
Implementation of Spore Display in Paenibacillus polymyxa with Different Hydrolytic Enzymes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. Strain and Plasmid Construction
2.3. Sporulation and Purification
2.4. Fluorescence Microscopy and Image Analysis
2.5. Cytometric Analysis
2.6. Hydrolysis Activity Assay
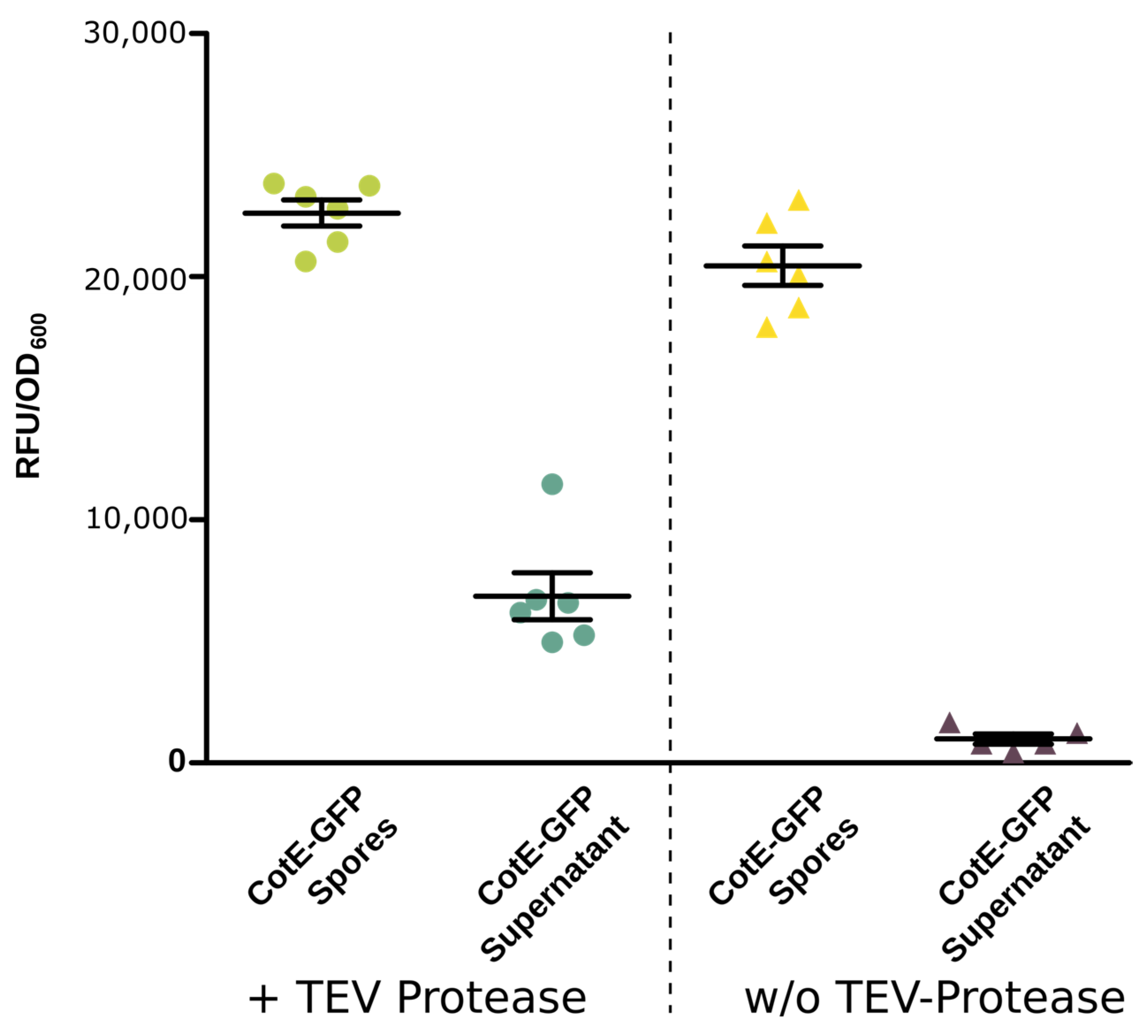
2.7. Tobacco Etch Virus (TEV) Cleavage Assay
2.8. Recycling of Spore Activity
2.9. Statistical Analysis
3. Results
3.1. Construction of Strains and Plasmids for the Expression of Fusion Proteins on the Spore Surface
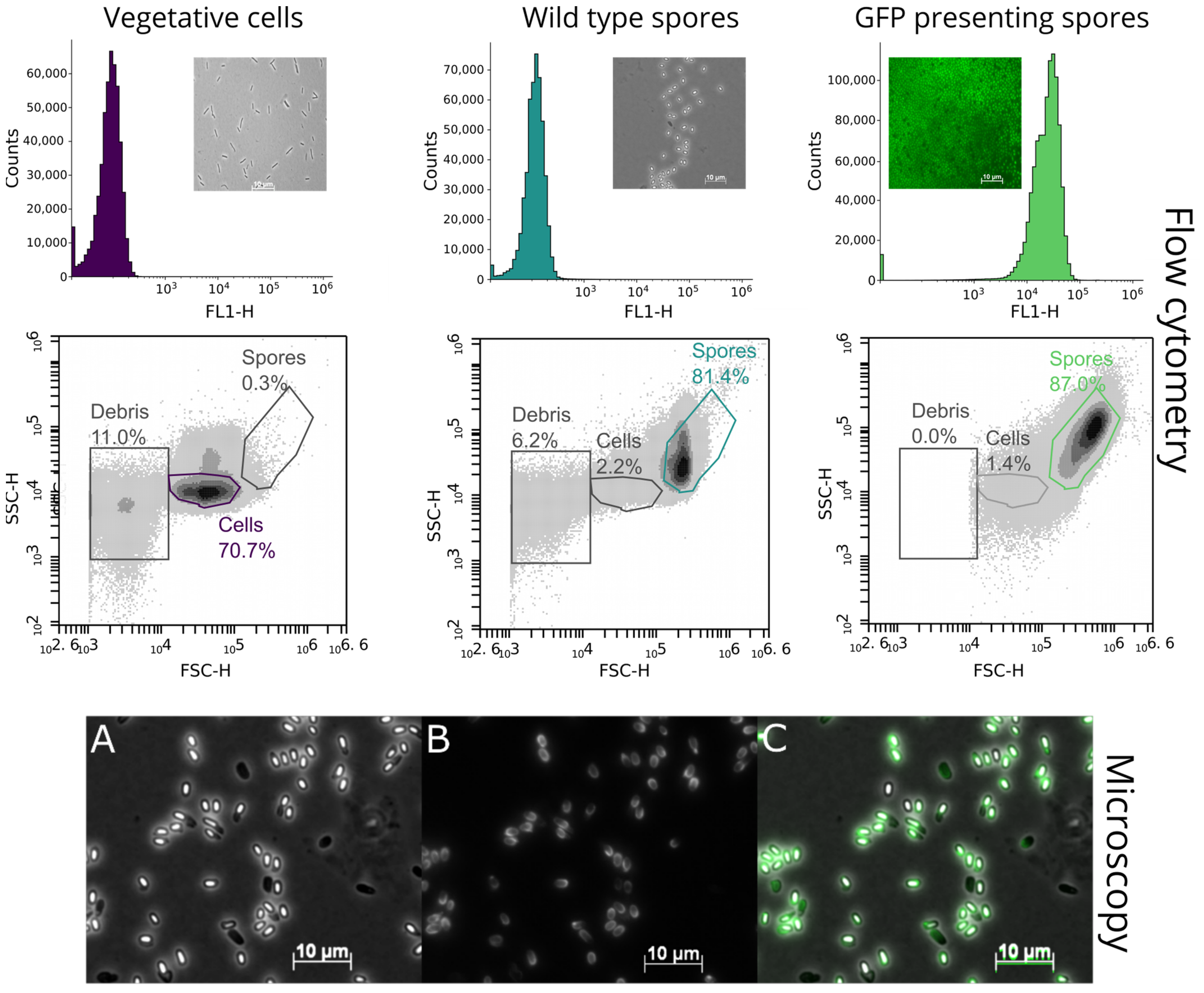
3.2. Identification of the Green Fluorescent Protein Display via Fluorescence Microscopy and Flow Cytometry
3.3. Enzyme Activity Assays of the CotE-Lip3, CotE-LipA, and CotE-PnbA Fusion Proteins
3.4. Recyclability and Reusability of the Spore Display
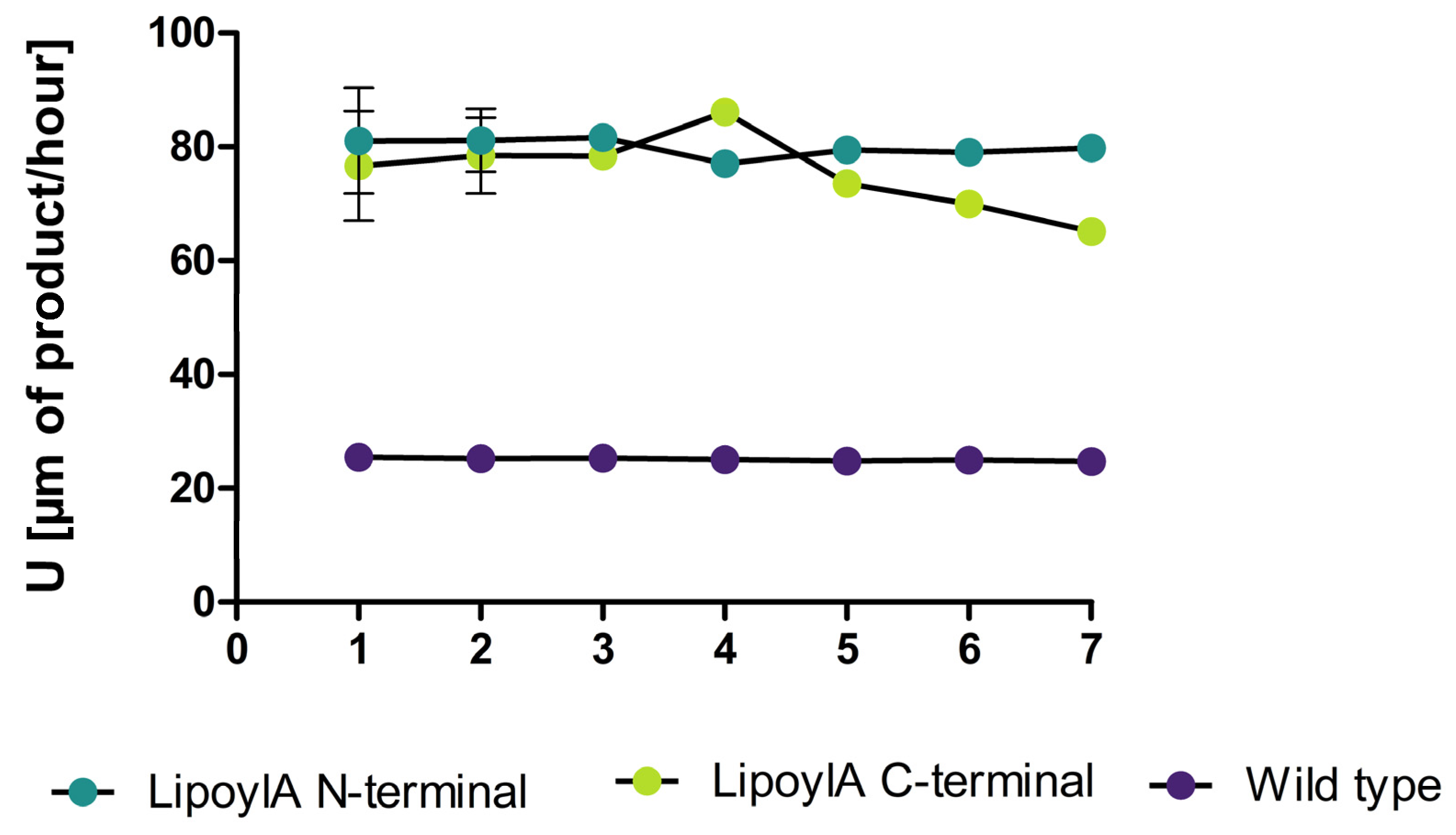
3.5. Hydrolytic Side Activity of the Lipoyl Synthase a Displayed on P. polymyxa Spores
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papadopoulou, C.-I.; Loizou, E.; Chatzitheodoridis, F. Priorities in Bioeconomy Strategies: A Systematic Literature Review. Energies 2022, 15, 7258. [Google Scholar] [CrossRef]
- Ramos, M.D.N.; Milessi, T.S.; Candido, R.G.; Mendes, A.A.; Aguiar, A. Enzymatic catalysis as a tool in biofuels production in Brazil: Current status and perspectives. Energy Sustain. Dev. 2022, 68, 103–119. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, Z.; Xu, J.; Wu, Q. Recent Advances in Photobiocatalysis for Selective Organic Synthesis. Org. Process Res. Dev. 2022, 26, 1900–1913. [Google Scholar] [CrossRef]
- Nunes, Y.L.; de Menezes, F.L.; de Sousa, I.G.; Cavalcante, A.L.G.; Cavalcante, F.T.T.; da Silva Moreira, K.; de Oliveira, A.L.B.; Mota, G.F.; da Silva Souza, J.E.; de Aguiar Falcao, I.R.; et al. Chemical and physical Chitosan modification for designing enzymatic industrial biocatalysts: How to choose the best strategy? Int. J. Biol. Macromol. 2021, 181, 1124–1170. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Asgher, M.; Cheng, H.; Yan, Y.; Iqbal, H.M.N. Multi-point enzyme immobilization, surface chemistry, and novel platforms: A paradigm shift in biocatalyst design. Crit. Rev. Biotechnol. 2019, 39, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Remonatto, D.; Miotti, R.H., Jr.; Monti, R.; Bassan, J.C.; de Paula, A.V. Applications of immobilized lipases in enzymatic reactors: A review. Process Biochem. 2022, 114, 1–20. [Google Scholar] [CrossRef]
- ASheldon, R.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: Robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Dorr, B.M.; Liu, D.R. A general strategy for the evolution of bond-forming enzymes using yeast display. Proc. Natl. Acad. Sci. USA 2011, 108, 11399–11404. [Google Scholar] [CrossRef] [PubMed]
- Hosse, R.J.; Rothe, A.; Power, B.E. A new generation of protein display scaffolds for molecular recognition. Protein Sci. 2006, 15, 14–27. [Google Scholar] [CrossRef]
- Bamba, T.; Guirimand, G.; Kondo, A.; Hasunuma, T. Enzyme display technology for lignocellulosic biomass valorization by yeast cell factories. Curr. Opin. Green. Sustain. Chem. 2022, 33, 100584. [Google Scholar] [CrossRef]
- Ghaemmaghami, S.; Huh, W.-K.; Bower, K.; Howson, R.W.; Belle, A.; Dephoure, N.; O’Shea, E.K.; Weissman, J.S. Global analysis of protein expression in yeast. Nature 2003, 425, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gupta, G.; Ahmad, T.; Mansoor, S.; Kaur, B. Enzyme Engineering: Current Trends and Future Perspectives. Food Rev. Int. 2021, 37, 121–154. [Google Scholar] [CrossRef]
- Chowdhury, R.; Maranas, C.D. From directed evolution to computational enzyme engineering—A review. AIChE J. 2020, 66, e16847. [Google Scholar] [CrossRef]
- Guirimand, G.G.Y.; Bamba, T.; Matsuda, M.; Inokuma, K.; Morita, K.; Kitada, Y.; Kobayashi, Y.; Yukawa, T.; Sasaki, K.; Ogino, C.; et al. Combined cell surface display of β-D-glucosidase (BGL), maltose transporter (MAL11) and overexpression of cytosolic xylose reductase (XR) in Saccharomyces cerevisiae enhance cellobiose/xylose co-utilization for xylitol bio-production from lignocellulosic biomass. Biotechnol. J. 2019, 14, 1800704. [Google Scholar]
- Aharoni, A.; Griffiths, A.D.; Tawfik, D.S. High-throughput screens and selections of enzyme-encoding genes. Curr. Opin. Chem. Biol. 2005, 9, 210–216. [Google Scholar] [CrossRef]
- Mandeep Gupta, G.K.; Shukla, P. Enzyme Engineering Techniques for Biotechnological Applications. In Microbial Enzymes and Biotechniques: Interdisciplinary Perspectives; Shukla, P., Ed.; Springer: Singapore, 2020; pp. 235–249. [Google Scholar]
- Ali, M.; Ishqi, H.M.; Husain, Q. Enzyme engineering: Reshaping the biocatalytic functions. Biotechnol. Bioeng. 2020, 117, 1877–1894. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Holmes, M.; Schaefer, E.; Mulchandani, A.; Ge, X. Highly active spore biocatalyst by self-assembly of co-expressed anchoring scaffoldin and multimeric enzyme. Biotechnol. Bioeng. 2018, 115, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Jose, J. Autodisplay: Efficient bacterial surface display of recombinant proteins. Appl. Microbiol. Biotechnol. 2006, 69, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Balderas Hernández, V.E.; Salas-Montantes, C.J.; Barba-De la Rosa, A.P.; De Leon-Rodriguez, A. Autodisplay of an endo-1,4-β-xylanase from Clostridium cellulovorans in Escherichia coli for xylans degradation. Enzyme Microb. Technol. 2021, 149, 109834. [Google Scholar] [CrossRef]
- Gutiérrez-García, A.K.; Alvarez-Guzmán, C.L.; De Leon-Rodriguez, A. Autodisplay of alpha amylase from Bacillus megaterium in E. coli for the bioconversion of starch into hydrogen, ethanol and succinic acid. Enzyme Microb. Technol. 2020, 134, 109477. [Google Scholar] [CrossRef]
- Strätker, K.; Haidar, S.; Dubiel, M.; Estévez-Braun, A.; Jose, J. Autodisplay of human PIP5K1α lipid kinase on Escherichia coli and inhibitor testing. Enzyme Microb. Technol. 2021, 143, 109717. [Google Scholar] [CrossRef]
- Huo, Z.; Zhang, N.; Raza, W.; Huang, X.; Yong, X.; Liu, Y.; Wang, D.; Li, S.; Shen, Q.; Zhang, R. Comparison of the spores of Paenibacillus polymyxa prepared at different temperatures. Biotechnol. Lett. 2012, 34, 925–933. [Google Scholar] [CrossRef]
- Karava, M.; Gockel, P.; Kabisch, J. Bacillus subtilis spore surface display of photodecarboxylase for the transformation of lipids to hydrocarbons. Sustain. Energy Fuels 2021, 5, 1727–1733. [Google Scholar] [CrossRef]
- Chen, H.; Ullah, J.; Jia, J. Progress in Bacillus subtilis Spore Surface Display Technology towards Environment, Vaccine Development, and Biocatalysis. J. Mol. Microbiol. Biotechnol. 2017, 27, 159–167. [Google Scholar] [CrossRef]
- Petrillo, C.; Castaldi, S.; Lanzilli, M.; Saggese, A.; Donadio, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The temperature of growth and sporulation modulates the efficiency of spore-display in Bacillus subtilis. Microb. Cell Factories 2020, 19, 185. [Google Scholar] [CrossRef]
- Kim, J. Surface display of lipolytic enzyme, Lipase A and Lipase B of Bacillus subtilis on the Bacillus subtilis spore. Biotechnol. Bioprocess. Eng. 2017, 22, 462–468. [Google Scholar] [CrossRef]
- Hui, Y.; Cui, Z.; Sim, S. Stress-Tolerant, Recyclable, and Renewable Biocatalyst Platform Enabled by Engineered Bacterial Spores. ACS Synth. Biol. 2022, 11, 2857–2868. [Google Scholar] [CrossRef]
- Knecht, L.D.; Pasini, P.; Daunert, S. Bacterial spores as platforms for bioanalytical and biomedical applications. Anal. Bioanal. Chem. 2011, 400, 977–989. [Google Scholar] [CrossRef]
- Isticato, R.; Ricca, E. Spore Surface Display. In The Bacterial Spore; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 349–366. [Google Scholar]
- Csibra, E.; Renders, M.; Pinheiro, V.B. Bacterial Cell Display as a Robust and Versatile Platform for Engineering Low-Affinity Ligands and Enzymes. ChemBioChem 2020, 21, 2844–2853. [Google Scholar] [CrossRef]
- Lin, P.; Yuan, H.; Du, J.; Liu, K.; Liu, H.; Wang, T. Progress in research and application development of surface display technology using Bacillus subtilis spores. Appl. Microbiol. Biotechnol. 2020, 104, 2319–2331. [Google Scholar] [CrossRef]
- Zhang, G.; An, Y.; Zabed, H.M.; Guo, Q.; Yang, M.; Yuan, J.; Li, W.; Sun, W.; Qi, X. Bacillus subtilis Spore Surface Display Technology: A Review of Its Development and Applications. J. Microbiol. Biotechnol. 2019, 29, 179–190. [Google Scholar]
- Curtis, D.; Golomb, B.L.; Ally, D.; Ficarra, F.A.; Salamzade, R.; Traag, B.A. Paenibacillus-based endospore display platform, products and methods. U.S. Patent 10,988,769, 27 April 2021. [Google Scholar]
- Langendries, S.; Goormachtig, S. Paenibacillus polymyxa, a Jack of all trades. Environ. Microbiol. 2021, 23, 5659–5669. [Google Scholar] [CrossRef]
- Hinc, K.; Iwanicki, A.; Obuchowski, M. New stable anchor protein and peptide linker suitable for successful spore surface display in B. subtilis. Microb. Cell Factories 2013, 12, 22. [Google Scholar] [CrossRef]
- Chen, X.; Zaro, J.L.; Shen, W.-C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef]
- van Rosmalen, M.; Krom, M.; Merkx, M. Tuning the Flexibility of Glycine-Serine Linkers To Allow Rational Design of Multidomain Proteins. Biochemistry 2017, 565, 6565–6574. [Google Scholar] [CrossRef]
- Hosseini-Abari, A.; Kim, B.-G.; Lee, S.-H.; Emtiazi, G.; Kim, W.; Kim, J.-H. Surface display of bacterial tyrosinase on spores of Bacillus subtilis using CotE as an anchor protein. J. Basic. Microbiol. 2016, 56, 1331–1337. [Google Scholar] [CrossRef]
- Kang, S.J.; Park, E.A.; Lee, D.H.; Hong, K.W. Comparison of the stability of eGFP displayed on the Bacillus subtilis spore surface using CotB and C-terminally truncated CotB proteins as an anchoring motif under extreme conditions. Appl. Biol. Chem. 2019, 62, 41. [Google Scholar] [CrossRef]
- Akram, F.; Mir, A.S.; Haq, I.U.; Roohi, A. An Appraisal on Prominent Industrial and Biotechnological Applications of Bacterial Lipases. Mol. Biotechnol. 2023, 65, 521–543. [Google Scholar] [CrossRef]
- Kazlauskas, R.J.; Bornscheuer, U.T. Biotransformations with Lipases. In Biotechnology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1998; pp. 36–191. [Google Scholar]
- Hama, S.; Noda, H.; Kondo, A. How lipase technology contributes to evolution of biodiesel production using multiple feedstocks. Curr. Opin. Biotechnol. 2018, 50, 57–64. [Google Scholar] [CrossRef]
- Rütering, M.; Cress, B.F.; Schilling, M.; Rühmann, B.; Koffas, M.A.G.; Sieber, V.; Schmid, J. Tailor-made exopolysaccharides—CRISPR-Cas9 mediated genome editing in Paenibacillus polymyxa. Synth. Biol. 2017, 2, ysx007. [Google Scholar] [CrossRef]
- Pedreira, T.; Elfmann, C.; Stülke, J. The current state of Subti Wiki, the database for the model organism Bacillus subtilis. Nucleic Acids Res. 2022, 50, D875–D882. [Google Scholar] [CrossRef]
- Hartig, S.M. Basic Image Analysis and Manipulation in ImageJ. Curr. Protoc. Mol. Biol. 2013, 102, 14.15.1–14.15.12. [Google Scholar] [CrossRef]
- Karava, M. Development of a Platform for Immobilization of Proteins Based on Bacillus subtilis Spores; Technische Universität Darmstadt: Darmstadt, Germany, 2021. [Google Scholar]
- Zhang, Y.; Ji, F.; Wang, J.; Pu, Z.; Jiang, B.; Bao, Y. Purification and characterization of a novel organic solvent-tolerant and cold-adapted lipase from Psychrobacter sp. ZY124. Extremophiles 2018, 22, 287–300. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 50, D20–D26. [Google Scholar] [CrossRef]
- Hwang, B.-Y.; Pan, J.-G.; Kim, B.-G.; Kim, J.-H. Functional Display of Active Tetrameric β-Galactosidase Using Bacillus subtilis Spore Display System. J. Nanosci. Nanotechnol. 2013, 13, 2313–2319. [Google Scholar] [CrossRef]
- Schilling, C.; Ciccone, R.; Sieber, V.; Schmid, J. Engineering of the 2,3-butanediol pathway of Paenibacillus polymyxa DSM 365. Metab. Eng. 2020, 61, 381–388. [Google Scholar] [CrossRef]
- Meliawati, M.; Schilling, C.; Schmid, J. Recent advances of Cas12a applications in bacteria. Appl. Microbiol. Biotechnol. 2021, 105, 2981–2990. [Google Scholar] [CrossRef]
- Parks, T.D.; Howard, E.D.; Wolpert, T.J.; Arp, D.J.; Dougherty, W.G. Expression and Purification of a Recombinant Tobacco Etch Virus NIa Proteinase: Biochemical Analyses of the Full-Length and a Naturally Occurring Truncated Proteinase Form. Virology 1995, 210, 194–201. [Google Scholar] [CrossRef]
- van Pouderoyen, G.; Eggert, T.; Jaeger, K.-E.; Dijkstra, B.W. The crystal structure of Bacillus subtilis lipase: A minimal α/β hydrolase fold enzyme1†1Edited by R. Huber†This paper is dedicated to the memory of Professor Charles Colson, Louvain-la-Neuve, who initiated the molecular research on Bacillus subtilis lipase. J. Mol. Biol. 2001, 309, 215–226. [Google Scholar]
- Martin, N.; Lombardía, E.; Altabe, S.G.; de Mendoza, D.; Mansilla, M.C. A lipA (yutB) Mutant, Encoding Lipoic Acid Synthase, Provides Insight into the Interplay between Branched-Chain and Unsaturated Fatty Acid Biosynthesis in Bacillus subtilis. J. Bacteriol. 2009, 191, 7447–7455. [Google Scholar] [CrossRef]
- Babtie, A.; Tokuriki, N.; Hollfelder, F. What makes an enzyme promiscuous? Curr. Opin. Chem. Biol. 2010, 14, 200–207. [Google Scholar] [CrossRef]
- Dartois, V.; Baulard, A.; Schanck, K.; Colson, C. Cloning, nucleotide sequence and expression in Escherichia coli of a lipase gene from Bacillus subtilis 168. Biochim. Biophys. Acta BBA—Gene Struct. Expr. 1992, 1131, 253–260. [Google Scholar] [CrossRef]
- Noby, N.; Saeed, H.; Embaby, A.M.; Pavlidis, I.V.; Hussein, A. Cloning, expression and characterization of cold active esterase (EstN7) from Bacillus cohnii strain N1: A novel member of family IV. Int. J. Biol. Macromol. 2018, 120, 1247–1255. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, S.; Gao, H.; Hu, N. Characterization of a cold-active esterase from Serratia sp. and improvement of thermostability by directed evolution. BMC Biotechnol. 2016, 16, 7. [Google Scholar] [CrossRef]
- Wei, W.; Ma, J.; Chen, S.-Q.; Cai, X.-H.; Wei, D.-Z. A novel cold-adapted type I pullulanase of Paenibacillus polymyxa Nws-pp2: In vivo functional expression and biochemical characterization of glucans hydrolyzates analysis. BMC Biotechnol. 2015, 15, 96. [Google Scholar] [CrossRef]
- Mattossovich, R.; Iacono, R.; Cangiano, G.; Cobucci-Ponzano, B.; Isticato, R.; Moracci, M.; Ricca, E. Conversion of xylan by recyclable spores of Bacillus subtilis displaying thermophilic enzymes. Microb. Cell Factories 2017, 16, 218. [Google Scholar] [CrossRef]
- Chen, L.; Mulchandani, A.; Ge, X. Spore-displayed enzyme cascade with tunable stoichiometry. Biotechnol. Prog. 2017, 33, 383–389. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Y.; Yang, R. Recent progress in Bacillus subtilis spore-surface display: Concept, progress, and future. Appl. Microbiol. Biotechnol. 2017, 101, 933–949. [Google Scholar] [CrossRef]
- Liu, X.; Han, R.; Cao, Y.; Turner, B.L.; Ma, L.Q. Enhancing Phytate Availability in Soils and Phytate-P Acquisition by Plants: A Review. Environ. Sci. Technol. 2022, 56, 9196–9219. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zander, M.; Schmid, J.; Kabisch, J. Implementation of Spore Display in Paenibacillus polymyxa with Different Hydrolytic Enzymes. Microorganisms 2024, 12, 1438. https://doi.org/10.3390/microorganisms12071438
Zander M, Schmid J, Kabisch J. Implementation of Spore Display in Paenibacillus polymyxa with Different Hydrolytic Enzymes. Microorganisms. 2024; 12(7):1438. https://doi.org/10.3390/microorganisms12071438
Chicago/Turabian StyleZander, Maximilian, Jochen Schmid, and Johannes Kabisch. 2024. "Implementation of Spore Display in Paenibacillus polymyxa with Different Hydrolytic Enzymes" Microorganisms 12, no. 7: 1438. https://doi.org/10.3390/microorganisms12071438
APA StyleZander, M., Schmid, J., & Kabisch, J. (2024). Implementation of Spore Display in Paenibacillus polymyxa with Different Hydrolytic Enzymes. Microorganisms, 12(7), 1438. https://doi.org/10.3390/microorganisms12071438