during fermentation and aerobic exposure. Microorganisms 2022, 10, 530. [Google Scholar] [CrossRef]
- Menlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physico- chemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef]
- Miao, X.; Niu, H.; Sun, M.; Li, D.; Hua, M.; Wang, J.; Su, Y. Structural characterization and properties of modified soybean meal protein via solid-state fermentation by Bacillus subtilis. Molecules 2023, 28, 8015. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Pan, J.; Miao, S.; Qin, L. Microbial community in Chinese traditional fermented acid rice soup (rice-acid) and its correlations with key organic acids and volatile compounds. Food Res. Int. 2020, 137, 109672. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.R.; Moss, M.O. Food Microbiology, 3rd ed.; The Royal Society of Chemistry: Cambridge, UK, 2008; p. 463. [Google Scholar]
- Kim, J.Y.; Yeo, S.H.; Baek, S.Y.; Choi, H.S. Molecular and morphological identification of fungal species isolated from bealmijang meju. J. Microbiol. Biotechn. 2011, 21, 1270–1279. [Google Scholar] [CrossRef]
- Dunn, M.J.; Bradd, S.J. Separation and Analysis of Membrane Proteins by SDS-Polyacrylamide Gel Electrophoresis. In Methods in Molecular Biology; Graham, J.M., Higgins, J.A., Eds.; Humana Press: Totowa, NJ, USA, 1998. [Google Scholar]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.D. Saccharomyces and non-Saccharomyces competition during microvinification under different sugar and nitrogen conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef]
- Palacios, M.F.; Easter, R.A.; Soltwedel, K.T.; Parsons, C.M.; Douglas, M.W.; Hymowitz, T.; Pettigrew, J.E. Effect of soybean variety and processing on growth performance of young chicks and pigs. J. Anim. Sci. 2004, 82, 1108–1114. [Google Scholar] [CrossRef]
- De Vuyst, L.; Weckx, S. The cocoa bean fermentation process: From ecosystem analysis to starter culture development. J. Appl. Microbiol. 2016, 121, 5–17. [Google Scholar] [CrossRef]
- Dong, Q.; Saneoka, H. Physiological characteristics, phytase activity, and mineral bioavailability of a low-phytate soybean line during germination. Plant Food Hum. Nutr. 2020, 75, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Khan, N.; Wang, Z.; Yu, P. Relationship of feeds protein structural makeup in common Prairie feeds with protein solubility, in situ ruminal degradation, and intestinal digestibility. Anim. Feed Sci. Technol. 2014, 194, 58–70. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of fermentation in improving nutritional quality of soybean meal—A review. Asian-Australas. J. Anim. Sci. 2015, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Yafetto, L.; Odamtten, G.T.; Wiafe-Kwagyan, M. Valorization of agro-industrial wastes into animal feed through microbial fermentation: A review of the global and Ghanaian case. Heliyon 2023, 9, 14814. [Google Scholar] [CrossRef] [PubMed]
- Niba, A.T.; Beal, J.D.; Kudi, A.C.; Brooks, P.H. Potential of bacterial fermentation as a biosafe method of improving feeds for pigs and poultry. Afr. J. Biotechnol. 2009, 8, 1758–1767. [Google Scholar]
- Chen, Q.; Liu, B.; Liu, G.; Shi, H.; Wang, J. Effect of Bacillus subtilis and Lactobacillus plantarum on solid-state fermentation of soybean meal. J. Sci. Food Agric. 2023, 103, 6070–6079. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, K.Z.; Mahran, M.Z.; Ramadan, M.M.; Ghanem, H.Z.; Fadel, M.; Mahmoud, M.H. A comparative study on flavour components and therapeutic properties of unfermented and fermented defatted soybean meal extract. Sci. Rep. 2020, 10, 5998. [Google Scholar] [CrossRef] [PubMed]
- Boukid, F.; Ganeshan, S.; Wang, Y.; Tülbek, M.C.; Nickerson, M.T. Bioengineered enzymes and precision fermentation in the food industry. Int. J. Mol. Sci. 2023, 24, 10156. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Dynamics changes of microorganisms community and fermentation quality in soybean meal prepared with lactic acid bacteria and Artemisia argyi through fermentation and aerobic exposure processes. Foods 2022, 11, 795. [Google Scholar] [CrossRef]
- Ko, Y.D.; Kim, J.H.; Adesogan, A.T.; Ha, H.M.; Kim, S.C. The effect of replacing rice straw with dry wormwood (Artemisia sp.) on intake, digestibility, nitrogen balance and ruminal fermentation characteristics in sheep. Anim. Feed Sci. Technol. 2006, 125, 99–110. [Google Scholar] [CrossRef]
- Huang, X.; Li, H.; Han, T.; Wang, J.; Ma, Z.; Yu, X. Isolation and identification of protease-producing Bacillus amyloliquefaciens LX-6 and its application in the solid fermentation of soybean meal. Front. Bioeng. Biotech. 2023, 11, 1226988. [Google Scholar] [CrossRef]
- Jones, D.J.C. The biochemistry of silage. J. Agric. Sci. 1991, 2, 117. [Google Scholar]
- Shi, C.; Zhang, Y.; Lu, Z. Solid-state fermentation of corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium for degrading antinutritional factors and enhancing nutritional value. J. Anim. Sci. Biotechnol. 2017, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Hutkins, R.W. Microbiology and Technology of Fermented Foods, 2nd ed.; John Wiley Press: London, UK, 2019. [Google Scholar]
- Hansen, E.B. Starter cultures: Uses in the Food Industry. In Encyclopedia of Food Microbiology; Academic Press: London, UK; Amsterdam, The Netherlands, 2014. [Google Scholar]
- McAllister, T.A.; Dunière, L.; Drouin, P.; Xu, S. Silage review: Using molecular approaches to define the microbial ecology of silage. J. Dairy Sci. 2018, 101, 4060–4074. [Google Scholar] [CrossRef] [PubMed]
- Namihira, T.; Shinzato, N.; Akamine, H.; Maekawa, T.M. Influence of nitrogen fertilization on tropical-grass silage assessed by ensiling process monitoring using chemical and microbial community analyses. J. Appl. Microbiol. 2010, 108, 1954–1965. [Google Scholar] [CrossRef] [PubMed]
- Preston, G.M.; Studholme, D.J.; Caldelari, I. Profiling the secretomes of plant pathogenic proteobacteria. FEMS Microbiol. Rev. 2005, 29, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Jiang, Y.; Cervantes, A.A.P.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157: H7 and silage additives. J. Dairy Sci. 2017, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, L.; Yu, H.; Wang, W.; Wu, G.; Qin, G.; Tan, Z.; Wang, Y.; Pang, H. Protease-producing lactic acid bacteria with antibacterial properties and their potential use in soybean meal fermentation. Chem. Biol. Technol. Agric. 2022, 9, 1. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of microbial community and fermentation quality in corn silage treated with lactic acid bacteria and Artemisia argyi during aerobic exposure. Toxins 2022, 14, 349. [Google Scholar] [CrossRef]
- Wang, Z.; Tan, Z.; Wu, G.; Wang, L.; Qin, G.; Wang, Y.; Pang, H. Microbial community and fermentation characteristic of whole-crop wheat silage treated by lactic acid bacteria and Artemisia argyi during ensiling and aerobic exposure. Front. Microbiol. 2022, 13, 1004495. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tan, Z.; Wu, G.; Wang, L.; Qin, G.; Wang, Y.; Pang, H. Investigation on fermentation characteristics and microbial communities of wheat straw silage with different proportion Artemisia argyi. Toxins 2023, 15, 330. [Google Scholar] [CrossRef]
- Wu, Z.; Luo, Y.; Bao, J.; Luo, Y.; Yu, Z. Additives affect the distribution of metabolic profile, microbial communities and antibiotic resistance genes in high-moisture sweet corn kernel silage. Bioresource Technol. 2020, 315, 123821. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Hou, Y.; Xu, H.; Huang, L.; Dabbour, M.; Mintah, B.K.; He, R.; Ma, H. Effect of solid-state fermentation by three different Bacillus species on composition and protein structure of soybean meal. J. Sci. Food Agric. 2022, 102, 557. [Google Scholar] [CrossRef] [PubMed]
- Fabiszewska, A.U.; Zielińska, K.J.; Wróbel, B. Trends in designing microbial silage quality by biotechnological methods using lactic acid bacteria inoculants: A minireview. World J. Microb. Biotechnol. 2019, 35, 76. [Google Scholar] [CrossRef] [PubMed]
- Spohner, S.C.; Müller, H.; Quitmann, H.; Czermak, P. Expression of enzymes for the usage in food and feed industry with Pichia pastoris. J. Biotechnol. 2015, 202, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Overwintering of vineyard yeasts: Survival of interacting yeast communities in grapes mummified on vines. Front. Microbiol. 2016, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Roncero, M.I.G.; Concepción, H.; Manuel, R.R. Fusarium as a model for studying virulence in soilborne plant pathogens. Physiol. Mol. Plant Pathol. 2003, 62, 87–98. [Google Scholar] [CrossRef]
- Suo, B.; Nie, W.; Wang, Y.; Ma, J.; Ai, Z. Microbial diversity of fermented dough and volatile compounds in steamed bread prepared with traditional Chinese starters. LWT-Food Sci. Technol. 2020, 126, 109350. [Google Scholar] [CrossRef]
- Ali, N.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Microbial diversity and fermentation profile of red clover silage inoculated with reconstituted indigenous and exogenous epiphytic microbiota. Bioresour. Technol. 2020, 314, 123606. [Google Scholar] [CrossRef]
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces cerevisiae metabolites produced during fermentation on bread quality parameters: A review. Crit. Rev. Food Sci. Nutr. 2018, 587, 1152–1164. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Wang, S.; Zhou, R.; Hu, X.; Chu, Y.; Wang, T. Characteristics of yeast flora in Chinese strong-flavoured liquor fermentation in the Yibin region of China. J. Inst. Brew. 2016, 122, 517–523. [Google Scholar] [CrossRef]
- Dong, T.; Liang, Y.; Shao, T.; Wang, W.; Ma, P.; Wang, W.; Li, J.; Yuan, X. Detoxifying mycotoxins and antifungal properties of two rumen-derived Enterococcus species in artificially contaminated corn silages. J. Sci. Food Agric. 2023, 103, 5981–5991. [Google Scholar] [CrossRef] [PubMed]
- Adewara, O.A.; Ogunbanwo, S.T. Acid stress responses of Lactobacillus amylovorus and Candida kefyr isolated from fermented sorghum gruel and their application in food fermentation. Can. J. Microbiol. 2022, 68, 269–280. [Google Scholar] [CrossRef]
- Skiada, A.; Pavleas, I.; Drogari-Apiranthitou, M. Epidemiology and diagnosis of Mucormycosis: An update. J. Fungi 2020, 6, 265. [Google Scholar] [CrossRef]
- Divyashree, S.; Shruthi, B.; Vanitha, P.R.; Sreenivasa, M.Y. Probiotics and their postbiotics for the control of opportunistic fungal pathogens: A review. Biotechnol. Rep. 2023, 38, e00800. [Google Scholar] [CrossRef]
- Menden, A.; Hall, D.; Broedlow, C.A.; Darcey, T.; Crawford, F.; Klatt, N.; Crynen, S.; Mullan, M.; Ait-Ghezala, G. Candida rugosa lipase alters the gastrointestinal environment in wild-type mice. Biomed. Pharmacother. 2020, 130, 110579. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment | Days of Ensiling (d) | SEM | p Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| 3-1 | 3-2 | 3-3 | 30-1 | 30-2 | 30-3 | ||||
| LAB | CK | 9.88 Ba | 9.79 Ba | 9.33 Bab | 8.59 Abc | 8.43 Bbc | 7.86 Bc | 0.24 | <0.001 |
| H | 11.31 Aa | 10.92 Aa | 10.80 Aac | 9.41 Abc | 9.72 Ac | 9.21 Abc | |||
| Yeast | CK | 4.06 Aa | 4.97 Aa | 5.51 Aab | ND Aab | ND Bab | ND Bb | 0.90 | <0.001 |
| H | ND Bb | 0.12 Aa | 0.36 Aa | 0.57 Aa | 0.15 Aa | 0.07 Aa | |||
| Bacilli | CK | 0.10 Ac | 0.06 Ac | 0.03 Ac | 0.06 Aa | 0.14 Ab | 0.27 Aa | 0.18 | <0.001 |
| H | 0.06 Ab | 0.14 Ab | 0.13 Ab | 0.46 Aa | 0.07 Aa | 0.20 Ba | |||
| Coliform bacteria | CK | 9.86 Aa | 9.89 Aa | 9.97 Aa | 7.74 Ab | 7.35 Ab | 7.26 Ab | 0.25 | <0.001 |
| H | 7.44 Ba | 5.57 Bc | 7.00 Bab | 5.92 Bbc | 5.46 Bc | 5.34 Bc | |||
| Aerobic bacteria | CK | 9.93 Aa | 9.99 Aa | 9.62 Aa | 9.46 Aa | 9.00 Aa | 5.77 Ab | 0.28 | <0.001 |
| H | 9.72 Aa | 9.84 Aa | 9.44 Aa | 6.73 Bbc | 7.23 Bb | 5.57 Ac | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, H.; Zhang, X.; Chen, C.; Ma, H.; Tan, Z.; Zhang, M.; Duan, Y.; Qin, G.; Wang, Y.; Jiao, Z.; et al. Combined Effects of Lactic Acid Bacteria and Protease on the Fermentation Quality and Microbial Community during 50 Kg Soybean Meal Fermentation Simulating Actual Production Scale. Microorganisms 2024, 12, 1339. https://doi.org/10.3390/microorganisms12071339
Pang H, Zhang X, Chen C, Ma H, Tan Z, Zhang M, Duan Y, Qin G, Wang Y, Jiao Z, et al. Combined Effects of Lactic Acid Bacteria and Protease on the Fermentation Quality and Microbial Community during 50 Kg Soybean Meal Fermentation Simulating Actual Production Scale. Microorganisms. 2024; 12(7):1339. https://doi.org/10.3390/microorganisms12071339
Chicago/Turabian StylePang, Huili, Xinyu Zhang, Chen Chen, Hao Ma, Zhongfang Tan, Miao Zhang, Yaoke Duan, Guangyong Qin, Yanping Wang, Zhen Jiao, and et al. 2024. "Combined Effects of Lactic Acid Bacteria and Protease on the Fermentation Quality and Microbial Community during 50 Kg Soybean Meal Fermentation Simulating Actual Production Scale" Microorganisms 12, no. 7: 1339. https://doi.org/10.3390/microorganisms12071339
APA StylePang, H., Zhang, X., Chen, C., Ma, H., Tan, Z., Zhang, M., Duan, Y., Qin, G., Wang, Y., Jiao, Z., & Cai, Y. (2024). Combined Effects of Lactic Acid Bacteria and Protease on the Fermentation Quality and Microbial Community during 50 Kg Soybean Meal Fermentation Simulating Actual Production Scale. Microorganisms, 12(7), 1339. https://doi.org/10.3390/microorganisms12071339