
High Prevalence of Antibiotic Resistance in Traditionally Fermented Foods as a Critical Risk Factor for Host Gut Antibiotic Resistome


Abstract
:1. Introduction
2. Methods and Materials
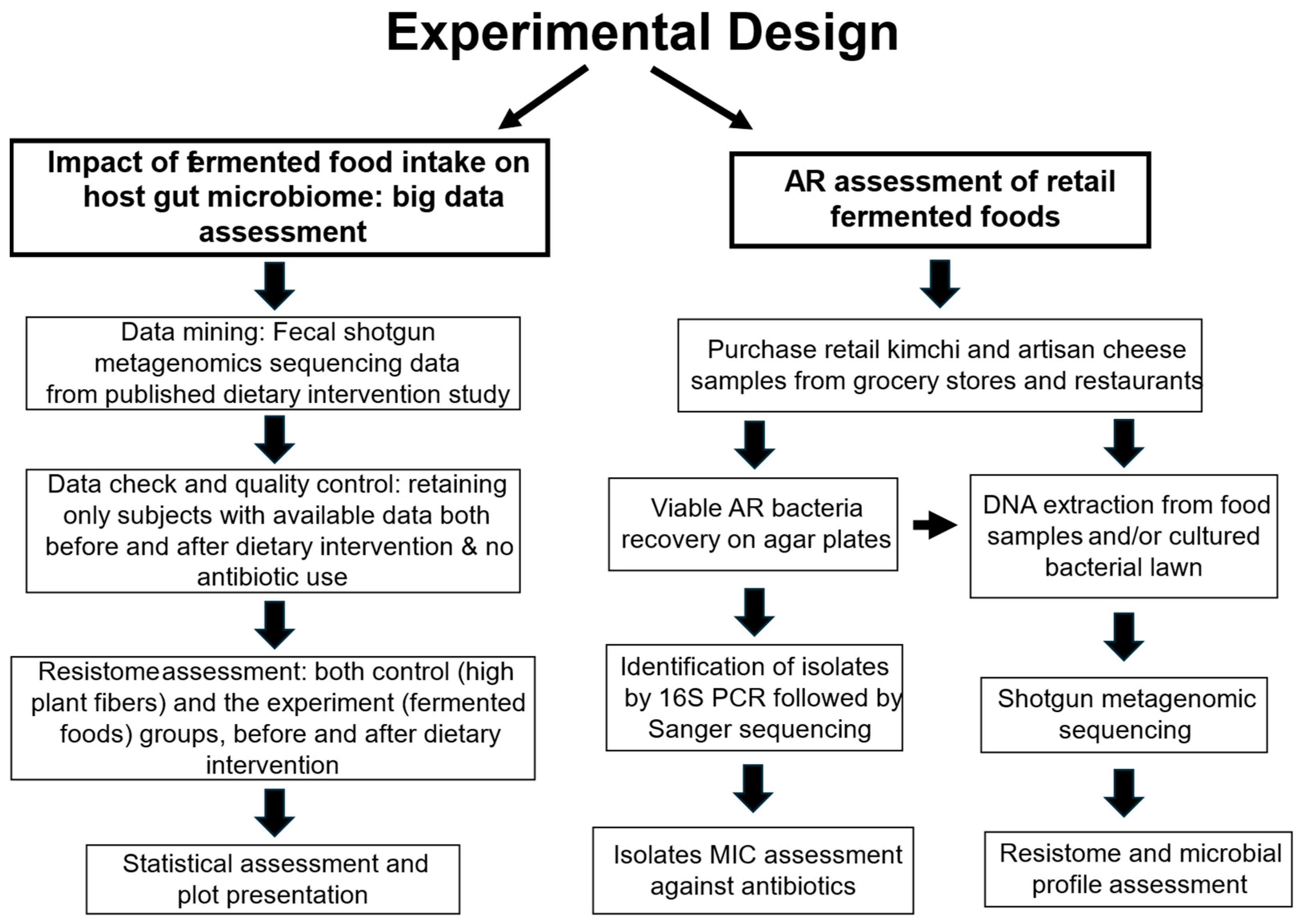
2.1. Overall Experimental Design
2.2. Host Gut Resistome Assessment
2.3. Data Transformation, Assessment, and Statistical Analysis
2.4. Source of Fermented Food Samples
2.5. Recovery and Assessment of Viable Bacteria from Fermented Food Samples
2.6. Antibiotic Resistome of Representative Fermented Food Items
3. Results
3.1. Fermented Food Intervention Led to the Rise of the Host Fecal Microbiota Resistome
3.2. Antibiotic Resistome in Traditionally Fermented Foods
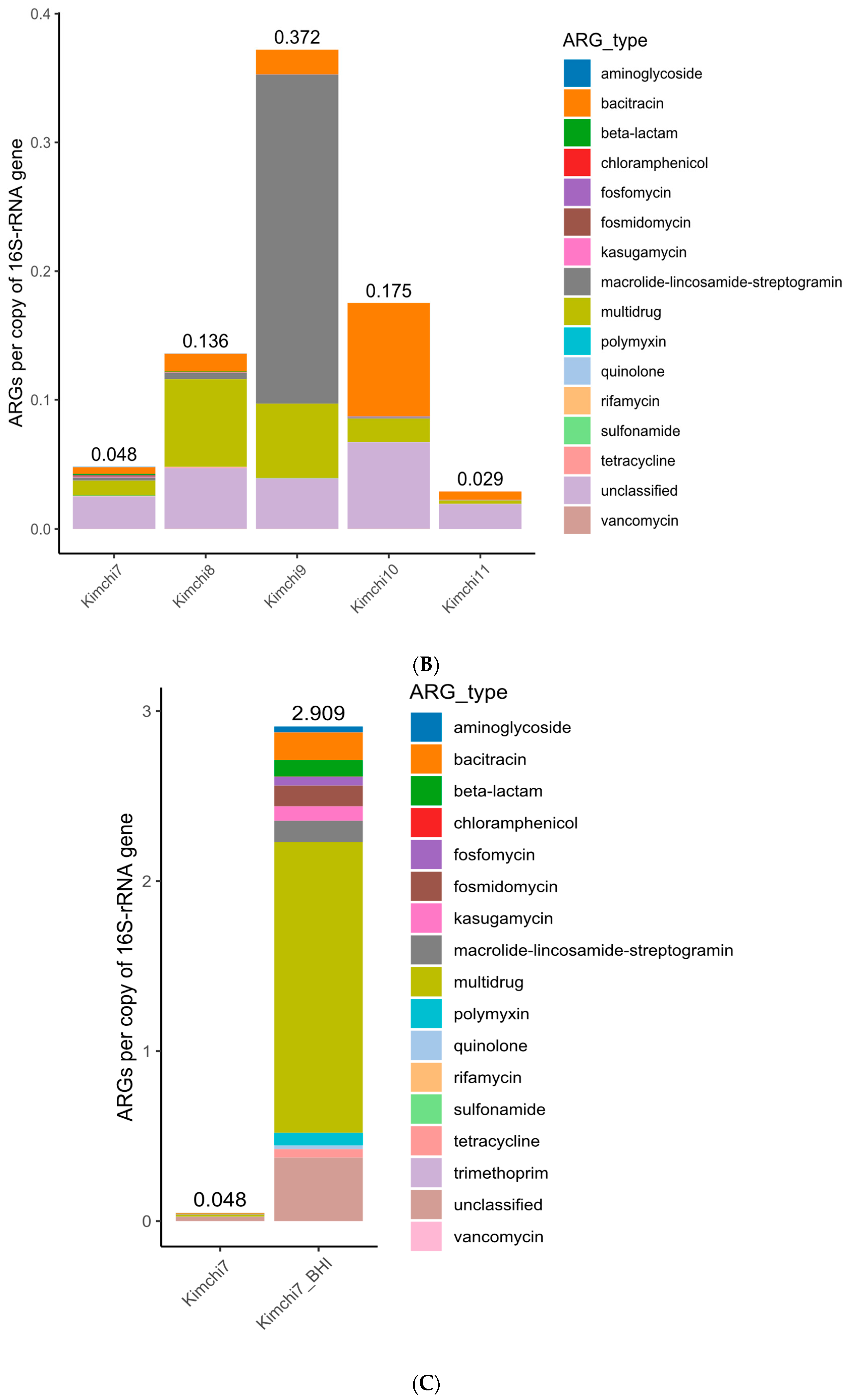
3.2.1. Antibiotic Ressitome Highly Prevalent but Diversified in Retail Kimchi Samples
3.2.2. Sample Analysis Approach Affected the Outcome Details of Kimchi Microbiome Assessments
3.2.3. Antibiotic Resistome Prevalent Even in Successfully Fermented Kimchi Samples
3.2.4. Antibiotic Resistome Highly Prevalent in Artisan Cheeses
3.2.5. Viable AR Bacteria Are Highly Prevalent in Traditionally Fermented Foods
3.2.6. Fermented Food-Originated Microbiota Can Be a Source of Many Drug-Resistant Pathogens When Exposed to Antibiotics
4. Discussion
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, Y.; Li, Y.; Zhang, L.; Wu, Z.; Huang, Y.; Yan, H.; Zhong, J.; Wang, L.-J.; Abdullah, H.M.; Wang, H.H. Antibiotic Administration Routes and Oral Exposure to Antibiotic Resistant Bacteria as Key Drivers for Gut Microbiota Disruption and Resistome in Poultry. Front. Microbiol. 2020, 11, 1319. [Google Scholar] [CrossRef]
- Blaser, M.J. Antibiotic Use and Its Consequences for the Normal Microbiome. Science 2016, 352, 544–545. [Google Scholar] [CrossRef]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut Immune Maturation Depends on Colonization with a Host-Specific Microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef]
- Foster, J.A.; McVey Neufeld, K.-A. Gut-Brain Axis: How the Microbiome Influences Anxiety and Depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. eBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human Gut Microbiota and Gastrointestinal Cancer. Genom. Proteom. Bioinforma. 2018, 16, 33–49. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile Infection. Nat. Rev. Dis. Primer 2016, 2, 16020. [Google Scholar] [CrossRef]
- Witkowski, M.; Weeks, T.L.; Hazen, S.L. Gut Microbiota and Cardiovascular Disease. Circ. Res. 2020, 127, 553–570. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Ten Threats to Global Health in 2019; World Health Organization: Geneva, Switzerland, 2019; Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (accessed on 30 December 2023).
- Wang, H.H. The Impact of Food and Drug, and the Mainstream Rescue Strategies on Host Gut Microbiota and Resistome [Conference Presentation]. In Proceedings of the 2022 Multistate AR Meeting, Ames, IA, USA, 27 October 2022. [Google Scholar]
- Zimmermann, P.; Curtis, N. The Effect of Antibiotics on the Composition of the Intestinal Microbiota—A Systematic Review. J. Infect. 2019, 79, 471–489. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, Y.; Zhang, L. Antimicrobial Resistance, Gut Microbiota, and Health. In Food Microbiology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 902–926. ISBN 978-1-68367-047-6. [Google Scholar]
- Wang, H.H. Commensal Bacteria, Microbial Ecosystems, and Horizontal Gene Transmission: Adjusting Our Focus for Strategic Breakthroughs against Antibiotic Resistance. In Food-Borne Microbes; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; pp. 267–281. ISBN 978-1-68367-135-0. [Google Scholar]
- Kelly, S.A.; Rodgers, A.M.; O’Brien, S.C.; Donnelly, R.F.; Gilmore, B.F. Gut Check Time: Antibiotic Delivery Strategies to Reduce Antimicrobial Resistance. Trends Biotechnol. 2020, 38, 447–462. [Google Scholar] [CrossRef]
- Land, M.H.; Rouster-Stevens, K.; Woods, C.R.; Cannon, M.L.; Cnota, J.; Shetty, A.K. Lactobacillus Sepsis Associated with Probiotic Therapy. Pediatrics 2005, 115, 178–181. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.; Zhou, Y.; Buckley, T.; Wang, H.H. Antibiotic Administration Routes Significantly Influence the Levels of Antibiotic Resistance in Gut Microbiota. Antimicrob. Agents Chemother. 2013, 57, 3659–3666. [Google Scholar] [CrossRef]
- Luber, A.D.; Jacobs, R.A.; Jordan, M.; Guglielmo, B.J. Relative Importance of Oral versus Intravenous Vancomycin Exposure in the Development of Vancomycin-Resistant Enterococci. J. Infect. Dis. 1996, 173, 1292–1294. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between Drugs and the Gut Microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outpatient Antibiotic Prescriptions—United States, 2020; CDC: Atlanta, GA, USA, 2020. Available online: https://www.cdc.gov/antibiotic-use/data/report-2021.html (accessed on 6 December 2022).
- Therapeutic Goods Administration (TGA). Faecal Microbiota Transplant Products Regulation; Therapeutic Goods Administration (TGA): Woden, Australia, 2021. Available online: https://www.tga.gov.au/products/biologicals-blood-and-tissues-and-advanced-therapies/biologicals/faecal-microbiota-transplant-products-regulation (accessed on 30 January 2023).
- U.S. Food and Drug Administration (FDA). FDA Approves First Fecal Microbiota Product; FDA: Silver Spring, MD, USA, 2022. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-fecal-microbiota-product (accessed on 10 July 2022).
- U.S. Food and Drug Administration (FDA). Fecal Microbiota for Transplantation: Safety Communication- Risk of Serious Adverse Reactions due to Transmission of Multi-Drug Resistant Organisms. FDA. Available online: https://www.fda.gov/safety/medical-product-safety-information/fecal-microbiota-transplantation-safety-communication-risk-serious-adverse-reactions-due (accessed on 10 July 2022).
- Liu, H.; Wang, H.H. Impact of Microbiota Transplant on Resistome of Gut Microbiota in Gnotobiotic Piglets and Human Subjects. Front. Microbiol. 2020, 11, 932. [Google Scholar] [CrossRef]
- O’Connor, A. The Best Foods to Feed Your Gut Microbiome. Washington Post, 20 September 2022. Available online: https://www.washingtonpost.com/wellness/2022/09/20/gut-health-microbiome-best-foods/ (accessed on 20 October 2022).
- Taylor, B.C.; Lejzerowicz, F.; Poirel, M.; Shaffer, J.P.; Jiang, L.; Aksenov, A.; Litwin, N.; Humphrey, G.; Martino, C.; Miller-Montgomery, S.; et al. Consumption of Fermented Foods Is Associated with Systematic Differences in the Gut Microbiome and Metabolome. mSystems 2020, 5, e00901-19. [Google Scholar] [CrossRef]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-Microbiota-Targeted Diets Modulate Human Immune Status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]
- Wang, H.H.; Manuzon, M.; Lehman, M.; Wan, K.; Luo, H.; Wittum, T.E.; Yousef, A.; Bakaletz, L.O. Food Commensal Microbes as a Potentially Important Avenue in Transmitting Antibiotic Resistance Genes. FEMS Microbiol. Lett. 2006, 254, 226–231. [Google Scholar] [CrossRef]
- Manuzon, M.Y.; Hanna, S.E.; Luo, H.; Yu, Z.; Harper, W.J.; Wang, H.H. Quantitative Assessment of the Tetracycline Resistance Gene Pool in Cheese Samples by Real-Time TaqMan PCR. Appl. Environ. Microbiol. 2007, 73, 1676–1677. [Google Scholar] [CrossRef]
- Jahan, M.; Holley, R.A. Transfer of Antibiotic Resistance from Enterococcus Faecium of Fermented Meat Origin to Listeria monocytogenes and Listeria innocua. Lett. Appl. Microbiol. 2016, 62, 304–310. [Google Scholar] [CrossRef]
- Li, X.; Alvarez, V.; Harper, W.J.; Wang, H.H. Persistent, Toxin-Antitoxin System-Independent, Tetracycline Resistance-Encoding Plasmid from a Dairy Enterococcus faecium Isolate. Appl. Environ. Microbiol. 2011, 77, 7096–7103. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Alvarez, V.; Harper, W.J.; Wang, H.H. Effective Antibiotic Resistance Mitigation during Cheese Fermentation. Appl. Environ. Microbiol. 2011, 77, 7171–7175. [Google Scholar] [CrossRef]
- Belletti, N.; Gatti, M.; Bottari, B.; Neviani, E.; Tabanelli, G.; Gardini, F. Antibiotic Resistance of Lactobacilli Isolated from Two Italian Hard Cheeses. J. Food Prot. 2009, 72, 2162–2169. [Google Scholar] [CrossRef]
- Comunian, R.; Daga, E.; Dupré, I.; Paba, A.; Devirgiliis, C.; Piccioni, V.; Perozzi, G.; Zonenschain, D.; Rebecchi, A.; Morelli, L.; et al. Susceptibility to Tetracycline and Erythromycin of Lactobacillus paracasei Strains Isolated from Traditional Italian Fermented Foods. Int. J. Food Microbiol. 2010, 138, 151–156. [Google Scholar] [CrossRef]
- Erginkaya, Z.; Turhan, E.U.; Tatlı, D. Determination of Antibiotic Resistance of Lactic Acid Bacteria Isolated from Traditional Turkish Fermented Dairy Products. Iran. J. Vet. Res. 2018, 19, 53–56. [Google Scholar]
- Guo, H.; Pan, L.; Li, L.; Lu, J.; Kwok, L.; Menghe, B.; Zhang, H.; Zhang, W. Characterization of Antibiotic Resistance Genes from Lactobacillus Isolated from Traditional Dairy Products. J. Food Sci. 2017, 82, 724–730. [Google Scholar] [CrossRef]
- Karasu, N.; Şimşek, Ö.; Çon, A.H. Technological and Probiotic Characteristics of Lactobacillus plantarum Strains Isolated from Traditionally Produced Fermented Vegetables. Ann. Microbiol. 2010, 60, 227–234. [Google Scholar] [CrossRef]
- Leech, J.; Cabrera-Rubio, R.; Walsh, A.M.; Macori, G.; Walsh, C.J.; Barton, W.; Finnegan, L.; Crispie, F.; O’Sullivan, O.; Claesson, M.J.; et al. Fermented-Food Metagenomics Reveals Substrate-Associated Differences in Taxonomy and Health-Associated and Antibiotic Resistance Determinants. mSystems 2020, 5, e00522-20. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Kromann, S.; Chen, M.; Shi, L.; Meng, H. Antibiotic Resistance of Lactobacillus Spp. and Streptococcus thermophilus Isolated from Chinese Fermented Milk Products. Foodborne Pathog. Dis. 2019, 16, 221–228. [Google Scholar] [CrossRef]
- Nawaz, M.; Wang, J.; Zhou, A.; Ma, C.; Wu, X.; Moore, J.E.; Millar, B.C.; Xu, J. Characterization and Transfer of Antibiotic Resistance in Lactic Acid Bacteria from Fermented Food Products. Curr. Microbiol. 2011, 62, 1081–1089. [Google Scholar] [CrossRef]
- Casaes Nunes, R.S.; Pires de Souza, C.; Pereira, K.S.; Del Aguila, E.M.; Flosi Paschoalin, V.M. Identification and Molecular Phylogeny of Coagulase-Negative Staphylococci Isolates from Minas Frescal Cheese in Southeastern Brazil: Superantigenic Toxin Production and Antibiotic Resistance. J. Dairy Sci. 2016, 99, 2641–2653. [Google Scholar] [CrossRef]
- Pan, L.; Hu, X.; Wang, X. Assessment of Antibiotic Resistance of Lactic Acid Bacteria in Chinese Fermented Foods. Food Control 2011, 22, 1316–1321. [Google Scholar] [CrossRef]
- Park, S.; Ji, Y.; Park, H.; Lee, K.; Park, H.; Beck, B.R.; Shin, H.; Holzapfel, W.H. Evaluation of Functional Properties of Lactobacilli Isolated from Korean White Kimchi. Food Control 2016, 69, 5–12. [Google Scholar] [CrossRef]
- Rebecchi, A.; Pisacane, V.; Callegari, M.L.; Puglisi, E.; Morelli, L. Ecology of Antibiotic Resistant Coagulase-Negative Staphylococci Isolated from the Production Chain of a Typical Italian Salami. Food Control 2015, 53, 14–22. [Google Scholar] [CrossRef]
- Touret, T.; Oliveira, M.; Semedo-Lemsaddek, T. Putative Probiotic Lactic Acid Bacteria Isolated from Sauerkraut Fermentations. PLoS ONE 2018, 13, e0203501. [Google Scholar] [CrossRef]
- Wang, J.; Li, M.; Wang, J.; Liu, M.; Yang, K.; Zhang, J.; Fan, M.; Wei, X. Antibiotic Resistance of Coagulase-Negative Staphylococci and Lactic Acid Bacteria Isolated from Naturally Fermented Chinese Cured Beef. J. Food Prot. 2018, 81, 2054–2063. [Google Scholar] [CrossRef]
- Yasir, M.; Al-Zahrani, I.A.; Bibi, F.; Abd El Ghany, M.; Azhar, E.I. New Insights of Bacterial Communities in Fermented Vegetables from Shotgun Metagenomics and Identification of Antibiotic Resistance Genes and Probiotic Bacteria. Food Res. Int. 2022, 157, 111190. [Google Scholar] [CrossRef]
- Zhou, N.; Zhang, J.X.; Fan, M.T.; Wang, J.; Guo, G.; Wei, X.Y. Antibiotic Resistance of Lactic Acid Bacteria Isolated from Chinese Yogurts. J. Dairy Sci. 2012, 95, 4775–4783. [Google Scholar] [CrossRef]
- Zielińska, D.; Rzepkowska, A.; Radawska, A.; Zieliński, K. In Vitro Screening of Selected Probiotic Properties of Lactobacillus Strains Isolated from Traditional Fermented Cabbage and Cucumber. Curr. Microbiol. 2015, 70, 183–194. [Google Scholar] [CrossRef]
- Wang, H.H.; Schaffner, D.W. Antibiotic Resistance: How Much Do We Know and Where Do We Go from Here? Appl. Environ. Microbiol. 2011, 77, 7093–7095. [Google Scholar] [CrossRef]
- Future Market Insights. Fermented Foods and Beverages Market; Future Market Insights: Newark, DE, USA, 2022; Available online: https://www.futuremarketinsights.com/reports/fermented-foods-and-beverages-market (accessed on 30 October 2022).
- Manskar, N.; Raskin, S. Here’s Why Sauerkraut and Kimchi Sales Have Surged during the Coronavirus. New York Post, 17 April 2020. Available online: https://nypost.com/2020/04/17/heres-why-sauerkraut-and-kimchi-sales-are-surging-thanks-to-coronavirus/ (accessed on 23 October 2022).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinforma. Oxf. Engl. 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Yin, X.; Jiang, X.-T.; Chai, B.; Li, L.; Yang, Y.; Cole, J.R.; Tiedje, J.M.; Zhang, T. ARGs-OAP v2.0 with an Expanded SARG Database and Hidden Markov Models for Enhancement Characterization and Quantification of Antibiotic Resistance Genes in Environmental Metagenomes. Bioinformatics 2018, 34, 2263–2270. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Southend Oaks, CA, USA, 2018. [Google Scholar]
- Patil, I. Visualizations with Statistical Details: The “ggstatsplot” Approach. J. Open Source Softw. 2021, 6, 3167. [Google Scholar] [CrossRef]
- Wickham, H. Data Analysis. In ggplot2: Elegant Graphics for Data Analysis; Wickham, H., Ed.; Use R! Springer International Publishing: Cham, Switzerland, 2016; pp. 189–201. ISBN 978-3-319-24277-4. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation. R Package Version 1.0.10; 2022. Available online: https://github.com/tidyverse/dplyr (accessed on 31 December 2022).
- Stock, I.; Grueger, T.; Wiedemann, B. Natural Antibiotic Susceptibility of Strains of Serratia marcescens and the S. liquefaciens Complex: S. liquefaciens Sensu Stricto, S. proteamaculans and S. grimesii. Int. J. Antimicrob. Agents 2003, 22, 35–47. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Antonoplis, A.; Zang, X.; Wegner, T.; Wender, P.A.; Cegelski, L. Vancomycin-Arginine Conjugate Inhibits Growth of Carbapenem-Resistant E. coli and Targets Cell-Wall Synthesis. ACS Chem. Biol. 2019, 14, 2065–2070. [Google Scholar] [CrossRef]
- Soares, G.M.S.; Figueiredo, L.C.; Faveri, M.; Cortelli, S.C.; Duarte, P.M.; Feres, M. Mechanisms of Action of Systemic Antibiotics Used in Periodontal Treatment and Mechanisms of Bacterial Resistance to These Drugs. J. Appl. Oral Sci. Rev. FOB 2012, 20, 295–309. [Google Scholar] [CrossRef]
- Farmer, J.J., III. Enterobacteriaceae: Introduction and Identification. In Manual of Clinical Microbiology; American Society for Microbiology: Washington, DC, USA, 2003; Volume 1, pp. 636–653. [Google Scholar]
- Mahlen, S.D. Serratia Infections: From Military Experiments to Current Practice. Clin. Microbiol. Rev. 2011, 24, 755–791. [Google Scholar] [CrossRef]
- Casado Muñoz, M.d.C.; Benomar, N.; Lerma, L.L.; Gálvez, A.; Abriouel, H. Antibiotic Resistance of Lactobacillus pentosus and Leuconostoc pseudomesenteroides Isolated from Naturally-Fermented Aloreña Table Olives throughout Fermentation Process. Int. J. Food Microbiol. 2014, 172, 110–118. [Google Scholar] [CrossRef]
- Fraqueza, M.J. Antibiotic Resistance of Lactic Acid Bacteria Isolated from Dry-Fermented Sausages. Int. J. Food Microbiol. 2015, 212, 76–88. [Google Scholar] [CrossRef]
- Kim, T.; Heo, S.; Na, H.-E.; Lee, G.; Kim, J.-H.; Kwak, M.-S.; Sung, M.-H.; Jeong, D.-W. Bacterial Community of Galchi-Baechu Kimchi Based on Culture-Dependent and—Independent Investigation and Selection of Starter Candidates. J. Microbiol. Biotechnol. 2022, 32, 341–347. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Magez, S.; Peeters, E.; Chung, M.S.; Kim, K.H.; Radwanska, M. Comprehensive Genomic Analysis Reveals Virulence Factors and Antibiotic Resistance Genes in Pantoea agglomerans KM1, a Potential Opportunistic Pathogen. PLoS ONE 2021, 16, e0239792. [Google Scholar] [CrossRef]
- Rengaraj, R.; Mariappan, S.; Sekar, U.; Kamalanadhan, A. Detection of Vancomycin Resistance among Enterococcus faecalis and Staphylococcus aureus. J. Clin. Diagn. Res. JCDR 2016, 10, DC04-06. [Google Scholar] [CrossRef]
- Gao, W.; Zhu, Y.; Ye, J.; Chu, H. Gut Non-Bacterial Microbiota Contributing to Alcohol-Associated Liver Disease. Gut Microbes 2021, 13, 1984122. [Google Scholar] [CrossRef]
- Angurana, S.K.; Bansal, A.; Singhi, S.; Aggarwal, R.; Jayashree, M.; Salaria, M.; Mangat, N.K. Evaluation of Effect of Probiotics on Cytokine Levels in Critically Ill Children With Severe Sepsis: A Double-Blind, Placebo-Controlled Trial. Crit. Care Med. 2018, 46, 1656–1664. [Google Scholar] [CrossRef]
- Aydoğan, S.; Dilli, D.; Özyazici, A.; Aydin, N.; Şimşek, H.; Orun, U.A.; Aksoy, Ö.N. Lactobacillus rhamnosus Sepsis Associated with Probiotic Therapy in a Term Infant with Congenital Heart Disease. Fetal Pediatr. Pathol. 2022, 41, 823–827. [Google Scholar] [CrossRef]
- Bertelli, C.; Pillonel, T.; Torregrossa, A.; Prod’hom, G.; Fischer, C.J.; Greub, G.; Giannoni, E. Bifidobacterium longum Bacteremia in Preterm Infants Receiving Probiotics. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 924–927. [Google Scholar] [CrossRef]
- Boyle, R.J.; Robins-Browne, R.M.; Tang, M.L.K. Probiotic Use in Clinical Practice: What Are the Risks? Am. J. Clin. Nutr. 2006, 83, 1256–1264; quiz 1446–1447. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.-C.; Chen, C.-L.; Feng, Y.; Chen, C.-C.; Lien, R.; Chiu, C.-H. Lactobacillus rhamnosus Sepsis Associated with Probiotic Therapy in an Extremely Preterm Infant: Pathogenesis and a Review for Clinicians. J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 2021, 54, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Colautti, A.; Arnoldi, M.; Comi, G.; Iacumin, L. Antibiotic Resistance and Virulence Factors in Lactobacilli: Something to Carefully Consider. Food Microbiol. 2022, 103, 103934. [Google Scholar] [CrossRef] [PubMed]
- Kochan, P.; Chmielarczyk, A.; Szymaniak, L.; Brykczynski, M.; Galant, K.; Zych, A.; Pakosz, K.; Giedrys-Kalemba, S.; Lenouvel, E.; Heczko, P.B. Lactobacillus Rhamnosus Administration Causes Sepsis in a Cardiosurgical Patient--Is the Time Right to Revise Probiotic Safety Guidelines? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2011, 17, 1589–1592. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, T.; Majarikar, S.; Deshmukh, M.; Ananthan, A.; Balasubramanian, H.; Keil, A.; Patole, S. Probiotic Sepsis in Preterm Neonates-a Systematic Review. Eur. J. Pediatr. 2022, 181, 2249–2262. [Google Scholar] [CrossRef]
- Ohishi, A.; Takahashi, S.; Ito, Y.; Ohishi, Y.; Tsukamoto, K.; Nanba, Y.; Ito, N.; Kakiuchi, S.; Saitoh, A.; Morotomi, M.; et al. Bifidobacterium Septicemia Associated with Postoperative Probiotic Therapy in a Neonate with Omphalocele. J. Pediatr. 2010, 156, 679–681. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample # | Rank | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| K1–K4 (BHI, pooled) | Multidrug 0.3189 * | Bacitracin 0.0901 | Unclassified 0.0780 | MLS ** 0.0684 | Fosfomycin 0.0465 |
| K1–K4 (BHI-Tet, pooled) | Multidrug 0.2556 | Tetracycline 0.1059 | Unclassified 0.0777 | Bacitracin 0.0485 | MLS 0.0422 |
| K1–K4 (BHI-Amp, pooled) | Multidrug 0.3235 | Bacitracin 0.1795 | β-lactam 0.0680 | MLS 0.0557 | Fosfomycin 0.0498 |
| K7 (BHI) | Multidrug 1.7085 | Unclassified 0.3728 | Bacitracin 0.1609 | MLS 0.1278 | Fosmidomycin 0.1216 |
| K7 | Unclassified 0.0248 | Multidrug 0.0118 | Bacitracin 0.0050 | MLS 0.0023 | Fosmidomycin 0.0012 |
| K8 | Multidrug 0.0681 | Unclassified 0.0469 | Bacitracin 0.0136 | MLS 0.0047 | Tetracycline 0.0006 |
| K9 | MLS 0.2557 | Multidrug 0.0577 | Unclassified 0.0390 | Bacitracin 0.0191 | Vancomycin 0.0001 |
| K10 | Bacitracin 0.0880 | Unclassified 0.0669 | Multidrug 0.0182 | MLS 0.0009 | Fosfomycin 0.0005 |
| K11 | Unclassified 0.0191 | Bacitracin 0.0063 | Multidrug 0.0026 | Tetracycline 0.0002 | MLS * 0.0002 |
| Isolate | Identity | Food Source | Recovered From | Resistance with MIC * (µg/mL) |
|---|---|---|---|---|
| K5-1 *** | Latilactobacillus sakei | Kimchi 5 | MRS-tet | Tet (>64) Van (>128) |
| K5-2 | Bacillus (licheniformis/paralicheniformis) | Kimchi 5 | BHI-ery | Amp (>64) Ery (>16) |
| K7-5 | Serratia quinivorans | Kimchi 7 | BHI/BHI-amp | Amp (>128) Van (>1024) ** |
| K7-6 *** | Klebsiella pneumoniae | Kimchi 7 | BHI/BHI-amp | Amp (>128) Ery (>100) ** Van (>1024) ** |
| K7-8 | Enterobacter sp. | Kimchi 7 | BHI/BHI-amp | Amp (>128) Ery (>100) ** Van (>1024) ** |
| K7-9 | Rahnella aquatilis | Kimchi 7 | BHI/BHI-amp | Amp (>128) Ery (>100) ** Van(>1024) ** |
| K8-6 | Bacillus licheniformis | Kimchi 8 | BHI-ery | Amp (>32) Ery (>16) |
| K9-1 *** | Latilactobacillus curvatus | Kimchi 9 | MRS-tet | Tet (>64) Van (>128) |
| K9-2 | Bacillus (licheniformis/paralicheniformis) | Kimchi 9 | BHI-ery | Amp (>16) Ery (>16) |
| K9-3 *** | Latilactobacillus sakei | Kimchi 9 | MRS-ery | Ery (>16) Van (>128) |
| K10-1 *** | Levilactobacillus brevis | Kimchi 10 | BHI-tet | Tet (>8) Van (>128) |
| K10-2 | Levilactobacillus brevis | Kimchi 10 | BHI-tet | Tet (>8) Van (>128) |
| K11-3 *** | Rahnella aquatilis | Kimchi 11 | MRS-amp | Amp (>128) Van (>512) ** |
| K12-1 *** | Leuconostoc citreum | Kimchi 12 | BHI-tet | Tet (>64) Van (>128) |
| K12-6 | Latilactobacillus sakei | Kimchi 12 | BHI-tet | Tet (>32) Van (>128) |
| K13-1 *** | Serratia marcescens | Kimchi 13 | BHI-tet | Amp (>128) Ery (>200) ** Tet (>128) Van (>1024) ** |
| K13-2 | Oceanobacillis jeddahense | Kimchi 13 | BHI-tet | Tet (>32) Ery (>16) |
| K14-2 *** | Levilactobacillus brevis | Kimchi 14 | MRS/MRS-tet | Tet (>16) |
| Sample | Top Genera | Percentage |
|---|---|---|
| Kimchi7 | Leuconostoc | 94.87% |
| Other | 3.17% | |
| Rahnella | 0.85% | |
| Weissella | 0.48% | |
| Levilactobacillus | 0.36% | |
| Xanthomonas | 0.27% | |
| Kimchi8 | Latilactobacillus | 55.98% |
| Leuconostoc | 28.90% | |
| Lactiplantibacillus | 7.49% | |
| Other | 5.46% | |
| Lactococcus | 1.59% | |
| Levilactobacillus | 0.58% | |
| Kimchi9 | Latilactobacillus | 81.60% |
| Leuconostoc | 11.79% | |
| Other | 3.45% | |
| Weissella | 2.02% | |
| Lactiplantibacillus | 0.59% | |
| Levilactobacillus | 0.55% | |
| Kimchi10 | Lactiplantibacillus | 54.60% |
| Leuconostoc | 32.89% | |
| Levilactobacillus | 9.80% | |
| Other | 1.86% | |
| Weissella | 0.55% | |
| Latilactobacillus | 0.30% | |
| Kimchi11 | Weissella | 58.85% |
| Leuconostoc | 39.19% | |
| Other | 1.41% | |
| Latilactobacillus | 0.23% | |
| Lactiplantibacillus | 0.19% | |
| Stenotrophomonas | 0.13% |
| Sample # | Rank | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| C1–C4 (BHI, pooled) | Multidrug 0.4704 * | MLS ** 0.0783 | Bacitracin 0.0353 | Fosfomycin 0.0265 | Tetracycline 0.0105 |
| C1–C4 (BHI-Tet, pooled) | Tetracycline 0.5377 | Multidrug 0.2359 | Unclassified 0.0876 | Bacitracin 0.0744 | MLS 0.0295 |
| C1–C4 (BHI-Amp, pooled) | Multidrug 0.0308 | Tetracycline 0.0093 | MLS 0.0084 | Bacitracin 0.0053 | Unclassified 0.0033 |
| C5 (BHI) | Multidrug 0.1343 | Fosfomycin 0.1123 | Unclassified 0.0709 | β-lactam 0.0617 | MLS 0.0555 |
| C5 (BHI-Tet) | Multidrug 0.0033 | β-lactam 0.0031 | Tetracycline 0.0014 | Bacitracin 0.0012 | Unclassified 0.0012 |
| C5 (BHI-Amp) | Multidrug 0.0040 | β-lactam 0.0038 | Unclassified 0.0015 | Bacitracin 0.0014 | MLS 0.0013 |
| C6 (BHI) | Multidrug 0.6202 | Tetracycline 0.4674 | Bacitracin 0.2046 | Unclassified 0.1750 | MLS 0.0694 |
| C7 (BHI) | Multidrug 0.3538 | Tetracycline 0.2740 | Unclassified 0.1442 | Bacitracin 0.1308 | MLS 0.0557 |
| C7 (BHI-Tet) | Multidrug 0.2558 | Tetracycline 0.2215 | Unclassified 0.1183 | Bacitracin 0.1176 | Quinolone 0.0167 |
| C7 (BHI-Amp) | Multidrug 0.0394 | Bacitracin 0.0085 | Tetracycline 0.0078 | Unclassified 0.0075 | β-lactam 0.0035 |
| C8 (BHI) | Multidrug 0.2605 | Unclassified 0.0737 | Vancomycin 0.0671 | Bacitracin 0.0472 | MLS 0.0345 |
| Isolate | Identity | Food Source | Recovered From | Resistance with MIC * (µg/mL) |
|---|---|---|---|---|
| C5-3 | Staphylococcus equorum | Cheese 5 | BHI-ery | Ery (>16) |
| C5-9 | Mammaliicoccus vitulinus | Cheese 5 | BHI-ery | Ery (>8) |
| C6-2 ** | Staphylococcus xylosus | Cheese 6 | BHI-tet | Tet (>16) |
| C6-4 | Staphylococcus saprophyticus | Cheese 6 | BHI-tet | Tet (>32) |
| C6-10 | Staphylococcus xylosus | Cheese 6 | BHI-tet | Tet (>32) Van (>128) |
| C6-11 ** | Enterococcus faecalis | Cheese 6 | MRS-ery | Ery (>4) Van (>128) |
| C7-1 | Staphylococcus xylosus | Cheese 7 | BHI-tet | Tet (>8) |
| C7-5 | Staphylococcus saprophyticus | Cheese 7 | BHI-tet | Tet (>32) |
| C8-11 | Staphylococcus equorum | Cheese 8 | BHI-ery | Ery (>16) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Fu, S.; Klein, M.S.; Wang, H. High Prevalence of Antibiotic Resistance in Traditionally Fermented Foods as a Critical Risk Factor for Host Gut Antibiotic Resistome. Microorganisms 2024, 12, 1433. https://doi.org/10.3390/microorganisms12071433
Li Y, Fu S, Klein MS, Wang H. High Prevalence of Antibiotic Resistance in Traditionally Fermented Foods as a Critical Risk Factor for Host Gut Antibiotic Resistome. Microorganisms. 2024; 12(7):1433. https://doi.org/10.3390/microorganisms12071433
Chicago/Turabian StyleLi, Yutong, Siying Fu, Matthias S. Klein, and Hua Wang. 2024. "High Prevalence of Antibiotic Resistance in Traditionally Fermented Foods as a Critical Risk Factor for Host Gut Antibiotic Resistome" Microorganisms 12, no. 7: 1433. https://doi.org/10.3390/microorganisms12071433