
Effects of Dietary Multi-Strain Probiotics on Growth Performance, Antioxidant Status, Immune Response, and Intestinal Microbiota of Hybrid Groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Feeding Conditions
2.2. Preparation of Probiotics and Feed
2.3. Sample Collection
2.4. Growth Performance
2.5. Determination of Enzymes Related to Digestion and Immune Function
2.6. Gene Expression Detection
2.7. Analysis of Intestinal Microbial Community
2.8. Statistical Analysis
3. Results
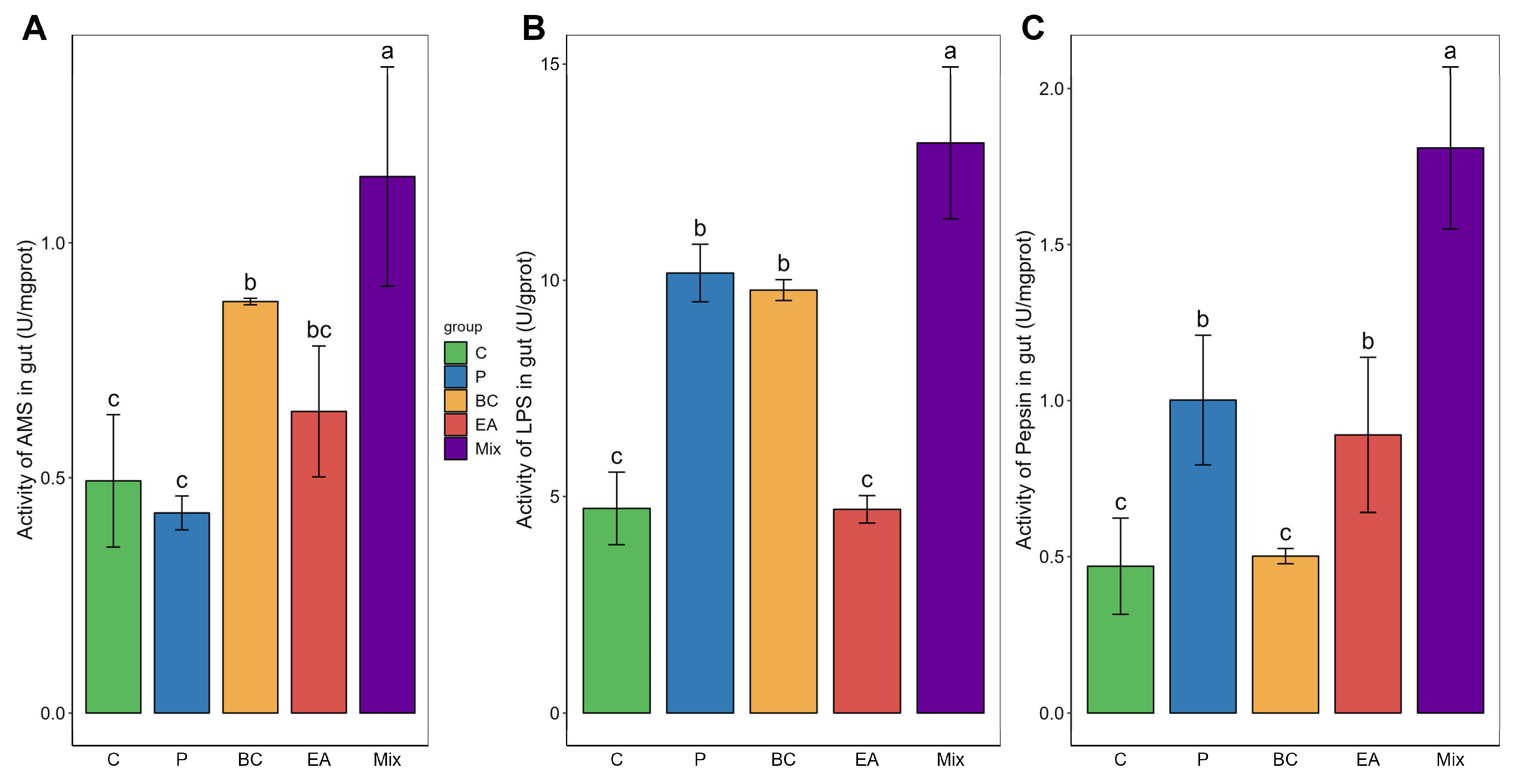
3.1. Growth Performance and Digestive Enzyme Activity
3.2. mRNA Expression Levels of Growth-Related Genes
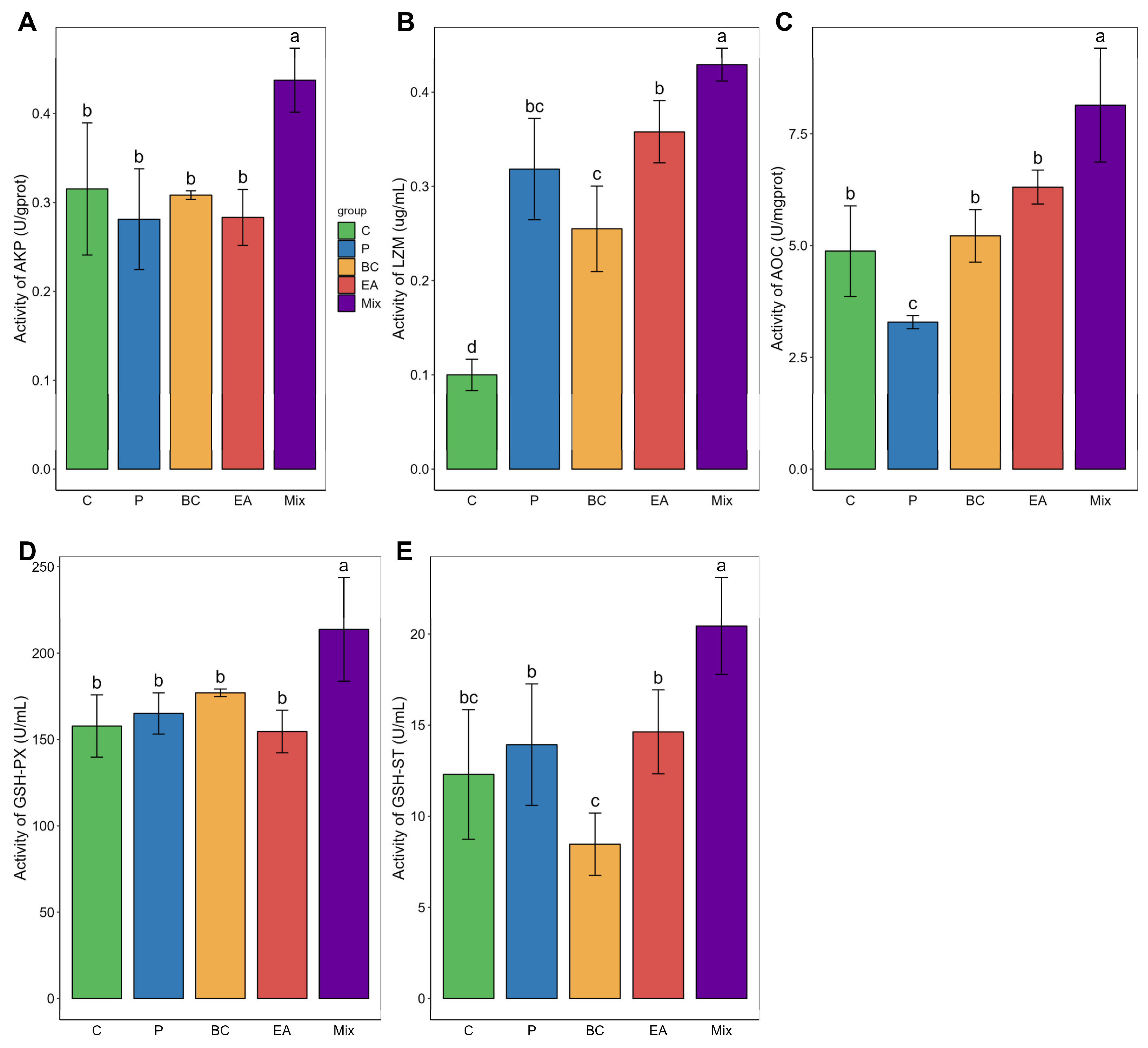
3.3. Immune Response and Antioxidant Activity
3.4. mRNA Expression Levels of Immune and Antioxidant Genes
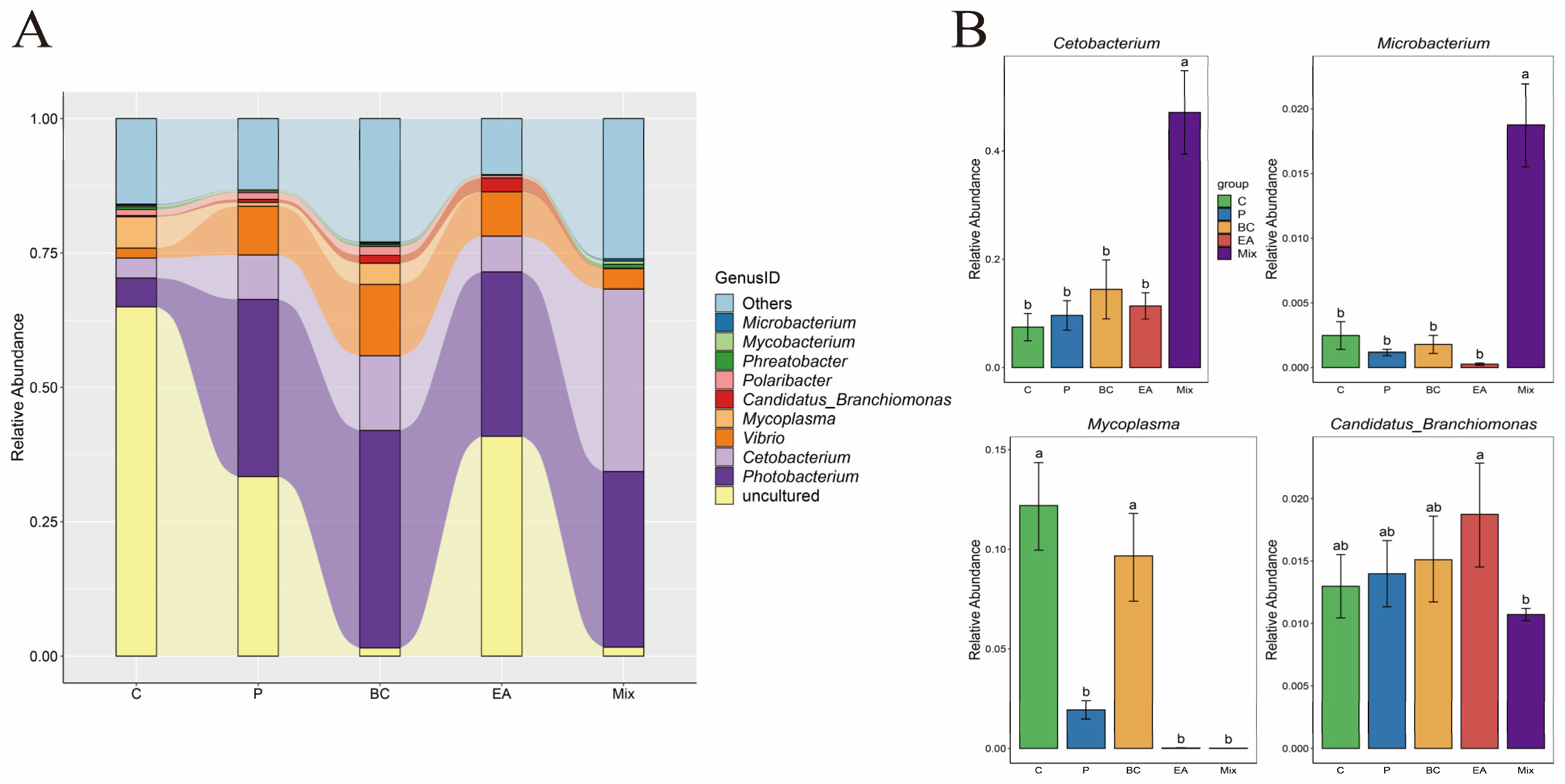
3.5. Effect of Probiotic Treatment on the Intestinal Microbiota of the Hybrid Groupers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: A review of dietary interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Opiyo, M.A.; Marijani, E.; Muendo, P.; Odede, R.; Leschen, W.; Charo-Karisa, H. A review of aquaculture production and health management practices of farmed fish in Kenya. Int. J. Vet. Sci. Med. 2018, 6, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.-F. Chinese wisdom and modern innovation of aquaculture. Water Biol. Secur. 2024, 100271. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish. Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.R.; Chaklader, M.R.; Shukry, M.; Ahmed, H.A.; Khallaf, M.A. A multispecies probiotic modulates growth, digestive enzymes, immunity, hepatic antioxidant activity, and disease resistance of Pangasianodon hypophthalmus fingerlings. Aquaculture 2023, 563, 738948. [Google Scholar] [CrossRef]
- Kong, Y.; Li, M.; Chu, G.; Liu, H.; Shan, X.; Wang, G.; Han, G. The positive effects of single or conjoint administration of lactic acid bacteria on Channa argus: Digestive enzyme activity, antioxidant capacity, intestinal microbiota and morphology. Aquaculture 2021, 531, 735852. [Google Scholar] [CrossRef]
- Ringø, E.; Van Doan, H.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, lactic acid bacteria and bacilli: Interesting supplementation for aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M. Impact of Probiotics on Colonizing Microbiota of the Gut. J. Clin. Gastroenterol. 2011, 45, S115–S119. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Liu, P.; Li, Y.; Song, D.; Long, W.; Wang, Z.; Yi, S.; Jiang, L. Gut microbiota, host genetics and phenotypes in aquatic animals: A review. Aquac. Rep. 2023, 31, 101648. [Google Scholar] [CrossRef]
- Xie, G.; Zhu, Y.; Zhong, Z.; Du, Q.; Wu, Y.; Xing, K.; Zhang, M.; Shu, H. Functional genomic characterization unveils probiotic features of Bacillus cereus G1-11 isolated from the gut of the hybrid grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂). LWT 2023, 184, 115088. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, A.; Mann, D. The Synergic Effect of Gut-Derived Probiotic Bacillus cereus SL1 And Ocimum sanctum on Growth, Intestinal Histopathology, Innate Immunity, and Expression of Enzymatic Antioxidant Genes in Fish, Cirrhinus mrigala (Hamilton, 1822). Probiotics Antimicrob. Proteins. 2023; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Jinendiran, S.; Nathan, A.A.; Ramesh, D.; Vaseeharan, B.; Sivakumar, N. Modulation of innate immunity, expression of cytokine genes and disease resistance against Aeromonas hydrophila infection in goldfish (Carassius auratus) by dietary supplementation with Exiguobacterium acetylicum S01. Fish Shellfish Immunol. 2019, 84, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Zan, Z.; Chen, K.; Wang, H.; Han, Z.; Sun, J. Effects of a multistrain probiotic on the growth, immune function and intestinal microbiota of the tongue sole Cynoglossus semilaevis. Aquaculture 2023, 575, 739813. [Google Scholar] [CrossRef]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.-M.; Kim, H.K.; Kim, O.-J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Shadrack, R.S.; Manabu, I.; Koshio, S.; Yokoyama, S.; Zhang, Y.; Mzengereza, K.; El Basuini, M.F.; Dawood, M.A.O. Effects of Single and Mixture Probiotic Supplements on Growth, Digestive Activity, Antioxidative Status, Immune and Growth-Related Genes, and Stress Response of Juvenile Red Sea Bream (Pagrus Major). Aquac. Nutr. 2022, 8968494. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Wang, Z.; Lu, Y.; Abarike, E.D.; Sakyi, M.E.; Li, Y.; Xie, C.X.; Hlordzi, V. Effects of three host-associated Bacillus species on mucosal immunity and gut health of Nile tilapia, Oreochromis niloticus and its resistance against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2020, 97, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-Q.; Jiang, C.; Ling, F.; Wang, G.-X. Effects of dietary supplementation of intestinal autochthonous bacteria on the innate immunity and disease resistance of grass carp (Ctenopharyngodon idellus). Aquaculture 2015, 438, 105–114. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, X.; Wu, X.; Gao, Y.; Li, X.; Dong, Y.; Yao, W. Effects of dietary leucine levels on growth, feed utilization, neuro-endocrine growth axis and TOR-related signaling genes expression of juvenile hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus ♂). Aquaculture 2019, 504, 172–181. [Google Scholar] [CrossRef]
- Yang, H.-L.; Sun, Y.-Z.; Hu, X.; Ye, J.-D.; Lu, K.-L.; Hu, L.-H.; Zhang, J.-J. Bacillus pumilus SE5 originated PG and LTA tuned the intestinal TLRs/MyD88 signaling and microbiota in grouper (Epinephelus coioides). Fish Shellfish Immunol. 2019, 88, 266–271. [Google Scholar] [CrossRef]
- Taj, S.; Ma, L.; Wu, X.; Ye, B.; Geng, L.; Zhou, Z.; Wang, X.; Mu, W. Effects of Dietary Histidine Levels on Growth Performance, Feed Utilization, and Expression of Related Genes of Juvenile Hybrid Grouper Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂. Aquac. Nutr. 2022, 2022, 7738843. [Google Scholar] [CrossRef]
- Yang, X.; Zhi, X.; Song, Z.; Wang, G.; Zhao, X.; Chi, S.; Tan, B. Flesh quality of hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂) fed with hydrolyzed porcine mucosa-supplemented low fishmeal diet. Anim. Nutr. 2022, 8, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, D.; Mao, M.; Wu, J.; Yang, Q.; Tan, B.; Chi, S. Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper. Metabolites 2022, 12, 1268. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-W.; Cai, L.; Qi, Z.-H.; Wang, C.; Liu, Y.; Wang, W.-N. Effects of a recombinant complement component C3b functional fragment α2MR (α2-macroglobulin receptor) additive on the immune response of juvenile orange-spotted grouper (Epinephelus coioides) after the exposure to cold shock challenge. Fish Shellfish Immunol. 2015, 45, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liu, H.; Yang, S.; Zhou, M.; Zhang, S.; Tan, B.; Yang, Y.; Zhang, H.; Xie, R.; Dong, X. Effects of High-Lipid Dietary Protein Ratio on Growth, Antioxidant Parameters, Histological Structure, and Expression of Antioxidant- and Immune-Related Genes of Hybrid Grouper. Animals 2023, 13, 3710. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wu, X.; Lu, S.; Gao, Y.; Yao, W.; Li, X.; Dong, Y.; Jin, Z. Dietary arginine affects growth, gut morphology, oxidation resistance and immunity of hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂) juveniles. Br. J. Nutr. 2018, 120, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Xun, P.; Huang, J.; Jiang, K.; Huang, X.; Yu, W.; Xu, C.; Lin, H. Effects of dietary Clostridium butyricum on the growth performance and intestinal health of hybrid groupers (Epinephelus lanceolatus♂ × E. fuscoguttatus♀). Aquac. Rep. 2023, 30, 101588. [Google Scholar] [CrossRef]
- Xu, J.-M.; Cai, G.-H.; Li, J.; Yang, H.-L.; Ye, J.-D.; Sun, Y.-Z. Commensal Bacillus pumilus SE5-Derived Peptidoglycan and Lipoteichoic Acid Showed Synergistic Effects in Improving Growth, Immunity, and Intestinal Health of Grouper (Epinephelus coioides). Probiotics Antimicrob. Proteins 2024, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zhu, Z.; Ge, H.; Zheng, L.; Huang, Z.; Wu, S. Immunity to nervous necrosis virus infections of orange-spotted grouper (Epinephelus coioides) by vaccination with virus-like particles. Fish Shellfish Immunol. 2016, 56, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Cao, H.; Jiang, W.; Hu, B.; Jian, S.; Wen, C.; Kajbaf, K.; Kumar, V.; Tao, Z.; Peng, M. Dietary supplementation of Bacillus cereus as probiotics in Pengze crucian carp (Carassius auratus var. Pengze): Effects on growth performance, fillet quality, serum biochemical parameters and intestinal histology. Aquac. Res. 2019, 50, 2207–2217. [Google Scholar] [CrossRef]
- Yang, G.; Shen, K.; Yu, R.; Wu, Q.; Yan, Q.; Chen, W.; Ding, L.; Kumar, V.; Wen, C.; Peng, M. Probiotic (Bacillus cereus) enhanced growth of Pengze crucian carp concurrent with modulating the antioxidant defense response and exerting beneficial impacts on inflammatory response via Nrf2 activation. Aquaculture 2020, 529, 735691. [Google Scholar] [CrossRef]
- Chen, X.; Yi, H.; Liu, S.; Zhang, Y.; Su, Y.; Liu, X.; Bi, S.; Lai, H.; Zeng, Z.; Li, G. Promotion of pellet-feed feeding in mandarin fish (Siniperca chuatsi) by Bdellovibrio bacteriovorus is influenced by immune and intestinal flora. Aquaculture 2021, 542, 736864. [Google Scholar] [CrossRef]
- Bernal, M.G.; Marrero, R.M.; Campa-Córdova, Á.I.; Mazón-Suástegui, J.M. Probiotic effect of Streptomyces strains alone or in combination with Bacillus and Lactobacillus in juveniles of the white shrimp Litopenaeus vannamei. Aquac. Int. 2017, 25, 927–939. [Google Scholar] [CrossRef]
- Ma, S.; Yu, D.; Liu, Q.; Zhao, M.; Xu, C.; Yu, J. Relationship between immune performance and the dominant intestinal microflora of turbot fed with different Bacillus species. Aquaculture 2022, 549, 737625. [Google Scholar] [CrossRef]
- Essa, M.A.; Elserafy, S.; Elezaby, M.; Daboor, S.; Esmael, N.A.; Lall, S.P. Effect of different dietary probiotics on growth, feed utilization and digestive enzymes activities of Nile tilapia, Oreochromis niloticus. J. Arab. Aquac. Soc. 2010, 5, 143–162. [Google Scholar]
- Siddik, M.A.B.; Foysal, M.J.; Fotedar, R.; Francis, D.S.; Gupta, S.K. Probiotic yeast Saccharomyces cerevisiae coupled with Lactobacillus casei modulates physiological performance and promotes gut microbiota in juvenile barramundi, Lates calcarifer. Aquaculture 2022, 546, 737346. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Tang, J.; Cai, J.; Yu, H.; Wang, Z.; Abarike, E.D.; Lu, Y.; Li, Y.; Afriyie, G. In vivo assessment of the probiotic potentials of three host-associated Bacillus species on growth performance, health status and disease resistance of Oreochromis niloticus against Streptococcus agalactiae. Aquaculture 2020, 527, 735440. [Google Scholar] [CrossRef]
- Picha, M.E.; Turano, M.J.; Beckman, B.R.; Borski, R.J. Endocrine Biomarkers of Growth and Applications to Aquaculture: A Minireview of Growth Hormone, Insulin-Like Growth Factor (IGF)-I, and IGF-Binding Proteins as Potential Growth Indicators in Fish. N. Am. J. Aquac. 2008, 70, 196–211. [Google Scholar] [CrossRef]
- Pierce, A.L.; Breves, J.P.; Moriyama, S.; Uchida, K.; Grau, E.G. Regulation of growth hormone (GH) receptor (GHR1 and GHR2) mRNA level by GH and metabolic hormones in primary cultured tilapia hepatocytes. Gen. Comp. Endocrinol. 2012, 179, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Funkenstein, B.; Skopal, T.; Rapoport, B.; Rebhan, Y.; Du, S.J.; Radaelli, G. Characterization and functional analysis of the 5’ flanking region of myosin light chain-2 gene expressed in white muscle of the gilthead sea bream (Sparus aurata). Comp. Biochem. Physiol. Part D Genom. Proteom. 2007, 2, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, Z.; Liu, Z.; Shen, J.; Feng, H.; Xue, L. Transgenic red carp (Cyprinus carpio) with LcMSTN1 propeptide: Enhanced growth and unchanged muscle fat content. Aquaculture 2021, 541, 736795. [Google Scholar] [CrossRef]
- Meidong, R.; Buatong, A.; Nakao, M.; Sakai, K.; Tongpim, S. Mixed culture of Bacillus aerius B81e and Lactiplantibacillus paraplantarum L34b-2 derived from in vivo screening using hybrid catfish exhibits high probiotic effects on Pangasius bocourti. J. Biosci. Bioeng. 2021, 132, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, G.; Vaseeharan, B.; Chen, J.-C.; Rekha, R.; Anjugam, M.; Arokiadhas, I. Dietary supplementation of probiotic Bacillus licheniformis Dahb1 improves growth performance, mucus and serum immune parameters, antioxidant enzyme activity as well as resistance against Aeromonas hydrophila in tilapia Oreochromis mossambicus. Fish Shellfish Immunol. 2018, 74, 501–508. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y.; Sun, G.; Li, X.; Liu, Z. Growth, immune response, antioxidant capability, and disease resistance of juvenile Atlantic salmon (Salmo salar L.) fed Bacillus velezensis V4 and Rhodotorula mucilaginosa compound. Aquaculture 2019, 500, 65–74. [Google Scholar] [CrossRef]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Yang, S.; Chen, W.; He, F.; Fu, S.; Jin, Z.; Zheng, C.; Zhang, X.; Ye, Z.; Jin, H.; Qian, S.; et al. Comparison of the roles of IgM in systemic and mucosal immunity via tissue distribution analysis in largemouth bass (Micropterus salmoides). Aquaculture 2020, 527, 735488. [Google Scholar] [CrossRef]
- Bilodeau, A.L.; Waldbieser, G.C. Activation of TLR3 and TLR5 in channel catfish exposed to virulent Edwardsiella ictaluri. Dev. Comp. Immunol. 2005, 29, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Haddad, G.; Hanington, P.C.; Wilson, E.C.; Grayfer, L.; Belosevic, M. Molecular and functional characterization of goldfish (Carassius auratus L.) transforming growth factor beta. Dev. Comp. Immunol. 2008, 32, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.-H.; Ye, J.-Y.; Zhang, Y.-X.; Xu, P.; Xie, J. Effects of dietary reduced glutathione on growth performance, non-specific immunity, antioxidant capacity and expression levels of IGF-I and HSP70 mRNA of grass carp (Ctenopharyngodon idella). Aquaculture 2015, 438, 39–46. [Google Scholar] [CrossRef]
- Tan, H.Y.; Chen, S.-W.; Hu, S.-Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 92, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lu, M.; Chen, G.; Cao, J.; Gao, F.; Wang, M.; Liu, Z.; Zhang, D.; Zhu, H.; Yi, M. Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2018, 76, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.M.; Mohammed, H.H.; Arias, C.R. Characterization of the gut microbiota of three commercially valuable warmwater fish species. J. Appl. Microbiol. 2014, 116, 1396–1404. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.; Egan, S. Opportunistic diseases in marine eukaryotes: Could Bacteroidota be the next threat to ocean life? Environ. Microbiol. 2022, 24, 4505–4518. [Google Scholar] [CrossRef] [PubMed]
- Evtushenko, L.I.; Takeuchi, M. The Family Microbacteriaceae. In The Prokaryotes: Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 1020–1098. [Google Scholar]
- Zhang, Y.; Qi, X.; Zhang, Z.; Jin, Z.; Wang, G.; Ling, F. Effects of dietary Cetobacterium somerae on the intestinal health, immune parameters and resistance against Nocardia seriolae of largemouth bass, Micropterus salmoides. Fish Shellfish Immunol. 2023, 135, 108693. [Google Scholar] [CrossRef] [PubMed]
- Mjølnerød, E.B.; Lillebostad, S.M.; Lagadec, E.; Plarre, H.; Moore, L.J.; Trösse, C.; Nylund, A. Host associated genotypes in the family Branchiomonaceae with proposal of Ca. Branchiomonas mykissicola n. sp. from rainbow trout (Oncorhynchus mykiss). Aquaculture 2024, 581, 740260. [Google Scholar] [CrossRef]
- Fu, X.; Huang, L.; Hou, D.; Chen, H.; Chen, C.; Zhang, D.; Guo, H. Sucrose-based biofloc system promotes the enrichment of specific gut bacterial taxa to enhance disease suppression in shrimp (Penaeus vannamei). Aquaculture 2024, 580, 740375. [Google Scholar] [CrossRef]
- Tao, B.; Zhang, C.; Li, X.; Li, X.; Lu, K.; Song, K.; Wang, L. Postbiotics of Bacillus subtilis LCBS1 have beneficial effects on bullfrogs (Lithobates catesbeianus). Aquaculture 2023, 574, 739699. [Google Scholar] [CrossRef]
- Wang, M.; Yi, M.; Lu, M.; Gao, F.; Liu, Z.; Huang, Q.; Li, Q.; Zhu, D. Effects of probiotics Bacillus cereus NY5 and Alcaligenes faecalis Y311 used as water additives on the microbiota and immune enzyme activities in three mucosal tissues in Nile tilapia Oreochromis niloticus reared in outdoor tanks. Aquac. Rep. 2020, 17, 100309. [Google Scholar] [CrossRef]
- Xiong, J.; Dai, W.; Zhu, J.; Liu, K.; Dong, C.; Qiu, Q. The Underlying Ecological Processes of Gut Microbiota Among Cohabitating Retarded, Overgrown and Normal Shrimp. Microb. Ecol. 2017, 73, 988–999. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Diet |
|---|---|
| Fish meal | 450.00 |
| Soybean meal | 130.00 |
| Flour | 237.40 |
| Beer yeast powder | 50.00 |
| Fish oil | 50.00 |
| Soybean oil | 50.00 |
| Lecithin | 10.00 |
| Vitamin premix 1 | 2.00 |
| Mineral premix 2 | 5.00 |
| Choline chloride (50%) | 5.00 |
| Antioxidant | 0.10 |
| Vitamin C | 0.50 |
| Monocalcium phosphate | 10.0 |
| Proximate Composition | |
| Moisture | 5.12 |
| Crude protein | 49.63 |
| Crude lipid | 14.81 |
| Ash | 10.36 |
| Target Genes | Primers Sequences (5′–3′) for qPCR | Amplification Efficiency (%) | Amplicon Size (bp) | Accession No. | Reference |
|---|---|---|---|---|---|
| β-actin | F: GACATCAAGGAGAAGCTGTG | 98 | 214 | AY510710.1 | [20] |
| R: TGCTGTTGTAGGTGGTCTCGT | |||||
| GHR1 | F: CACAGACTTCTATGCCCAGGT R: GTGTAGCCGCTTCCTTCAG | 99 | 144 | KR269817.1 | [21] |
| IGF-1 | F: GTGCGATGTGCTGTATCTCCTG R: GCCATAGCCTGTTGGTTTACTG | 104 | 179 | XM_033614181.1 | [22] |
| IGF-2 | F: CCGCAAAGATACGGACACCA R: GCCGACGCTATTTCCACAAC | 101 | 146 | XM_033635279.1 | [22] |
| MyoD | F: GGCTCAGCAAGGTCAACGA R: CTCGATGTAGCTGATGGCGT | 98 | 113 | XM_033635008.1 | [22] |
| MyoG | F: GAGGAGCACCTTGATGAACCC R: TGGGCTCACTTGAAGACGACA | 102 | 191 | XM_033625713.1 | [22] |
| MyHC | F: AAGGTGTTGGCTGAGTGGAAA R: TGGATGCTCTTGCCAGTCTCA | 96 | 221 | XM_033644959.1 | [22] |
| CAT | F: GCTCTATCCGCTCCTCTTCTCCTC R: GTAGTTCCTGACGACGGTGATGTG | 105 | 115 | KT884509.1 | [23] |
| CTL | F: GGCAAGATGGGAGCCTAAC R: ATGCGACGAATGTCCTGAA | 95 | 121 | FJ805451.1 | [24] |
| SOD | F: GTTGGAGACCTGGGAAATGTGACTG R: CCATTGAGGGTGAGCATCTTGTCC | 102 | 83 | AY735008.1 | [23] |
| GPx | F: TCCTCTGTGGAAGTGGCTGA R: TCATCCAGGGGTCCGTATCT | 99 | 132 | HQ441085.1 | [25] |
| HSP70 | F: GTCCTGATCAAACGAAACACCA R: CACGCTCACCCTCATAAACCT | 101 | 128 | AY423555.2 | [26] |
| TGF-β | F: AACATCCCGCTACCTCGCTT R: TCCGCTCATCCTCATTCCCT | 94 | 116 | XM_049576571.1 | [27] |
| IgM | F: ACCGTGACCCTGACTTGCTATG R: CCCGATGGACCTGACAATAGC | 95 | 78 | GU988694.1 | [28] |
| TLR 3 | F: CTGGCTTACTACAACCACCCC | 105 | 226 | HQ857748.1 | [29] |
| R: CAAACTCCCTGCCCTCTTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, G.; Chen, X.; Feng, Y.; Yu, Z.; Lu, Q.; Li, M.; Ye, Z.; Lin, H.; Yu, W.; Shu, H. Effects of Dietary Multi-Strain Probiotics on Growth Performance, Antioxidant Status, Immune Response, and Intestinal Microbiota of Hybrid Groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂). Microorganisms 2024, 12, 1358. https://doi.org/10.3390/microorganisms12071358
Xie G, Chen X, Feng Y, Yu Z, Lu Q, Li M, Ye Z, Lin H, Yu W, Shu H. Effects of Dietary Multi-Strain Probiotics on Growth Performance, Antioxidant Status, Immune Response, and Intestinal Microbiota of Hybrid Groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂). Microorganisms. 2024; 12(7):1358. https://doi.org/10.3390/microorganisms12071358
Chicago/Turabian StyleXie, Guangting, Xiaoying Chen, Yuwei Feng, Zhide Yu, Qiuqin Lu, Manfeng Li, Ziqi Ye, Heizhao Lin, Wei Yu, and Hu Shu. 2024. "Effects of Dietary Multi-Strain Probiotics on Growth Performance, Antioxidant Status, Immune Response, and Intestinal Microbiota of Hybrid Groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂)" Microorganisms 12, no. 7: 1358. https://doi.org/10.3390/microorganisms12071358
APA StyleXie, G., Chen, X., Feng, Y., Yu, Z., Lu, Q., Li, M., Ye, Z., Lin, H., Yu, W., & Shu, H. (2024). Effects of Dietary Multi-Strain Probiotics on Growth Performance, Antioxidant Status, Immune Response, and Intestinal Microbiota of Hybrid Groupers (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂). Microorganisms, 12(7), 1358. https://doi.org/10.3390/microorganisms12071358