
Diversity, Methane Oxidation Activity, and Metabolic Potential of Microbial Communities in Terrestrial Mud Volcanos of the Taman Peninsula

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site Description, Sample Collection, and Chemical Analysis
2.2. Methane Content and Rates of Microbial Methane Oxidation
2.3. DNA Extraction and 16S rRNA Gene Amplicon and Metagenome Library Preparation, Sequencing, and Analysis
3. Results and Discussion
3.1. Geochemical Characteristics of the Studied Sites
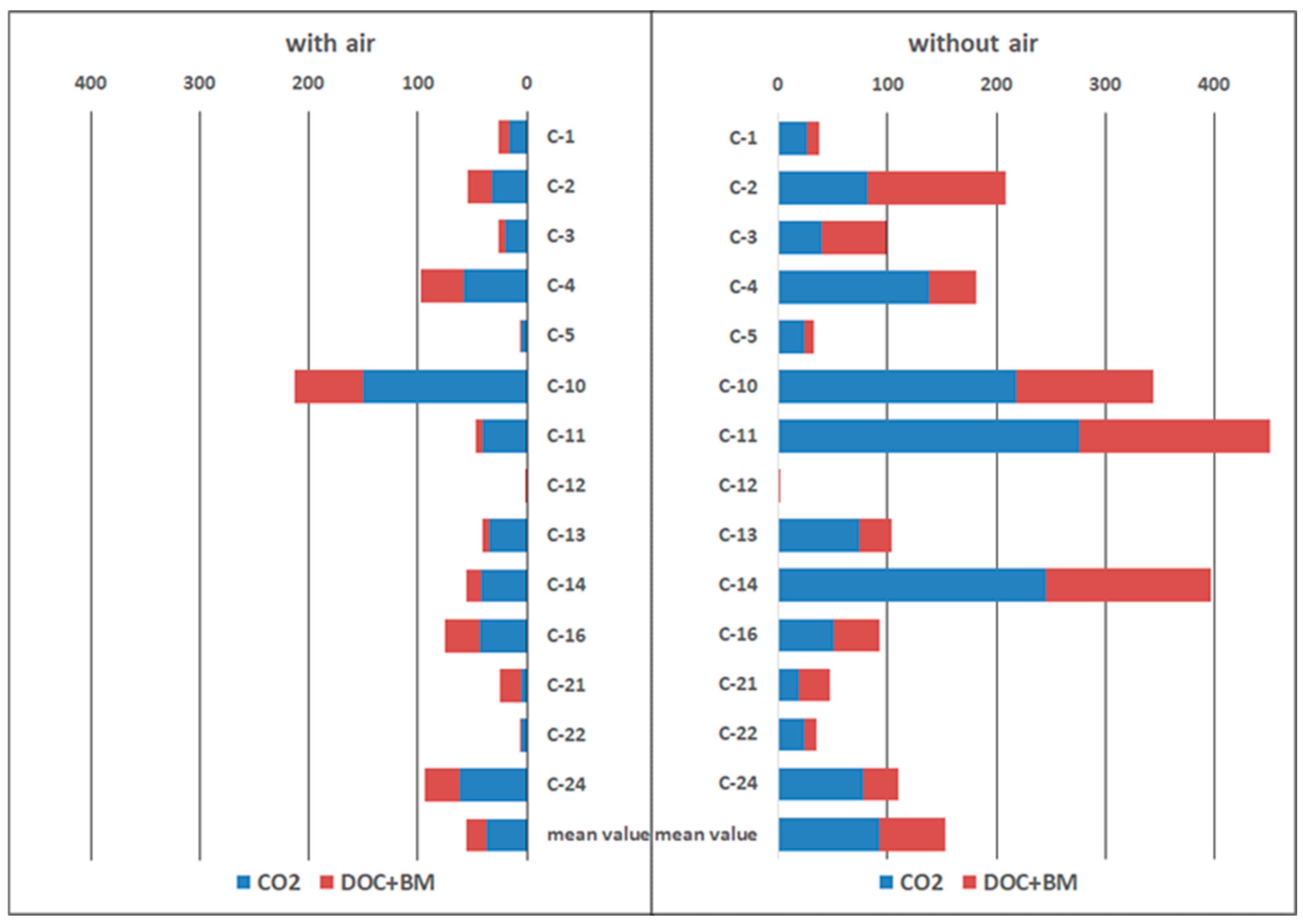
3.2. Microbial Methane Oxidation
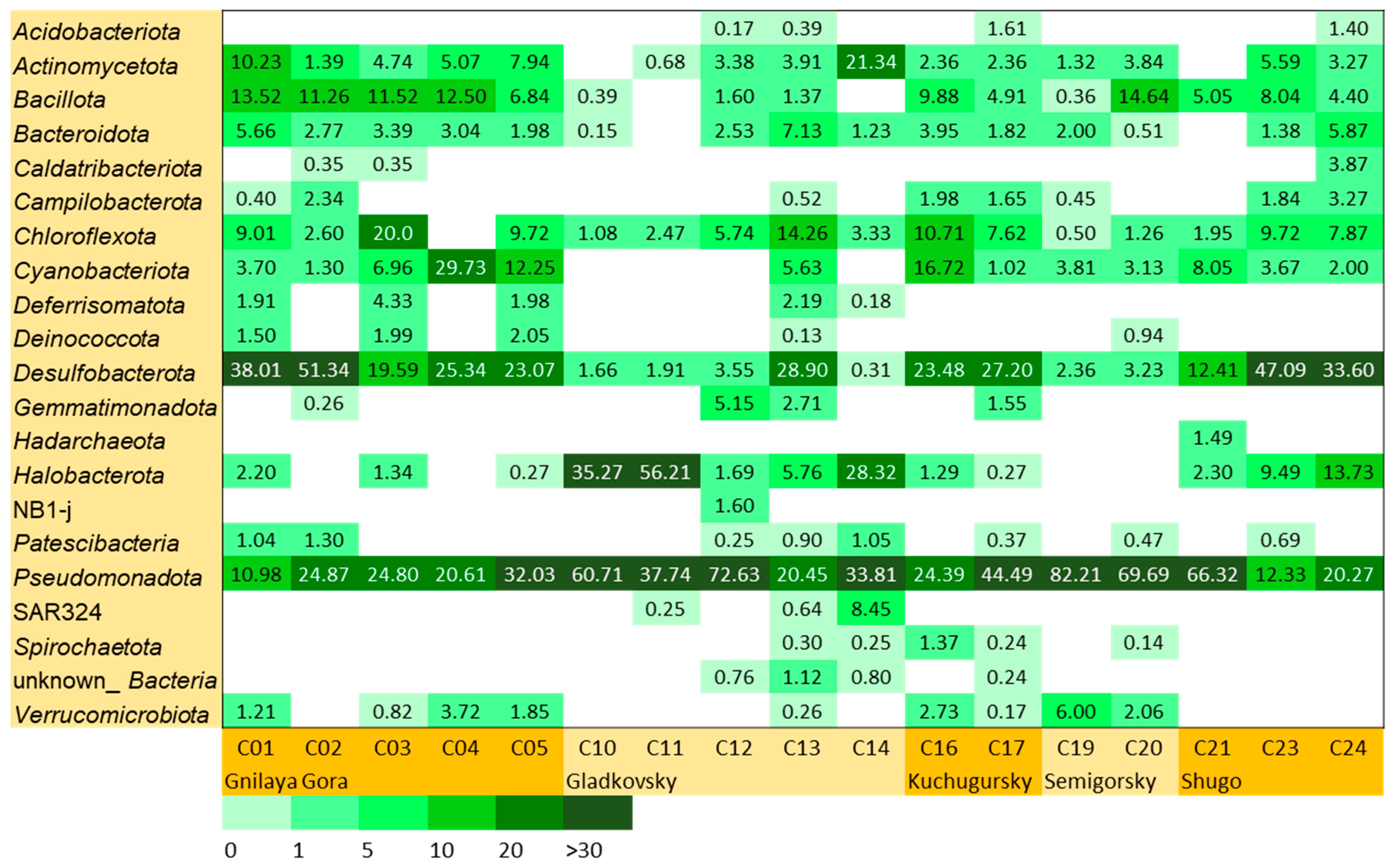
3.3. Microbial Community Abundance and Composition
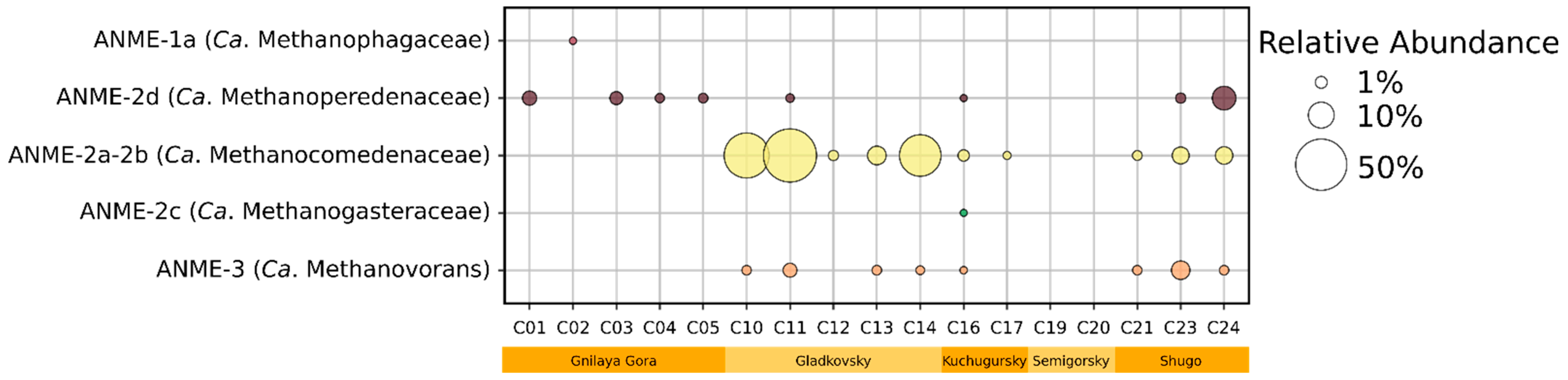
3.4. MAGs’ General Characteristics and Phylogenetic Identification
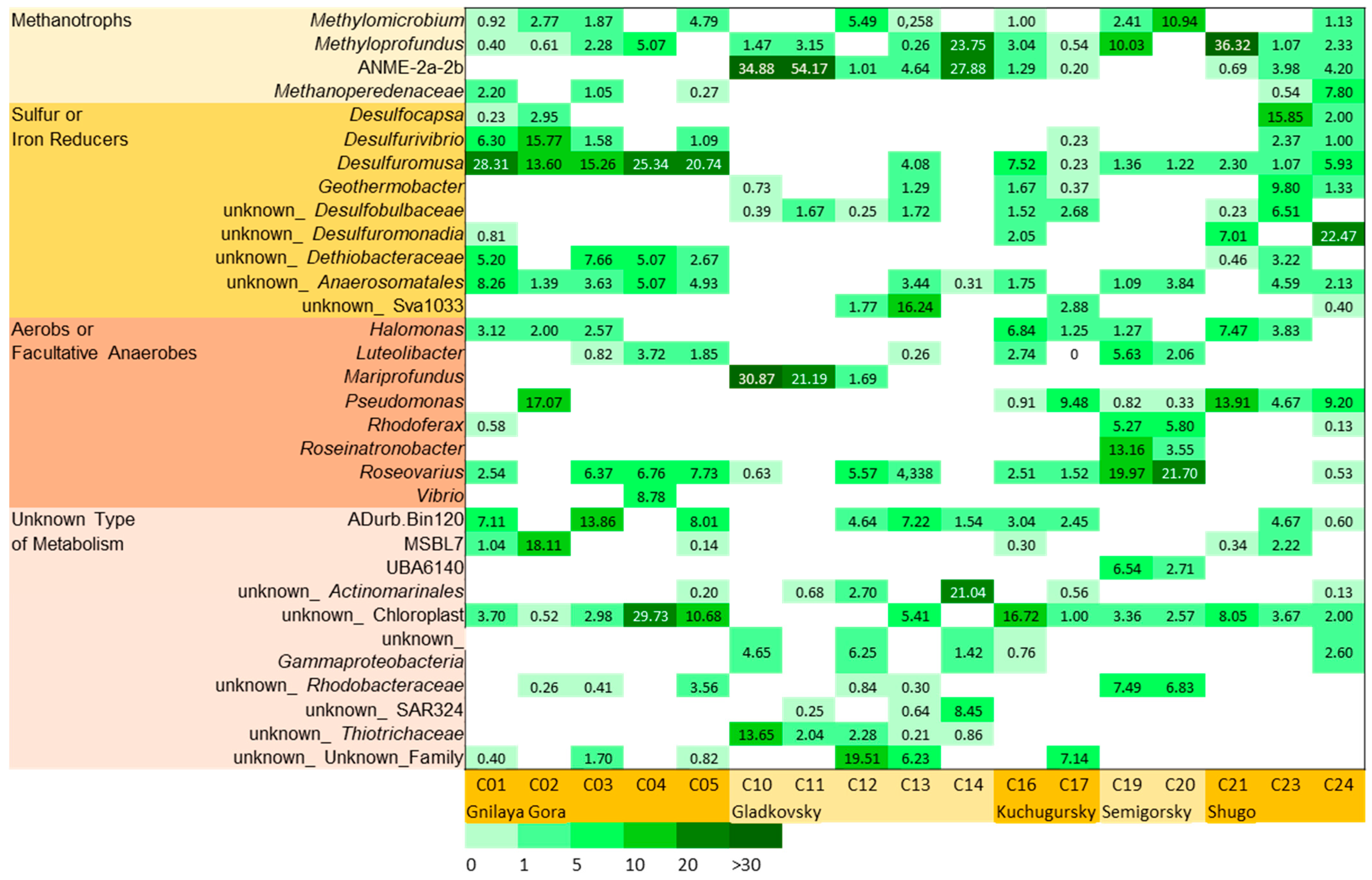
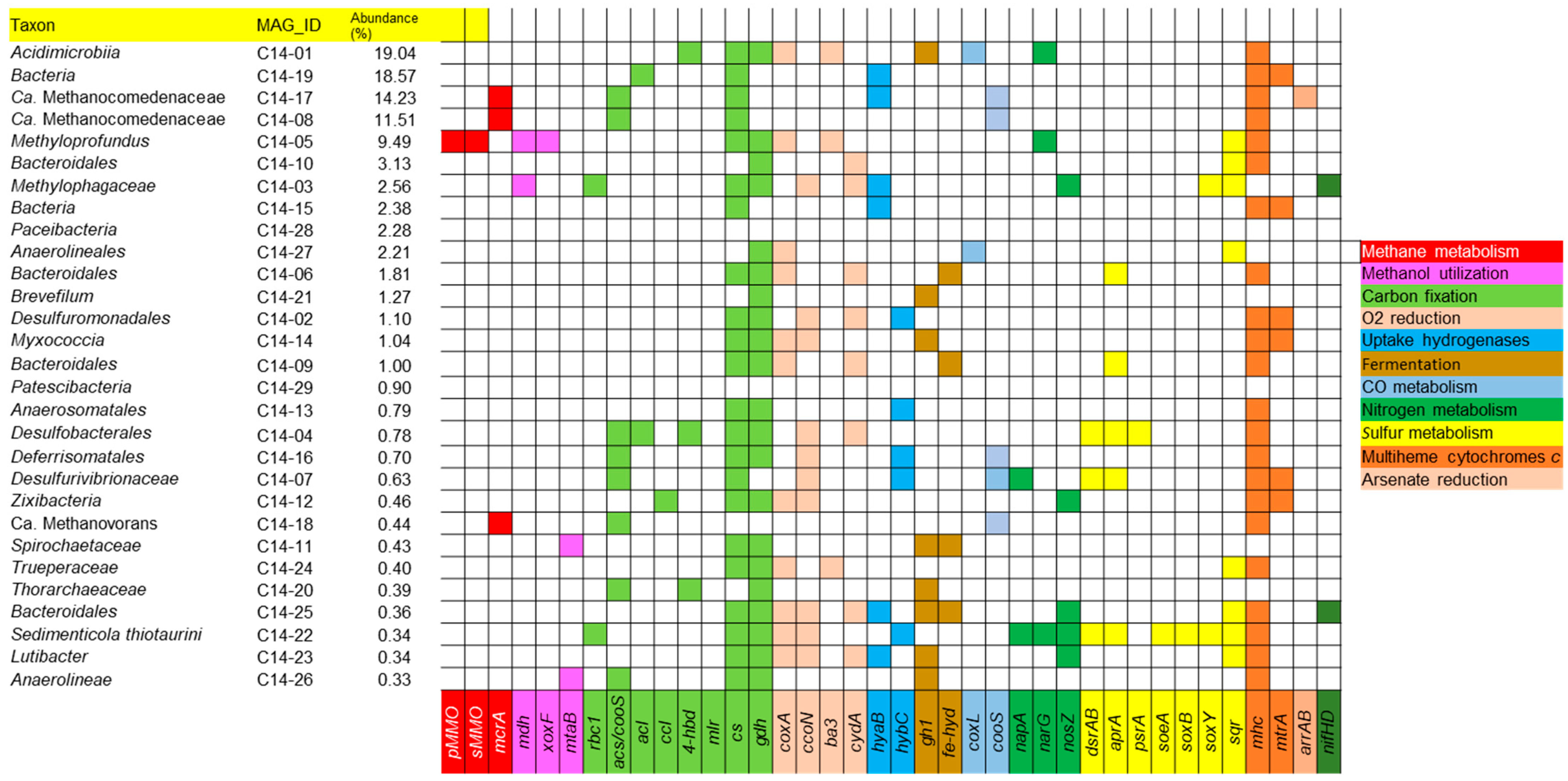
3.5. Metabolic Capabilities in MAGs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mazzini, A.; Etiope, G. Mud volcanism: An updated review. Earth Sci. Rev. 2017, 168, 81–112. [Google Scholar] [CrossRef]
- Kopf, A.J. Significance of mud volcanism. Rev. Geophys. 2002, 40, 2-1–2-52. [Google Scholar] [CrossRef]
- Tu, T.-H.; Wu, L.-W.; Lin, Y.-S.; Imachi, H.; Lin, L.-H.; Wang, P.-L. Microbial community composition and functional capacity in a terrestrial ferruginous, sulfate-depleted mud volcano. Front. Microbiol. 2017, 8, 2137. [Google Scholar] [CrossRef] [PubMed]
- Slobodkin, A.I.; Ratnikova, N.M.; Slobodkina, G.B.; Klyukina, A.A.; Chernyh, N.A.; Merkel, A.Y. Composition and metabolic potential of Fe(III)-reducing enrichment cultures of methanotrophic ANME-2a archaea and associated bacteria. Microorganisms 2023, 11, 555. [Google Scholar] [CrossRef]
- Alain, K.; Holler, T.; Musat, F.; Elvert, M.; Treude, T.; Krüger, M. Microbiological investigation of methane- and hydrocarbon-discharging mud volcanoes in the Carpathian Mountains, Romania. Environ. Microbiol. 2006, 8, 574–590. [Google Scholar] [CrossRef] [PubMed]
- Arun, A.B.; Young, C.C.; Chen, W.M.; Hung, M.H.; Lai, W.A.; Chou, J.H.; Rekha, P.D.; Shen, F.T.; Su, S.P. Belliella pelovolcani sp. nov., isolated from a mud-volcano in Taiwan. Int. J. Syst. Evol. Microbiol. 2009, 59, 2534–2537. [Google Scholar] [PubMed]
- Ghiasian, M.; Akhavan Sepahy, A.; Amoozegar, M.; Saadatmand, S.; Shavandi, M. Bacterial diversity determination using culture-dependent and culture-independent methods. Glob. J. Environ. Sci. Manag. 2017, 3, 153–164. [Google Scholar] [CrossRef]
- Sepanian, E.; Sepahy, A.A.; Hosseini, F. Isolation and characterization of bacterial species from Ain mud volcano, Iran. Microbiology 2018, 87, 282–289. [Google Scholar] [CrossRef]
- Frolova, A.A.; Merkel, A.Y.; Kuchierskaya, A.A.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Pseudodesulfovibrio alkaliphilus, sp. nov., an alkaliphilic sulfate-reducing bacterium isolated from a terrestrial mud volcano. Antonie Leeuwenhoek 2021, 114, 1387–1397. [Google Scholar] [CrossRef]
- Ratnikova, N.M.; Slobodkin, A.I.; Merkel, A.Y.; Kopitsyn, D.S.; Kevbrin, V.V.; Bonch-Osmolovskaya, E.A.; Slobodkina, G.B. Sulfurimonas crateris sp. nov., a facultative anaerobic sulfur-oxidizing chemolithoautotrophic bacterium isolated from a terrestrial mud volcano. Int. J. Syst. Evol. Microbiol. 2020, 70, 487–492. [Google Scholar]
- Manna, S.K.; Das, B.K.; Mohanty, B.P.; Bandopadhyay, C.; Das, N.; Baitha, R.; Das, A.K. Exploration of heterotrophic bacterial diversity in sediments of the mud volcano in the Andaman and Nicobar Islands, India. Environ. Nanotechnol. Monit. Manag. 2021, 16, 100465. [Google Scholar]
- Wrede, C.; Brady, S.; Rockstroh, S.; Dreier, A.; Kokoschka, S.; Heinzelmann, S.M.; Heller, C.; Reitner, J.; Taviani, M.; Daniel, R.; et al. Aerobic and anaerobic methane oxidation in terrestrial mud volcanoes in the Northern Apennines. Sediment. Geol. 2012, 263, 210–219. [Google Scholar] [CrossRef]
- Green-Saxena, A.; Feyzullayev, A.; Hubert, R.J.; Kallmeyer, J.; Krueger, M.; Sauer, P.; Schulz, H.-M.; Orphan, V.J. Active sulfur cycling by diverse mesophilic and thermophilic microorganisms in terrestrial mud volcanoes of Azerbaijan. Environ. Microbiol. 2012, 14, 3271–3286. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.W.; Chang, Y.H.; Tang, S.L.; Tseng, C.H.; Chiang, P.W.; Chang, K.T.; Sun, C.H.; Chen, Y.G.; Kuo, H.C.; Wang, C.H.; et al. Metabolic stratification driven by surface and subsurface interactions in a terrestrial mud volcano. ISME J. 2012, 6, 2280–2290. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-L.; Chiu, Y.-P.; Cheng, T.-W.; Chang, Y.-H.; Tu, W.-X.; Lin, L.-H. Spatial variations of community structures and methane cycling across a transect of Lei-Gong-Hou mud volcanoes in eastern Taiwan. Front. Microbiol. 2014, 5, 121. [Google Scholar] [CrossRef]
- Yang, H.-M.; Lou, K.; Sun, J.; Zhang, T.; Ma, X.-L. Prokaryotic diversity of an active mud volcano in the Usu City of Xinjiang, China. J. Basic Microbiol. 2012, 52, 79–85. [Google Scholar] [CrossRef]
- Lin, Y.T.; Tu, T.H.; Wei, C.L.; Rumble, D.; Lin, L.H.; Wang, P.L. Steep redox gradient and biogeochemical cycling driven by deeply sourced fluids and gases in a terrestrial mud volcano. FEMS Microbiol. Ecol. 2018, 94, fiy171. [Google Scholar] [CrossRef]
- Ren, G.; Ma, A.; Zhang, Y.; Deng, Y.; Zheng, G.; Zhuang, X.; Zhuang, G.; Fortin, D. Electron acceptors for anaerobic oxidation of methane drive microbial community structure and diversity in mud volcanoes. Environ. Microbiol. 2018, 20, 2370–2385. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Haque, S.; Beckles, D.; Schmitt-Kopplin, P.; Harir, M.; Schneider, B.; Stumpp, C.; Wagner, D.A. A chemical and microbial characterization of selected mud volcanoes in Trinidad reveals pathogens introduced by surface water and rain water. Sci. Total Environ. 2020, 707, 136087. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.-H.; Chen, L.-L.; Chiu, Y.-P.; Lin, L.-H.; Wu, L.-W.; Italiano, F.; Shyu, J.B.H.; Raisossadat, S.N.; Wang, P.-L. The biogeographic pattern of microbial communities inhabiting terrestrial mud volcanoes across the Eurasian continent. Biogeosciences 2022, 19, 831–843. [Google Scholar] [CrossRef]
- Megyes, M.; Móga, J.; Strat, D.; Borsodi, A.K. Bacterial and archaeal taxonomic diversity of mud volcanoes (Beciu, Romania) via metagenomic approach. Geomicrobiol. J. 2021, 38, 532–539. [Google Scholar] [CrossRef]
- Hsu, H.-C.; Chen, J.-S.; Nagarajan, V.; Hussain, B.; Huang, S.-W.; Rathod, J.; Hsu, B.-M. Assessment of temporal effects of a mud volcanic eruption on the bacterial community and their predicted metabolic functions in the mud volcanic sites of Niaosong, Southern Taiwan. Microorganisms 2021, 9, 2315. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, V.; Tsai, H.C.; Chen, J.S.; Koner, S.; Kumar, R.S.; Chao, H.C.; Hsu, B.M. Systematic assessment of mineral distribution and diversity of microbial communities and its interactions in the Taiwan subduction zone of mud volcanoes. Environ. Res. 2023, 216, 114536. [Google Scholar] [CrossRef] [PubMed]
- Miyake, N.; Ishimaru, R.; Komatsu, G.; Matsui, T. Characterization of archaeal and bacterial communities thriving in methane-seeping on-land mud volcanoes, Niigata, Japan. Int. Microbiol. 2023, 26, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Mardanov, A.V.; Kadnikov, V.V.; Beletsky, A.V.; Ravin, N.V. Sulfur and methane-oxidizing microbial community in a terrestrial mud volcano revealed by metagenomics. Microorganisms 2020, 8, 1333. [Google Scholar] [CrossRef] [PubMed]
- Merkel, A.Y.; Chernyh, N.A.; Pimenov, N.V.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Diversity and metabolic potential of the terrestrial mud volcano microbial community with a high abundance of archaea mediating the anaerobic oxidation of methane. Life 2021, 11, 953. [Google Scholar] [CrossRef] [PubMed]
- Knittel, K.; Boetius, A. Anaerobic oxidation of methane: Progress with an unknown process. Annu. Rev. Microbiol. 2009, 63, 311–334. [Google Scholar] [CrossRef]
- Haroon, M.F.; Hu, S.; Shi, Y.; Imelfort, M.; Keller, J.; Hugenholtz, P.; Yuan, Z.; Tyson, G.W. Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage. Nature 2013, 500, 567–570. [Google Scholar] [CrossRef]
- Bhattarai, S.; Cassarini, C.; Lens, P.N.L. Physiology and distribution of archaeal methanotrophs that couple anaerobic oxidation of methane with sulfate reduction. Microbiol. Mol. Biol. Rev. 2019, 83, e00074-18. [Google Scholar] [CrossRef]
- Raghoebarsing, A.A.; Pol, A.; Van de Pas-Schoonen, K.T.; Smolders, A.J.; Ettwig, K.F.; Rijpstra, W.I.; Schouten, S.; Damsté, J.S.; den Camp, H.J.; Jetten, M.S.; et al. A microbial consortium couples anaerobic methane oxidation to denitrification. Nature 2006, 440, 918–921. [Google Scholar] [CrossRef]
- Beal, E.J.; House, C.H.; Orphan, V.J. Manganese-and iron-dependent marine methane oxidation. Science 2009, 325, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Leu, A.O.; Xie, G.J.; Guo, J.; Feng, Y.; Zhao, J.X.; Tyson, G.W.; Yuan, Z.; Hu, S. A methanotrophic archaeon couples anaerobic oxidation of methane to Fe (III) reduction. ISME J. 2018, 12, 1929–1939. [Google Scholar] [CrossRef]
- Leu, A.O.; Cai, C.; McIlroy, S.J.; Southam, G.; Orphan, V.J.; Yuan, Z.; Hu, S.; Tyson, G.W. Anaerobic methane oxidation coupled to manganese reduction by members of the Methanoperedenaceae. ISME J. 2020, 14, 1030–1041. [Google Scholar] [CrossRef]
- Zhang, X.; Joyce, G.H.; Leu, A.O.; Zhao, J.; Rabiee, H.; Virdis, B.; Tyson, G.W.; Yuan, Z.; McIlroy, S.J.; Hu, S. Multi-heme cytochrome-mediated extracellular electron transfer by the anaerobic methanotroph ‘Candidatus Methanoperedens nitroreducens’. Nat. Commun. 2023, 14, 6118. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, L.; Holler, T.; Knittel, K.; Meyerdierks, A.; Amann, R. Identification of the dominant sulfate-reducing bacterial partner of anaerobic methanotrophs of the ANME-2 clade. Environ. Microbiol. 2010, 12, 2327–2340. [Google Scholar] [CrossRef]
- Murali, R.; Yu, H.; Speth, D.R.; Wu, F.; Metcalfe, K.S.; Crémière, A.; Laso-Pèrez, R.; Malmstrom, R.R.; Goudeau, D.; Woyke, T.; et al. Physiological potential and evolutionary trajectories of syntrophic sulfate-reducing bacterial partners of anaerobic methanotrophic archaea. PLoS Biol. 2023, 21, e3002292. [Google Scholar] [CrossRef]
- Wegener, G.; Krukenberg, V.; Ruff, S.E.; Kellermann, M.Y.; Knittel, K. Metabolic capabilities of microorganisms involved in and associated with the anaerobic oxidation of methane. Front. Microbiol. 2016, 7, 869. [Google Scholar] [CrossRef]
- Skennerton, C.T.; Chourey, K.; Iyer, R.; Hettich, R.L.; Tyson, G.W.; Orphan, V.J. Methane-fueled syntrophy through extracellular electron transfer: Uncovering the genomic traits conserved within diverse bacterial partners of anaerobic methanotrophic archaea. mBio 2017, 8, e00530-17. [Google Scholar] [CrossRef] [PubMed]
- Krukenberg, V.; Harding, K.; Richter, M.; Glöckner, F.O.; Gruber-Vodicka, H.R.; Adam, B.; Berg, J.S.; Knittel, K.; Tegetmeyer, H.E.; Boetius, A.; et al. Candidatus Desulfofervidus auxilii, a hydrogenotrophic sulfate-reducing bacterium involved in the thermophilic anaerobic oxidation of methane. Environ. Microbiol. 2016, 8, 3073–3091. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, Y.; Lee, H.-S.; Jin, P. Significance of anaerobic oxidation of methane (AOM) in mitigating methane emission from major natural and anthropogenic sources: A review of AOM rates in recent publications. Environ. Sci. Adv. 2022, 1, 401–425. [Google Scholar] [CrossRef]
- Lavrushin, V.Y.; Kopf, A.; Dehle, A.; Stepanets, M.I. Formation of mudvolcanic fluids in Taman (Russia) and Kakhetia (Georgia): Evidence from boron isotopes. Lithol. Miner. Resour. 2003, 38, 120–153. [Google Scholar] [CrossRef]
- Khomyakova, M.A.; Merkel, A.Y.; Kopitsyn, D.S.; Slobodkin, A.I. Pelovirga terrestris gen. nov., sp. nov., anaerobic, alkaliphilic, fumarate-, arsenate-, Fe(III)- and sulfur-reducing bacterium isolated from a terrestrial mud volcano. System. Appl. Microbiol. 2022, 45, 126304. [Google Scholar] [CrossRef] [PubMed]
- Khomyakova, M.A.; Merkel, A.Y.; Segliuk, V.S.; Slobodkin, A.I. Desulfatitalea alkaliphila sp. nov., an alkalipilic sulfate- and arsenate- reducing bacterium isolated from a terrestrial mud volcano. Extremophiles 2023, 27, 12. [Google Scholar] [CrossRef] [PubMed]
- Slobodkina, G.B.; Merkel, A.Y.; Novikov, A.A.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Pelomicrobium methylotrophicum gen. nov., sp. nov. a moderately thermophilic, facultatively anaerobic, lithoautotrophic and methylotrophic bacterium isolated from a terrestrial mud volcano. Extremophiles 2020, 24, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Slobodkina, G.; Ratnikova, N.; Merkel, A.; Kevbrin, V.; Kuchierskaya, A.; Slobodkin, A. Lithoautotrophic lifestyle of the widespread genus Roseovarius revealed by physiological and genomic characterization of Roseovarius autotrophicus sp. nov. FEMS Microbiol. Ecol. 2022, 98, fiac113. [Google Scholar] [CrossRef]
- Slobodkina, G.; Merkel, A.; Ratnikova, N.; Kuchierskaya, A.; Slobodkin, A. Sedimenticola hydrogenitrophicus sp. nov. a chemolithoautotrophic bacterium isolated from a terrestrial mud volcano, and proposal of Sedimenticolaceae fam. nov. in the order. Chromatiales. System. Appl. Microbiol. 2023, 46, 126451. [Google Scholar] [CrossRef]
- Rusanov, I.I.; Savvichev, A.S.; Yusupov, S.K.; Pimenov, N.V.; Ivanov, M.V. Production of exometabolites in the microbial oxidation of methane in ma-rine ecosystems. Microbiology 1998, 67, 590–596. [Google Scholar]
- Stroeva, A.R.; Klyukina, A.A.; Vidishcheva, O.N.; Poludetkina, E.N.; Solovyeva, M.A.; Pyrkin, V.O.; Gavirova, L.A.; Birkeland, N.-K.; Akhmanov, G.G.; Bonch-Osmolovskaya, E.A.; et al. Structure of benthic microbial communities in the Northeastern part of the Barents Sea. Microorganisms 2024, 12, 387. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Pimenov, N.V.; Savvichev, A.S.; Rusanov, I.I.; Lein, A.Y.; Ivanov, M.V. Microbiological processes of the carbon and sulfur cycle on cold methane seeps of the North Atlantic. Microbiology 2000, 69, 831–843. [Google Scholar]
- Ruff, S.E.; Biddle, J.F.; Teske, A.P.; Knittel, K.; Boetius, A.; Ramette, A. Global dispersion and local diversification of the methane seep microbiome. Proc. Natl. Acad. Sci. USA 2015, 112, 4015–4020. [Google Scholar] [CrossRef]
- Tavormina, P.L.; Hatzenpichler, R.; McGlynn, S.; Chadwick, G.; Dawson, K.S.; Connon, S.A.; Orphan, V.J. Methyloprofundus sedimenti gen. nov., sp. nov., an obligate methanotroph from ocean sediment belonging to the ‘deep sea-1’ clade of marine methanotrophs. Int. J. Syst. Evol. Microbiol. 2015, 65, 251–259. [Google Scholar] [CrossRef]
- Hirayama, H.; Takaki, Y.; Abe, M.; Imachi, H.; Ikuta, T.; Miyazaki, J.; Tasumi, E.; Uematsu, K.; Tame, A.; Tsuda, M.; et al. Multispecies populations of methanotrophic Methyloprofundus and cultivation of a likely dominant species from the Iheya North deep-sea hydrothermal field. Appl. Environ. Microbiol. 2022, 88, e0075821. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, G.L.; Skennerton, C.T.; Laso-Perez, R.; Leu, A.O.; Speth, D.R.; Yu, H.; Morgan-Lang, C.; Hatzenpichler, R.; Goudeau, D.; Malmstrom, R.; et al. Comparative genomics reveals electron transfer and syntrophic mechanisms differentiating methanotrophic and methanogenic archaea. PLoS Biol. 2022, 20, e3001508. [Google Scholar] [CrossRef] [PubMed]
- Khomyakova, M.A.; Zavarzina, D.G.; Merkel, A.Y.; Klyukina, A.A.; Pikhtereva, V.A.; Gavrilov, S.N.; Slobodkin, A.I. The first cultivated representatives of the actinobacterial lineage OPB41 isolated from subsurface environments constitute a novel order Anaerosomatales. Front. Microbiol. 2022, 13, 1047580. [Google Scholar] [CrossRef]
- Wunder, L.C.; Aromokeye, D.A.; Yin, X.; Richter-Heitmann, T.; Willis-Poratti, G.; Schnakenberg, A.; Otersen, C.; Dohrmann, I.; Römer, M.; Bohrmann, G.; et al. Iron and sulfate reduction structure microbial communities in (sub-)Antarctic sediments. ISME J. 2021, 12, 3587–3604. [Google Scholar] [CrossRef]
- Wang, L.; Shao, Z. Aerobic denitrification and heterotrophic sulfur oxidation in the genus Halomonas revealed by six novel species characterizations and genome-based analysis. Front. Microbiol. 2021, 12, 652766. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D.; Rentz, J.A.; Lilburn, T.G.; Davis, R.E.; Aldrich, H.; Chan, C.; Moyer, C.L. A novel lineage of Proteobacteria involved in formation of marine Fe-oxidizing microbial mat communities. PLoS ONE 2007, 2, e667. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Timmers, P.H.A.; Welte, C.U.; Koehorst, J.J.; Plugge, C.M.; Jetten, M.S.M.; Stams, A.J.M. Reverse methanogenesis and respiration in methanotrophic archaea. Archaea 2017, 2017, 1654237. [Google Scholar] [CrossRef]
- Mehta, P.; Mishra, S.; Ghose, T. Methanol accumulation by resting cells of Methylosinus trichosporium (I). J. Gen. Appl. Microbiol. 1987, 33, 221–229. [Google Scholar] [CrossRef]
- Farr, O.; Gaudu, N.; Danger, G.; Russell, M.J.; Ferry, D.; Nitschke, W.; Duval, S. Methanol on the rocks: Green rust transformation promotes the oxidation of methane. J. R. Soc. Interface 2023, 206, 20230386. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A. Ecological aspects of the distribution of different autotrophic CO2 fixation pathways. Appl. Environ. Microbiol. 2011, 77, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Nunoura, T.; Chikaraishi, Y.; Izaki, R.; Suwa, T.; Sato, T.; Harada, T.; Mori, K.; Kato, Y.; Miyazaki, M.; Shimamura, S.; et al. A primordial and reversible TCA cycle in a facultatively chemolithoautotrophic thermophile. Science 2018, 359, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Mall, A.; Sobotta, J.; Huber, C.; Tschirner, C.; Kowarschik, S.; Bačnik, K.; Mergelsberg, M.; Boll, M.; Hügler, M.; Eisenreich, W.; et al. Reversibility of citrate synthase allows autotrophic growth of a thermophilic bacterium. Science 2018, 359, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Andrea, I.; Guedes, I.A.; Hornung, B.; Boeren, S.; Lawson, C.E.; Sousa, D.Z.; Bar-Even, A.; Claassens, N.J.; Stams, A.J.M. The reductive glycine pathway allows autotrophic growth of Desulfovibrio desulfuricans. Nat. Commun. 2020, 11, 5090. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, R.S.; Watmough, N.J. The bacterial cytochrome cbb3 oxidases. Biochim. Biophys. Acta 2004, 1655, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Tourova, T.P.; Mussmann, M.; Muyzer, G. Dethiobacter alkaliphilus gen. nov. sp. nov., and Desulfurivibrio alkaliphilus gen. nov. sp. nov.: Two novel representatives of reductive sulfur cycle from soda lakes. Extremophiles 2008, 12, 431–439. [Google Scholar]
- Flood, B.E.; Jones, D.S.; Bailey, J.V. Sedimenticola thiotaurini sp. nov., a sulfur-oxidizing bacterium isolated from salt marsh sediments, and emended descriptions of the genus Sedimenticola and Sedimenticola selenatireducens. Int. J. Syst. Evol. Microbiol. 2015, 65, 2522–2530. [Google Scholar] [CrossRef]
- Frigaard, N.U.; Dahl, C. Sulfur metabolism in phototrophic sulfur bacteria. Adv. Microb. Physiol. 2009, 54, 103–200. [Google Scholar] [CrossRef] [PubMed]
- de Zwart, J.M.M.; Nelisse, P.N.; Kuenen, J.G. Isolation and characterization of Methylophaga sulfidovorans sp. nov.: An obligately methylotrophic, aerobic, dimethylsulfide oxidizing bacterium from microbial mat. FEMS Microbiol. Ecol. 1996, 20, 261–270. [Google Scholar] [CrossRef]
- Boden, R.; Kelly, D.P.; Murrell, J.C.; Schafer, H. Oxidation of dimethylsulfide to tetrathionate by Methylophaga thiooxidans sp. nov.: A new link in the sulfur cycle. Environ. Microbiol. 2010, 12, 2688–2699. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, B.; Schicklberger, M.; Kuermann, J.; Spormann, A.M.; Gescher, J. Periplasmic electron transfer via the c-type cytochromes MtrA and FccA of Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2009, 75, 7789–7796. [Google Scholar] [CrossRef] [PubMed]
- Frolova, A.A.; Merkel, A.Y.; Kevbrin, V.V.; Kopitsyn, D.S.; Slobodkin, A.I. Sulfurospirillum tamanensis sp. nov., a facultatively anaerobic alkaliphilic bacterium from a terrestrial mud volcano. Microbiology 2023, 92, 21–29. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample # | Volcano | Sampling Site | pH | Salinity, g L−1 | Eh, mV | CH4, mM | Cl−, mM | PO4−, mM | SO42−, mM | NO3−, mM |
|---|---|---|---|---|---|---|---|---|---|---|
| C01 | Gnilaya Gora | Gryphon | 8.47 | 20 | −295 | 101.1 | 241.1 | 0.00 | 5.79 | 0.00 |
| C02 | Gnilaya Gora | Gryphon | 8.66 | 14 | −175 | 556.7 | 158.8 | 0.07 | 1.07 | 0.00 |
| C03 | Gnilaya Gora | Salsa | 9.10 | 20 | −320 | 201.1 | 412.0 | 0.00 | 0.00 | 0.00 |
| C04 | Gnilaya Gora | Mud pool | 8.64 | 12 | −348 | 536.3 | 162.8 | 0.02 | 0.09 | 0.92 |
| C05 | Gnilaya Gora | Mud pool | 8.94 | 20 | −302 | 47.7 | 216.1 | 0.00 | 1.95 | 0.00 |
| C10 | Gladkovsky | Mud pool | 6.75 | 20 | −124 | 668.0 | 307.5 | 0.00 | 0.06 | 0.00 |
| C11 | Gladkovsky | Mud pool | 6.72 | 20 | −110 | 661.5 | 300.8 | 0.00 | 0.08 | 0.00 |
| C12 | Gladkovsky | Mud pool | 7.07 | 30 | 15 | 3.3 | 402.2 | 0.00 | 0.00 | 0.00 |
| C13 | Gladkovsky | Mud pool | 7.08 | 30 | −255 | 105.9 | 393.4 | 0.00 | 0.00 | 0.00 |
| C14 | Gladkovsky | Mud pool | 7.02 | 20 | −217 | 439.1 | 291.2 | 0.00 | 0.00 | 0.00 |
| C16 | Kuchugursky | Main crater | 7.43 | 15 | −212 | 640.5 | 153.9 | 0.00 | 8.54 | 0.00 |
| C17 | Kuchugursky | Mud pool | 6.92 | 20 | nd | nd | 147.4 | 0.00 | 63.52 | 0.00 |
| C19 | Semigorsky | Mud pool | 8.20 | 10 | nd | nd | 5.7 | 0.00 | 0.08 | 0.00 |
| C20 | Semigorsky | Mud pool | 8.35 | 9 | nd | nd | 37.2 | 0.00 | 0.02 | 0.00 |
| C21 | Shugo | Gryphon | 7.23 | 20 | −196 | 456.8 | 199.2 | 0.00 | 0.20 | 0.00 |
| C23 | Shugo | Gryphon | 6.94 | 12 | nd | nd | 95.4 | 0.00 | 0.21 | 0.00 |
| C24 | Shugo | Gryphon | 7.41 | 20 | −157 | 917.3 | 193.7 | 0.00 | 0.21 | 0.00 |
| Bin ID | Domain | Taxon * | Abundance % | Completeness % | Contamination % | # Contigs | Genome Size, Mbp | 16S rRNA Gene, bp |
|---|---|---|---|---|---|---|---|---|
| C14-01 | B | c_Acidimicrobiia | 19.04 | 100.00 | 2.14 | 181 | 3.91 | 1171 |
| C14-19 | B | d_Bacteria | 18.57 | 94.78 | 0.84 | 157 | 3.19 | 1558 |
| C14-17 | A | f_Ca. Methanocomedenaceae | 14.23 | 95.42 | 0.00 | 131 | 2.06 | - |
| C14-08 | A | f_Ca. Methanocomedenaceae | 11.51 | 98.69 | 0.00 | 74 | 1.82 | - |
| C14-05 | B | g_Methyloprofundus | 9.49 | 99.31 | 1.55 | 189 | 3.28 | - |
| C14-10 | B | o_Bacteroidales | 3.13 | 97.85 | 1.90 | 50 | 3.48 | 1533 |
| C14-03 | B | f_Methylophagaceae | 2.56 | 99.82 | 0.65 | 96 | 2.44 | - |
| C14-15 | B | d_Bacteria | 2.38 | 96.40 | 0.00 | 255 | 2.72 | - |
| C14-28 | B | p_Paceibacteria | 2.28 | 76.85 | 1.12 | 68 | 1.17 | 1474 |
| C14-27 | B | o_Anaerolineales | 2.21 | 80.18 | 2.73 | 99 | 2.69 | 1517 |
| C14-06 | B | o_Bacteroidales | 1.81 | 99.28 | 1.90 | 74 | 3.81 | - |
| C14-21 | B | g_Brevefilum | 1.27 | 90.00 | 2.73 | 104 | 2.66 | 1522 |
| C14-02 | B | o_Desulfuromonadales | 1.10 | 100.00 | 0.22 | 118 | 2.72 | - |
| C14-14 | B | c_Myxococcia | 1.04 | 96.55 | 2.60 | 75 | 6.11 | 768 |
| C14-09 | B | o_Bacteroidales | 1.00 | 98.57 | 1.19 | 119 | 4.61 | - |
| C14-29 | B | p_Patescibacteria | 0.90 | 73.76 | 0.99 | 49 | 1.46 | 1512 |
| C14-13 | B | o_Anaerosomatales | 0.79 | 96.66 | 3.61 | 52 | 1.98 | 432 |
| C14-04 | B | o_Desulfobacterales | 0.78 | 99.35 | 2.26 | 269 | 4.94 | 556 |
| C14-16 | B | o_Deferrisomatales | 0.70 | 95.70 | 2.69 | 470 | 5.78 | - |
| C14-07 | B | f_Desulfurivibrionaceae | 0.63 | 98.80 | 2.35 | 287 | 2.52 | - |
| C14-12 | B | p_Zixibacteria | 0.46 | 96.70 | 1.20 | 242 | 3.06 | - |
| C14-18 | A | g_Ca. Methanovorans | 0.44 | 95.01 | 0.98 | 263 | 1.88 | - |
| C14-11 | B | f_Spirochaetaceae | 0.43 | 97.20 | 2.00 | 344 | 3.68 | - |
| C14-24 | B | f_Trueperaceae | 0.40 | 84.95 | 2.97 | 666 | 3.26 | 696 |
| C14-20 | A | f_Thorarchaeaceae | 0.39 | 90.18 | 3.27 | 703 | 4.03 | 1556 |
| C14-25 | B | o_Bacteroidales | 0.36 | 83.63 | 3.83 | 1235 | 6.00 | - |
| C14-23 | B | g_Lutibacter | 0.34 | 85.01 | 3.24 | 674 | 2.49 | - |
| C14-22 | B | g_Sedimenticola | 0.34 | 85.63 | 1.39 | 663 | 3.03 | - |
| C14-26 | B | c_Anaerolineae | 0.33 | 80.49 | 0.48 | 1021 | 4.22 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slobodkin, A.I.; Rusanov, I.I.; Slobodkina, G.B.; Stroeva, A.R.; Chernyh, N.A.; Pimenov, N.V.; Merkel, A.Y. Diversity, Methane Oxidation Activity, and Metabolic Potential of Microbial Communities in Terrestrial Mud Volcanos of the Taman Peninsula. Microorganisms 2024, 12, 1349. https://doi.org/10.3390/microorganisms12071349
Slobodkin AI, Rusanov II, Slobodkina GB, Stroeva AR, Chernyh NA, Pimenov NV, Merkel AY. Diversity, Methane Oxidation Activity, and Metabolic Potential of Microbial Communities in Terrestrial Mud Volcanos of the Taman Peninsula. Microorganisms. 2024; 12(7):1349. https://doi.org/10.3390/microorganisms12071349
Chicago/Turabian StyleSlobodkin, Alexander I., Igor I. Rusanov, Galina B. Slobodkina, Aleksandra R. Stroeva, Nikolay A. Chernyh, Nikolai V. Pimenov, and Alexander Y. Merkel. 2024. "Diversity, Methane Oxidation Activity, and Metabolic Potential of Microbial Communities in Terrestrial Mud Volcanos of the Taman Peninsula" Microorganisms 12, no. 7: 1349. https://doi.org/10.3390/microorganisms12071349