
The Microbiome of Fertilization-Stage Maize Silks (Style) Encodes Genes and Expresses Traits That Potentially Promote Survival in Pollen/Style Niches and Host Reproduction
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Methodology Overview
2.2. In Vitro Testing of Microbial Traits
2.3. Whole-Genome Sequencing, Gene Annotation, and Mining
3. Results
3.1. Overview of Whole-Genome Mining and In Vitro Testing for Traits Relevant to the Silk Environment
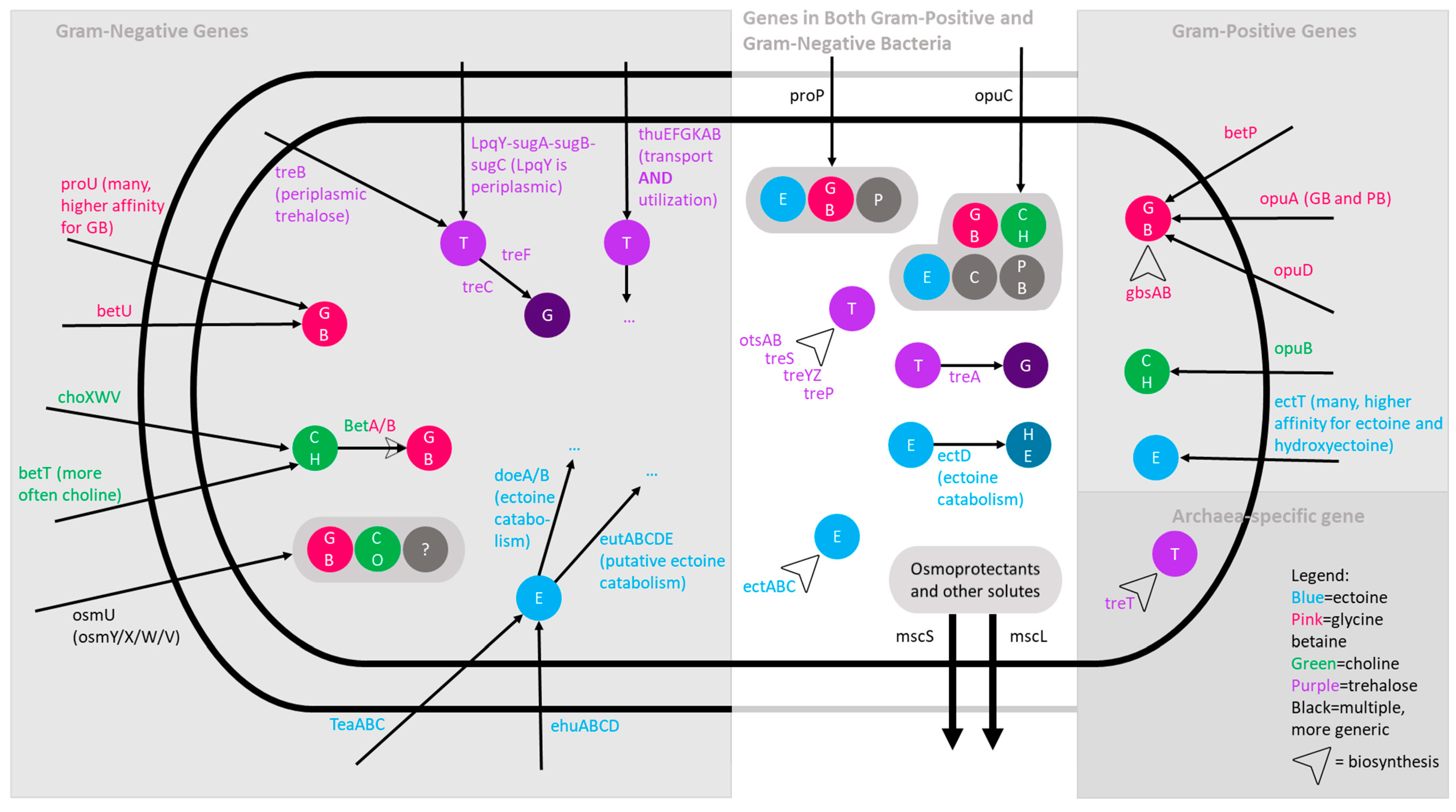
3.2. Water Limitation-Associated Traits
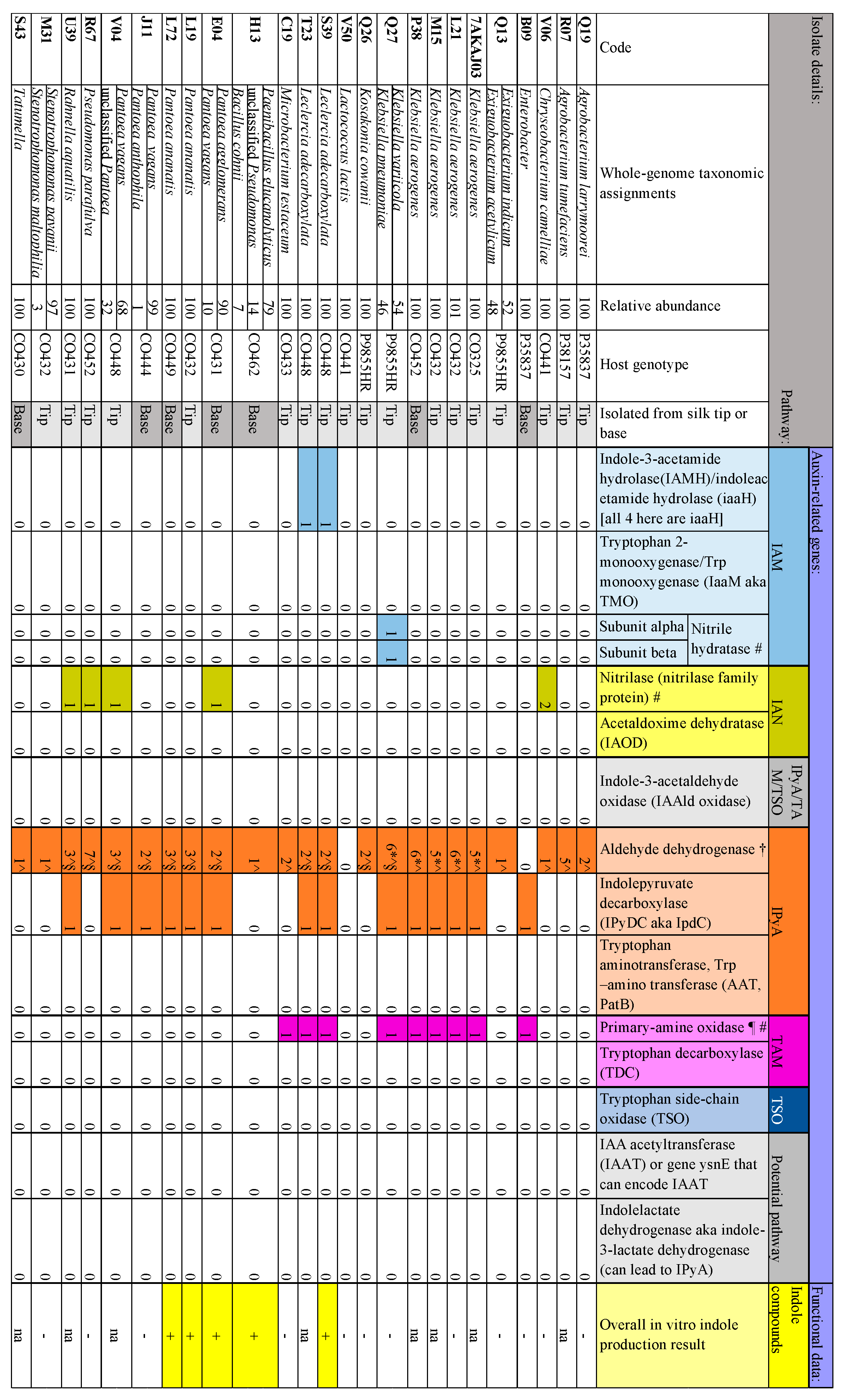
3.3. Indole Compound-Associated Traits (Auxin Biosynthesis)
3.4. Nitrogen Limitation Traits
3.5. Pollen/Fertilization Signaling Traits
3.6. Aluminum Tolerance Traits
4. Discussion
4.1. Overview
4.2. TSM Traits in the Context of Vertical Transmission
4.3. Tolerance of the TSM to Silk/Pollen Abiotic Stresses
4.3.1. Desiccation Tolerance
4.3.2. Nitrogen Limitation
4.3.3. Aluminum Tolerance
4.3.4. Acid Tolerance
4.4. Evidence for Possible Selection on the TSM for Signaling Compounds That Promote Host Reproduction
4.4.1. Auxin Biosynthesis
4.4.2. GABA
4.4.3. ACC Deaminase
4.4.4. Nitric Oxide
4.5. Some TSM Isolates Show Multiple Adaptations
4.6. Prior Studies Show Pantoea Can Benefit Plants and Be Vertically Transmitted
4.7. Study Limitations and Future Experiments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sauter, M. A Guided Tour: Pollen Tube Orientation in Flowering Plants. Chin. Sci. Bull. 2009, 54, 2376–2382. [Google Scholar] [CrossRef]
- Kiesselbach, T. The Structure and Reproduction of Corn; Brown, D., Schaefer, S., Eds.; 50th Anniv.; Cold Spring Harbour Laboratory Press: Cold Spring Harbour, NY, USA, 1999. [Google Scholar]
- Turc, O.; Bouteillé, M.; Fuad-Hassan, A.; Welcker, C.; Tardieu, F. The Growth of Vegetative and Reproductive Structures (Leaves and Silks) Respond Similarly to Hydraulic Cues in Maize. New Phytol. 2016, 212, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, E.M.; Shrestha, A.; Rinne, J.; Lynch, M.D.J.; Shearer, C.R.; Limay-Rios, V.; Reid, L.M.; Raizada, M.N. Transmitting Silks of Maize Have a Complex and Dynamic Microbiome. Sci. Rep. 2021, 11, 13215. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.E.H.; Shrestha, A.; Rinne, J.; Limay-Rios, V.; Reid, L.; Raizada, M.N. The Cultured Microbiome of Pollinated Maize Silks Shifts after Infection with Fusarium graminearum and Varies by Distance from the Site of Pathogen Inoculation. Pathogens 2023, 12, 1322. [Google Scholar] [CrossRef] [PubMed]
- Diniz, G.F.D.; Figueiredo, J.E.F.; Lana, U.G.P.; Marins, M.S.; Silva, D.D.; Cota, L.V.; Marriel, I.E.; Oliveira-Paiva, C.A. Microorganisms from Corn Stigma with Biocontrol Potential of Fusarium verticillioides. Braz. J. Biol. 2022, 82, e262567. [Google Scholar] [CrossRef] [PubMed]
- Johnston-Monje, D.; Raizada, M.N. Conservation and Diversity of Seed Associated Endophytes in Zea across Boundaries of Evolution, Ethnography and Ecology. PLoS ONE 2011, 6, e20396. [Google Scholar] [CrossRef] [PubMed]
- Rijavec, T.; Lapanje, A.; Dermastia, M.; Rupnik, M. Isolation of Bacterial Endophytes from Germinated Maize Kernels. Can. J. Microbiol. 2007, 53, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Sheibani-Tezerji, R.; Naveed, M.; Jehl, M.A.; Sessitsch, A.; Rattei, T.; Mitter, B. The Genomes of Closely Related Pantoea ananatis Maize Seed Endophytes Having Different Effects on the Host Plant Differ in Secretion System Genes and Mobile Genetic Elements. Front. Microbiol. 2015, 6, 440. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zuo, S.; Xu, L.; Zou, Y.; Song, W. Study on Diversity of Endophytic Bacterial Communities in Seeds of Hybrid Maize and Their Parental Lines. Arch. Microbiol. 2012, 194, 1001–1012. [Google Scholar] [CrossRef]
- Kim, H.; Jeon, J.; Lee, K.K.; Lee, Y.H. Longitudinal Transmission of Bacterial and Fungal Communities from Seed to Seed in Rice. Commun. Biol. 2022, 5, 772. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Z.; Zhang, R.; Xu, T.; Zhao, J.; Liu, Y. Diversity of Endophytic Bacteria in Hybrid Maize Seeds and Bacillus mojavensis J2416-7 May Be Capable of Vertical Transmission. Arch. Microbiol. 2022, 204, 213. [Google Scholar] [CrossRef]
- Hodgson, S.; de Cates, C.; Hodgson, J.; Morley, N.J.; Sutton, B.C.; Gange, A.C. Vertical Transmission of Fungal Endophytes Is Widespread in Forbs. Ecol. Evol. 2014, 4, 1199–1208. [Google Scholar] [CrossRef]
- Zampino, D.; Duro, A.; Sciandrello, S.; Parafati, L.; Restuccia, C. Pollen Viability and Endophytic Yeast Species of Cistus creticus and C. monspeliensis. Plant Biosyst. 2020, 155, 384–393. [Google Scholar] [CrossRef]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial Seed Endophytes: Genera, Vertical Transmission and Interaction with Plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal Bacteria Play a Role in Mating Preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef]
- Theis, K.R.; Venkataraman, A.; Dycus, J.A.; Koonter, K.D.; Schmitt-Matzen, E.N.; Wagner, A.P.; Holekamp, K.E.; Schmidt, T.M. Symbiotic Bacteria Appear to Mediate Hyena Social Odors. Proc. Natl. Acad. Sci. USA 2013, 110, 19832–19837. [Google Scholar] [CrossRef]
- Pacini, E.; Dolferus, R. Pollen Developmental Arrest: Maintaining Pollen Fertility in a World with a Changing Climate. Front. Plant Sci. 2019, 10, 679. [Google Scholar] [CrossRef]
- Kostić, A.; Kaluđerović, L.M.; Dojčinović, B.P.; Barać, M.B.; Babić, V.B.; Mačukanović-Jocić, M.P. Preliminary Investigation of Mineral Content of Pollen Collected from Different Serbian Maize Hybrids–Is There Any Potential Nutritional Value? J. Sci. Food Agric. 2017, 97, 2803–2809. [Google Scholar] [CrossRef]
- Feijó, J.A.; Sainhas, J.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Growing Pollen Tubes Possess a Constitutive Alkaline Band in the Clear Zone and a Growth-Dependent Acidic Tip. J. Cell Biol. 1999, 144, 483. [Google Scholar] [CrossRef]
- Hepler, P.K.; Rounds, C.M.; Winship, L.J. Control of Cell Wall Extensibility during Pollen Tube Growth. Mol. Plant 2013, 6, 998–1017. [Google Scholar] [CrossRef]
- Tupý, J.; Řhová, L. Changes and Growth Effect of pH in Pollen Tube Culture. J. Plant Physiol. 1984, 115, 1–10. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y.; Reger, B.J.; Heslop-Harrison, J. The Pollen-Stigma Interaction in the Grasses. 6. The Stigma ('silk’) of Zea mays L. as Host to the Pollens of Sorghum bicolor (L.) Moench and Pennisetum americanum (L.) Leeke. Acta Bot. Neerl. 1984, 33, 205–227. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Lausser, A.; Márton, M.L. Using Maize as a Model to Study Pollen Tube Growth and Guidance, Cross-Incompatibility and Sperm Delivery in Grasses. Ann. Bot. 2011, 108, 727–737. [Google Scholar] [CrossRef]
- Uhart, S.A.; Andrade, F.H. Nitrogen Deficiency in Maize: II. Carbon-Nitrogen Interaction Effects on Kernel Number and Grain Yield. Crop Sci. 1995, 35, 1384–1389. [Google Scholar] [CrossRef]
- Uhart, S.A.; Andrade, F.H. Nitrogen Deficiency in Maize: I. Effects on Crop Growth, Development, Dry Matter Partitioning, and Kernel Set. Crop Sci. 1995, 35, 1376–1383. [Google Scholar] [CrossRef]
- Borrás, L.; Vitantonio-Mazzini, L.N. Maize Reproductive Development and Kernel Set under Limited Plant Growth Environments. J. Exp. Bot. 2018, 69, 3235–3243. [Google Scholar] [CrossRef]
- Thompson, M.E.H. Discovery and Testing of Pollinated Maize Silk-Associated Microbes Including Microbiome Assisted Selection of Biocontrol Agents against Fusarium graminearum. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2024. [Google Scholar]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in Situ Assay for Indoleacetic Acid Production by Bacteria Immobilized on a Nitrocellulose Membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef]
- Hernández-Fernández, G.; Galán, B.; Carmona, M.; Castro, L.; García, J. Transcriptional Response of the Xerotolerant Arthrobacter sp. Helios Strain to PEG-Induced Drought Stress. bioRxiv 2022. [Google Scholar] [CrossRef]
- Latif, M.; Bukhari, S.A.H.; Alrajhi, A.A.; Alotaibi, F.S.; Ahmad, M.; Shahzad, A.N.; Dewidar, A.Z.; Mattar, M.A. Inducing Drought Tolerance in Wheat through Exopolysaccharide-Producing Rhizobacteria. Agronomy 2022, 12, 1140. [Google Scholar] [CrossRef]
- Huang, S.; Wang, X.; Liu, X.; He, G.; Wu, J. Isolation, Identification, and Characterization of an Aluminum-Tolerant Bacterium Burkholderia sp. SB1 from an Acidic Red Soil. Pedosphere 2018, 28, 905–912. [Google Scholar] [CrossRef]
- Lim, J.C.; Goh, K.M.; Shamsir, M.S.; Ibrahim, Z.; Chong, C.S. Characterization of Aluminum Resistant Anoxybacillus sp. SK 3–4 Isolated from a Hot Spring. J. Basic. Microbiol. 2015, 55, 514–519. [Google Scholar] [CrossRef]
- Herrero, M.P.; Johnson, R.R. Drought Stress and Its Effects on Maize Reproductive Systems. Crop Sci. 1981, 21, 105–110. [Google Scholar] [CrossRef]
- Wood, J.M. Osmosensing by Bacteria. Sci. STKE 2006, 2006, pe43. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. Bacterial Osmoadaptation: The Role of Osmolytes in Bacterial Stress and Virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef]
- Christgen, S.L.; Becker, D.F. Role of Proline in Pathogen and Host Interactions. Antioxid. Redox Signal 2019, 30, 683–709. [Google Scholar] [CrossRef]
- Meadows, J.A.; Correspondence, M.J.W.; Wargo, M.J. Carnitine in Bacterial Physiology and Metabolism. Microbiology 2015, 161, 1161–1174. [Google Scholar] [CrossRef]
- Booth, I.R.; Blount, P. The MscS and MscL Families of Mechanosensitive Channels Act as Microbial Emergency Release Valves. J. Bacteriol. 2012, 194, 4802–4809. [Google Scholar] [CrossRef]
- Padilla, L.; Morbach, S.; Krämer, R.; Agosin, E. Impact of Heterologous Expression of Escherichia coli UDP-Glucose Pyrophosphorylase on Trehalose and Glycogen Synthesis in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2004, 70, 3845–3854. [Google Scholar] [CrossRef]
- Lee, H.J.; Yoon, Y.S.; Lee, S.J. Mechanism of Neuroprotection by Trehalose: Controversy Surrounding Autophagy Induction. Cell Death Dis. 2018, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Lee, S.J.; Boos, W. TreT, a Novel Trehalose Glycosyltransferring Synthase of the Hyperthermophilic Archaeon Thermococcus litoralis. J. Biol. Chem. 2004, 279, 47890–47897. [Google Scholar] [CrossRef] [PubMed]
- Ruhal, R.; Kataria, R.; Choudhury, B. Trends in Bacterial Trehalose Metabolism and Significant Nodes of Metabolic Pathway in the Direction of Trehalose Accumulation. Microb. Biotechnol. 2013, 6, 493–502. [Google Scholar] [CrossRef]
- Klein, W.; Horlacher, R.; Boos, W. Molecular Analysis of TreB Encoding the Escherichia coli Enzyme II Specific for Trehalose. J. Bacteriol. 1995, 177, 4043–4052. [Google Scholar] [CrossRef]
- Kalera, K.; Stothard, A.I.; Woodruff, P.J.; Swarts, B.M. The Role of Chemoenzymatic Synthesis in Advancing Trehalose Analogues as Tools for Combatting Bacterial Pathogens. Chem. Commun. 2020, 56, 11528–11547. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Weinrick, B.; Veeraraghavan, U.; Besra, G.S.; Jacobs, W.R. Trehalose-Recycling ABC Transporter LpqY-SugA-SugB-SugC Is Essential for Virulence of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 107, 21761–21766. [Google Scholar] [CrossRef]
- Ampomah, O.Y.; Avetisyan, A.; Hansen, E.; Svenson, J.; Huser, T.; Jensen, J.B.; Bhuvaneswari, T.V. The ThuEFGKAB Operon of Rhizobia and Agrobacterium tumefaciens Codes for Transport of Trehalose, Maltitol, and Isomers of Sucrose and Their Assimilation through the Formation of Their 3-Keto Derivatives. J. Bacteriol. 2013, 195, 3797–3807. [Google Scholar] [CrossRef]
- Kuhlmann, A.U.; Bremer, E. Osmotically Regulated Synthesis of the Compatible Solute Ectoine in Bacillus pasteurii and Related Bacillus spp. Appl. Environ. Microbiol. 2002, 68, 772–783. [Google Scholar] [CrossRef]
- Kuhlmann, A.U.; Hoffmann, T.; Bursy, J.; Jebbar, M.; Bremer, E. Ectoine and Hydroxyectoine as Protectants against Osmotic and Cold Stress: Uptake through the SigB-Controlled Betaine-Choline-Carnitine Transporter-Type Carrier Ectt from Virgibacillus pantothenticus. J. Bacteriol. 2011, 193, 4699–4708. [Google Scholar] [CrossRef]
- Kuhlmann, S.I.; Van Scheltinga, A.C.T.; Bienert, R.; Kunte, H.J.; Ziegler, C. 1.55 Å Structure of the Ectoine Binding Protein TeaA of the Osmoregulated TRAP-Transporter TeaABC from Halomonas elongata. Biochemistry 2008, 47, 9475–9485. [Google Scholar] [CrossRef]
- Grammann, K.; Volke, A.; Kunte, H.J. New Type of Osmoregulated Solute Transporter Identified in Halophilic Members of the Bacteria Domain: TRAP Transporter TeaABC Mediates Uptake of Ectoine and Hydroxyectoine in Halomonas elongata DSM 2581T. J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef]
- Schweikhard, E.S.; Kuhlmann, S.I.; Org Kunte, H.-J.; Grammann, K.; Ziegler, C.M. Structure and Function of the Universal Stress Protein TeaD and Its Role in Regulating the Ectoine Transporter TeaABC of Halomonas elongata DSM 2581 T. Biochemistry 2010, 49, 2194–2204. [Google Scholar] [CrossRef]
- Yu, Q.; Cai, H.; Zhang, Y.; He, Y.; Chen, L.; Merritt, J.; Zhang, S.; Dong, Z. Negative Regulation of Ectoine Uptake and Catabolism in Sinorhizobium meliloti: Characterization of the EhuR Gene. J. Bacteriol. 2016, 199, e00119-16. [Google Scholar] [CrossRef]
- Reshetnikov, A.S.; Rozova, O.N.; Trotsenko, Y.A.; But, S.Y.; Khmelenina, V.N.; Mustakhimov, I.I. Ectoine Degradation Pathway in Halotolerant Methylotrophs. PLoS ONE 2020, 15, e0232244. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, S.; Lv, P.; Sun, S.; Ma, C.; Xu, P.; Su, H.; Yang, C. High Ectoine Production by an Engineered Halomonas hydrothermalis Y2 in a Reduced Salinity Medium. Microb. Cell Fact. 2019, 18, 184. [Google Scholar] [CrossRef]
- Richter, A.A.; Mais, C.N.; Czech, L.; Geyer, K.; Hoeppner, A.; Smits, S.H.J.; Erb, T.J.; Bange, G.; Bremer, E. Biosynthesis of the Stress-Protectant and Chemical Chaperon Ectoine: Biochemistry of the Transaminase EctB. Front. Microbiol. 2019, 10, 2811. [Google Scholar] [CrossRef]
- Ly, A.; Henderson, J.; Lu, A.; Culham, D.E.; Wood, J.M. Osmoregulatory Systems of Escherichia coli: Identification of Betaine-Carnitine-Choline Transporter Family Member BetU and Distributions of BetU and TrkG among Pathogenic and Nonpathogenic Isolates. J. Bacteriol. 2004, 186, 296–306. [Google Scholar] [CrossRef]
- Rübenhagen, R.; Rönsch, H.; Jung, H.; Krämer, R.; Morbach, S. Osmosensor and Osmoregulator Properties of the Betaine Carrier BetP from Corynebacterium glutamicum in Proteoliposomes. J. Biol. Chem. 2000, 275, 735–741. [Google Scholar] [CrossRef]
- Frossard, S.M.; Khan, A.A.; Warrick, E.C.; Gately, J.M.; Hanson, A.D.; Oldham, M.L.; Sanders, D.A.; Csonka, L.N. Identification of a Third Osmoprotectant Transport System, the OsmU System, in Salmonella enterica. J. Bacteriol. 2012, 194, 3861–3871. [Google Scholar] [CrossRef]
- Kempf, B.; Bremer, E. OpuA, an Osmotically Regulated Binding Protein-Dependent Transport System for the Osmoprotectant Glycine Betaine in Bacillus subtilis. J. Biol. Chem. 1995, 270, 16701–16713. [Google Scholar] [CrossRef]
- Ziegler, C.; Bremer, E.; Krämer, R. The BCCT Family of Carriers: From Physiology to Crystal Structure. Mol. Microbiol. 2010, 78, 13–34. [Google Scholar] [CrossRef]
- Boncompagni, E.; Østerås, M.; Poggi, M.-C.; Le Rudulier, D. Occurrence of Choline and Glycine Betaine Uptake and Metabolism in the Family Rhizobiaceae and Their Roles in Osmoprotection. Appl. Environ. Microbiol. 1999, 65, 2072–2077. [Google Scholar] [CrossRef]
- Pocard, J.A.; Vincent, N.; Boncompagni, E.; Tombras Smith, L.; Poggi, M.C.; Le Rudulier, D. Molecular Characterization of the Bet Genes Encoding Glycine Betaine Synthesis in Sinorhizobium meliloti 102F34. Microbiology (N Y) 1997, 143, 1369–1379. [Google Scholar] [CrossRef]
- Aragón-Aranda, B.; Palacios-Chaves, L.; Salvador-Bescós, M.; de Miguel, M.J.; Muñoz, P.M.; Vences-Guzmán, M.Á.; Zúñiga-Ripa, A.; Lázaro-Antón, L.; Sohlenkamp, C.; Moriyón, I.; et al. The Phospholipid N-Methyltransferase and Phosphatidylcholine Synthase Pathways and the ChoXWV Choline Uptake System Involved in Phosphatidylcholine Synthesis Are Widely Conserved in Most, but Not All Brucella Species. Front. Microbiol. 2021, 12, 614243. [Google Scholar] [CrossRef]
- Rath, H.; Reder, A.; Hoffmann, T.; Hammer, E.; Seubert, A.; Bremer, E.; Völker, U.; Mäder, U. Management of Osmoprotectant Uptake Hierarchy in Bacillus subtilis via a SigB-Dependent Antisense RNA. Front. Microbiol. 2020, 11, 622. [Google Scholar] [CrossRef]
- MacMillan, S.V.; Alexander, D.A.; Culham, D.E.; Kunte, H.J.; Marshall, E.V.; Rochon, D.; Wood, J.M. The Ion Coupling and Organic Substrate Specificities of Osmoregulatory Transporter ProP in Escherichia coli. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1420, 30–44. [Google Scholar] [CrossRef]
- Culham, D.E.; Lasby, B.; Marangoni, A.G.; Milner, J.L.; Steer, B.A.; Van Nues, R.W.; Wood, J.M. Isolation and Sequencing of Escherichia coli Gene ProP Reveals Unusual Structural Features of the Osmoregulatory Proline/Betaine Transporter, ProP. J. Mol. Biol. 1993, 229, 268–276. [Google Scholar] [CrossRef]
- Jebbar, M.; Talibart, R.; Gloux, K.; Bernard, T.; Blanco, C. Osmoprotection of Escherichia coli by Ectoine: Uptake and Accumulation Characteristics. J. Bacteriol. 1992, 174, 5027–5035. [Google Scholar] [CrossRef]
- Wood, J.M. Proline Porters Effect the Utilization of Proline as Nutrient or Osmoprotectant for Bacteria. J. Membr. Biol. 1988, 106, 183–202. [Google Scholar] [CrossRef]
- Mól, R.; Filek, M.; Machačkova, I.; Matthys-Rochon, E. Ethylene Synthesis and Auxin Augmentation in Pistil Tissues Are Important for Egg Cell Differentiation after Pollination in Maize. Plant Cell Physiol. 2004, 45, 1396–1405. [Google Scholar] [CrossRef]
- Li, X.M.; Sang, Y.L.; Zhao, X.Y.; Zhang, X.S. High-Throughput Sequencing of Small RNAs from Pollen and Silk and Characterization of MiRNAs as Candidate Factors Involved in Pollen-Silk Interactions in Maize. PLoS ONE 2013, 8, 72852. [Google Scholar] [CrossRef]
- Duca, D.R.; Glick, B.R. Indole-3-Acetic Acid Biosynthesis and Its Regulation in Plant-Associated Bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, S.P.; Cueto, L.; García-Estrada, C.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Singh, S.P.; Angel Blázquez, M.; Sansinenea, E. Auxins of Microbial Origin and Their Use in Agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 8549–8565. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Harper, C.P. The Roles of Auxin during Interactions between Bacterial Plant Pathogens and Their Hosts. J. Exp. Bot. 2018, 69, 245–254. [Google Scholar] [CrossRef]
- Morffy, N.; Strader, L.C. Old Town Roads: Routes of Auxin Biosynthesis across Kingdoms. Curr. Opin. Plant Biol. 2020, 55, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Li, S.; Zhang, N.; Cui, X.; Zhou, X.; Zhang, G.; Shen, Q.; Zhang, R. Analysis and Cloning of the Synthetic Pathway of the Phytohormone Indole-3-Acetic Acid in the Plant-Beneficial Bacillus amyloliquefaciens SQR9. Microb. Cell Fact. 2015, 14, 130. [Google Scholar] [CrossRef]
- Shao, J.; Li, Y.; Li, Z.; Xu, Z.; Xun, W.; Zhang, N.; Feng, H.; Miao, Y.; Shen, Q.; Zhang, R. Participating Mechanism of a Major Contributing Gene YsnE for Auxin Biosynthesis in Bacillus amyloliquefaciens SQR9. J. Basic. Microbiol. 2021, 61, 569–575. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-Acetic Acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef]
- Spaepen, S.; Versées, W.; Gocke, D.; Pohl, M.; Steyaert, J.; Vanderleyden, J. Characterization of Phenylpyruvate Decarboxylase, Involved in Auxin Production of Azospirillum brasilense. J. Bacteriol. 2007, 189, 7626–7633. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and Plant-Microbe Interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef]
- Duca, D.; Rose, D.R.; Glick, B.R. Characterization of a Nitrilase and a Nitrile Hydratase from Pseudomonas sp. Strain UW4 That Converts Indole-3-Acetonitrile to Indole-3-Acetic Acid. Appl. Environ. Microbiol. 2014, 80, 4640–4649. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Bacterial Biosynthesis of Indole-3-Acetic Acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef]
- Pace, H.C.; Brenner, C. The Nitrilase Superfamily: Classification, Structure and Function. Genome Biol. 2001, 2, reviews0001.1–reviews0001.9. [Google Scholar] [CrossRef]
- Koga, J.; Adachi, T.; Hidaka, H. Molecular Cloning of the Gene for Indolepyruvate Decarboxylase from Enterobacter cloacae. Mol. General. Genet. 1991, 226, 10–16. [Google Scholar] [CrossRef]
- Idris, E.S.E.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-Dependent Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth Promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant-Microbe Interact. 2007, 20, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Perley, J.E.; Stowe, B.B. On the Ability of Taphrina deformans to Produce Indoleacetic Acid from Tryptophan by Way of Tryptamine. Plant Physiol. 1966, 41, 234–237. [Google Scholar] [CrossRef]
- Oberhansli, T.; Defago, G.; Haas, D. Indole-3-Acetic Acid (IAA) Synthesis in the Biocontrol Strain CHA0 of Pseudomonas fluorescens: Role of Tryptophan Side Chain Oxidase. J. Gen. Microbiol. 1991, 137, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- McClerklin, S.A.; Lee, S.G.; Harper, C.P.; Nwumeh, R.; Jez, J.M.; Kunkel, B.N. Indole-3-Acetaldehyde Dehydrogenase-Dependent Auxin Synthesis Contributes to Virulence of Pseudomonas syringae Strain DC3000. PLoS Pathog. 2018, 14, e1006811. [Google Scholar] [CrossRef] [PubMed]
- Debruin, J.L.; Hemphill, B.; Schussler, J.R. Silk Development and Kernel Set in Maize as Related to Nitrogen Stress. Crop Sci. 2018, 58, 2581–2592. [Google Scholar] [CrossRef]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen Tube Growth and Guidance Is Regulated by POP2, an Arabidopsis Gene That Controls GABA Levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The Versatile GABA in Plants. Plant Signal Behav. 2021, 16, e1862565. [Google Scholar] [CrossRef]
- Phuengjayaem, S.; Booncharoen, A.; Tanasupawat, S. Characterization and Comparative Genomic Analysis of Gamma-Aminobutyric Acid (GABA)-Producing Lactic Acid Bacteria from Thai Fermented Foods. Biotechnol. Lett. 2021, 43, 1637–1648. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA Production and Structure of GadB/GadC Genes in Lactobacillus and Bifidobacterium Strains from Human Microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L. Microbial Response to Acid Stress: Mechanisms and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef]
- McCraw, S.L.; Park, D.H.; Jones, R.; Bentley, M.A.; Rico, A.; Ratcliffe, R.G.; Kruger, N.J.; Collmer, A.; Preston, G.M. GABA (g-Aminobutyric Acid) Uptake via the GABA Permease GabP Represses Virulence Gene Expression in Pseudomonas syringae Pv. Tomato DC3000. Mol. Plant-Microbe Interact. 2016, 29, 938–949. [Google Scholar] [CrossRef]
- Pandey, S.; Ranade, S.A.; Nagar, P.K.; Kumar, N. Role of Polyamines and Ethylene as Modulators of Plant Senescence. J. Biosci. 2000, 25, 291–299. [Google Scholar] [CrossRef]
- Dar, R.A.; Nisar, S.; Tahir, I. Ethylene: A Key Player in Ethylene Sensitive Flower Senescence: A Review. Sci. Hortic. 2021, 290, 110491. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Bacterial Modulation of Plant Ethylene Levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef]
- Glick, B.R. Modulation of Plant Ethylene Levels by the Bacterial Enzyme ACC Deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef]
- Moon, Y.S.; Ali, S. Possible Mechanisms for the Equilibrium of ACC and Role of ACC Deaminase-Producing Bacteria. Appl. Microbiol. Biotechnol. 2022, 106, 877–887. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.C. Plant Growth Promotion under Water: Decrease of Waterlogging-Induced ACC and Ethylene Levels by ACC Deaminase-Producing Bacteria. Front. Microbiol. 2018, 9, 1096. [Google Scholar] [CrossRef]
- Zhou, L.Z.; Dresselhaus, T. Friend or Foe: Signaling Mechanisms during Double Fertilization in Flowering Seed Plants. Curr. Top. Dev. Biol. 2019, 131, 453–496. [Google Scholar] [CrossRef]
- Prado, A.M.; Porterfield, D.M.; Feijó, J.A. Nitric Oxide Is Involved in Growth Regulation and Re-Orientation of Pollen Tubes. Development 2004, 131, 2707–2714. [Google Scholar] [CrossRef]
- Sudhamsu, J.; Crane, B.R. Bacterial Nitric Oxide Synthases: What Are They Good For? Trends Microbiol. 2009, 17, 212–218. [Google Scholar] [CrossRef]
- Filippovich, S.Y. Bacterial NO Synthases. Biochemistry 2010, 75, 1217–1224. [Google Scholar] [CrossRef]
- Arai, H.; Kodama, T.; Igarashi, Y. Effect of Nitrogen Oxides on Expression of the nir and nor Genes for Denitrification in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1999, 170, 19–24. [Google Scholar] [CrossRef]
- Ambika Manirajan, B.; Ratering, S.; Rusch, V.; Schwiertz, A.; Geissler-Plaum, R.; Cardinale, M.; Schnell, S. Bacterial Microbiota Associated with Flower Pollen Is Influenced by Pollination Type, and Shows a High Degree of Diversity and Species-Specificity. Environ. Microbiol. 2016, 18, 5161–5174. [Google Scholar] [CrossRef]
- Barra, P.J.; Viscardi, S.; Jorquera, M.A.; Duran, P.A.; Valentine, A.J.; De la Luz Mora, M. Understanding the Strategies to Overcome Phosphorus-Deficiency and Aluminum-Toxicity by Ryegrass Endophytic and Rhizosphere Phosphobacteria. Front. Microbiol. 2018, 9, 1155. [Google Scholar] [CrossRef]
- Ligaba, A.; Shen, H.; Shibata, K.; Yamamoto, Y.; Tanakamaru, S.; Matsumoto, H. The Role of Phosphorus in Aluminium-Induced Citrate and Malate Exudation from Rape (Brassica napus). Physiol. Plant 2004, 120, 575–584. [Google Scholar] [CrossRef]
- Wang, Q.F.; Zhao, Y.; Yi, Q.; Li, K.Z.; Yu, Y.X.; Chen, L.M. Overexpression of Malate Dehydrogenase in Transgenic Tobacco Leaves: Enhanced Malate Synthesis and Augmented Al-Resistance. Acta Physiol. Plant 2010, 32, 1209–1220. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Naamala, J.; Dakora, F.D. Nature and Mechanisms of Aluminium Toxicity, Tolerance and Amelioration in Symbiotic Legumes and Rhizobia. Biol. Fertil. Soils 2018, 54, 309–318. [Google Scholar] [CrossRef]
- Rubio-Sanz, L.; Prieto, R.I.; Imperial, J.; Palacios, J.M.; Brito, B. Functional and Expression Analysis of the Metal-Inducible DmeRF System from Rhizobium leguminosarum Bv. Viciae. Appl. Environ. Microbiol. 2013, 79, 6414–6422. [Google Scholar] [CrossRef]
- Beris, F.S.; De Smet, L.; Karaoglu, H.; Canakci, S.; Van Beeumen, J.; Belduz, A.O. The ATPase Activity of the G2alt Gene Encoding an Aluminium Tolerance Protein from Anoxybacillus gonensis G2. J. Microbiol. 2011, 49, 641–650. [Google Scholar] [CrossRef]
- Jo, J.; Jang, Y.S.; Kim, K.Y.; Kim, M.H.; Kim, I.J.; Chung, W. Il Isolation of ALU1-P Gene Encoding a Protein with Aluminum Tolerance Activity from Arthrobacter viscosus. Biochem. Biophys. Res. Commun. 1997, 239, 835–839. [Google Scholar] [CrossRef]
- Feijó, J.A.; Malhó, R.; Obermeyer, G. Ion Dynamics and Its Possible Role during in Vitro Pollen Germination and Tube Growth. Protoplasma 1995, 187, 155–167. [Google Scholar] [CrossRef]
- Sahu, P.K.; Mishra, S. Effect of Hybridization on Endophytes: The Endo-Microbiome Dynamics. Symbiosis 2021, 84, 369–377. [Google Scholar] [CrossRef]
- Seeley, T.D. The Honey Bee Colony as a Superorganism. Am. Sci. 1989, 77, 546–553. [Google Scholar]
- Chichiriccò, G. Post-shedding Dehydration and in Vivo Temporal Germination of Crocus Pollen. Grana 2007, 44, 142–147. [Google Scholar] [CrossRef]
- Flis, S.E.; Glenn, A.R.; Dilworth, M.J. The Interaction between Aluminium and Root Nodule Bacteria. Soil. Biol. Biochem. 1993, 25, 403–417. [Google Scholar] [CrossRef]
- Holdaway-Clarke, T.L.; Hepler, P.K. Control of Pollen Tube Growth: Role of Ion Gradients and Fluxes. New Phytol. 2003, 159, 539–563. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takai, K. Sulfur Metabolisms in Epsilon-and Gamma-Proteobacteria in Deep-Sea Hydrothermal Fields. Front. Microbiol. 2011, 2, 192. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Maruyama, F.; Ogram, A.V.; Navarrete, O.U.; Lagos, L.M.; Inostroza, N.G.; Acuña, J.J.; Rilling, J.I.; de La Luz Mora, M. Rhizobacterial Community Structures Associated with Native Plants Grown in Chilean Extreme Environments. Microb. Ecol. 2016, 72, 633–646. [Google Scholar] [CrossRef]
- Rudnick, P.; Meletzus, D.; Green, A.; He, L.; Kennedy, C. Regulation of Nitrogen Fixation by Ammonium in Diazotrophic Species of Proteobacteria. Soil. Biol. Biochem. 1997, 29, 831–841. [Google Scholar] [CrossRef]
- Czech, L.; Gertzen, C.; Smits, S.H.J.; Bremer, E. Guilty by Association: Importers, Exporters and MscS-Type Mechanosensitive Channels Encoded in Biosynthetic Gene Clusters for the Stress-Protectant Ectoine. Environ. Microbiol. 2022, 24, 5306–5331. [Google Scholar] [CrossRef]
- Vandrich, J.; Pfeiffer, F.; Alfaro-Espinoza, G.; Kunte, H.J. Contribution of Mechanosensitive Channels to Osmoadaptation and Ectoine Excretion in Halomonas elongata. Extremophiles 2020, 24, 421–432. [Google Scholar] [CrossRef]
- Salgado-Aguilar, M.; Molnar, T.; Pons-Hernández, J.L.; Covarrubias-Prieto, J.; Ramírez-Pimentel, J.G.; Raya-Pérez, J.C.; Hearne, S.; Iturriaga, G.; Salgado-Aguilar, M.; Molnar, T.; et al. Physiological and Biochemical Analyses of Novel Drought-Tolerant Maize Lines Reveal Osmoprotectant Accumulation at Silking Stage. Chil. J. Agric. Res. 2020, 80, 241–252. [Google Scholar] [CrossRef]
- De Rudder, K.E.E.; Sohlenkamp, C.; Geiger, O. Plant-Exuded Choline Is Used for Rhizobial Membrane Lipid Biosynthesis by Phosphatidylcholine Synthase. J. Biol. Chem. 1999, 274, 20011–20016. [Google Scholar] [CrossRef]
- Peel, G.J.; Mickelbart, M.V.; Rhodes, D. Choline Metabolism in Glycinebetaine Accumulating and Non-Accumulating near-Isogenic Lines of Zea mays and Sorghum bicolor. Phytochemistry 2010, 71, 404–414. [Google Scholar] [CrossRef]
- Masterson, R.V.; Russell, P.R.; Atherly, A.G. Nitrogen Fixation (nif) Genes and Large Plasmids of Rhizobium japonicum. J. Bacteriol. 1982, 152, 928. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martínez, L.; Silva, J.; Martínez-Romero, E. Klebsiella variicola, a Novel Species with Clinical and Plant-Associated Isolates. Syst. Appl. Microbiol. 2004, 27, 27–35. [Google Scholar] [CrossRef]
- Yang, L.; Yang, K. Biological Function of Klebsiella variicola and Its Effect on the Rhizosphere Soil of Maize Seedlings. PeerJ 2020, 8, e9894. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Gutiérrez, J.P.; Lopez-Lavalle, L.A.B. Seed-Transmitted Bacteria and Fungi Dominate Juvenile Plant Microbiomes. Front. Microbiol. 2021, 12, 2945. [Google Scholar] [CrossRef]
- Dumigan, C.R.; Muileboom, J.; Gregory, J.; Shrestha, A.; Hewedy, O.A.; Raizada, M.N. Ancient Relatives of Modern Maize from the Center of Maize Domestication and Diversification Host Endophytic Bacteria That Confer Tolerance to Nitrogen Starvation. Front. Plant Sci. 2021, 12, 1908. [Google Scholar] [CrossRef]
- Hoover, T.; Imperial, J.; Ludden, P.; Shah, V. Biosynthesis of the Iron-Molybdenum Cofactor of Nitrogenase. Biofactors 1988, 1, 199–205. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Huang, S.; Li, L.; Gao, Q.; Wang, Y.; Zhang, S.; Huang, S.; Yuan, L.; Wen, Y.; et al. A Highly Conserved Core Bacterial Microbiota with Nitrogen-Fixation Capacity Inhabits the Xylem Sap in Maize Plants. Nat. Commun. 2022, 13, 3361. [Google Scholar] [CrossRef]
- Farh, M.E.A.; Kim, Y.J.; Sukweenadhi, J.; Singh, P.; Yang, D.C. Aluminium Resistant, Plant Growth Promoting Bacteria Induce Overexpression of Aluminium Stress Related Genes in Arabidopsis thaliana and Increase the Ginseng Tolerance against Aluminium Stress. Microbiol. Res. 2017, 200, 45–52. [Google Scholar] [CrossRef]
- Shi, Q.; Jin, J.; Liu, Y.; Zhang, Y.; Cai, Z.; Ma, Q.; Cheng, Y.; Wen, R.; Nian, H.; Lian, T. High Aluminum Drives Different Rhizobacterial Communities between Aluminum-Tolerant and Aluminum-Sensitive Wild Soybean. Front. Microbiol. 2020, 11, 1996. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, X.Q.; Aizawa, T.; Sunairi, M.; Shen, R.F. High Aluminum Tolerance of Rhodotorula sp. RS1 Is Associated with Thickening of the Cell Wall Rather than Chelation of Aluminum Ions. Pedosphere 2013, 23, 29–38. [Google Scholar] [CrossRef]
- Noller, A.C.; McEllistrem, M.C.; Stine, O.C.; Morris, J.G.; Boxrud, D.J.; Dixon, B.; Harrison, L.H. Multilocus Sequence Typing Reveals a Lack of Diversity among Escherichia coli O157:H7 Isolates That Are Distinct by Pulsed-Field Gel Electrophoresis. J. Clin. Microbiol. 2003, 41, 675–679. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Raizada, M.N. Interactions between Co-Habitating Fungi Elicit Synthesis of Taxol from an Endophytic Fungus in Host Taxus Plants. Front. Microbiol. 2013, 4, 3. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Greenwood, J.S.; Bombarely, A.; Mueller, L.A.; Tsao, R.; Mosser, D.D.; Raizada, M.N. An Endophyte Constructs Fungicide-Containing Extracellular Barriers for Its Host Plant. Curr. Biol. 2015, 25, 2570–2576. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Yu, F.; Chen, X.; Zhao, H.; Li, H.; Wu, J. Indole-3-Acetic Acid Biosynthesis Pathways in the Plant-Beneficial Bacterium Arthrobacter pascens ZZ21. Int. J. Mol. Sci. 2018, 19, 443. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of Gaba (γ-Aminobutyric Acid) by Microorganisms: A Review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef]
- Lang, J.; Gonzalez-Mula, A.; Taconnat, L.; Clement, G.; Faure, D. The Plant GABA Signaling Downregulates Horizontal Transfer of the Agrobacterium tumefaciens Virulence Plasmid. New Phytol. 2016, 210, 974–983. [Google Scholar] [CrossRef]
- Crane, B.R.; Sudhamsu, J.; Patel, B.A. Bacterial Nitric Oxide Synthases. Annu. Rev. Biochem. 2010, 79, 445–470. [Google Scholar] [CrossRef]
- He, J.M.; Bai, X.L.; Wang, R.B.; Cao, B.; She, X.P. The Involvement of Nitric Oxide in Ultraviolet-B-Inhibited Pollen Germination and Tube Growth of Paulownia tomentosa in Vitro. Physiol. Plant 2007, 131, 273–282. [Google Scholar] [CrossRef]
- Crane, B.R. The Enzymology of Nitric Oxide in Bacterial Pathogenesis and Resistance. Biochem. Soc. Trans. 2008, 36, 1149–1154. [Google Scholar] [CrossRef]
- Mishra, A.; Chauhan, P.S.; Chaudhry, V.; Tripathi, M.; Nautiyal, C.S. Rhizosphere Competent Pantoea agglomerans Enhances Maize (Zea mays) and Chickpea (Cicer arietinum L.) Growth, without Altering the Rhizosphere Functional Diversity. Antonie Van Leeuwenhoek 2011, 100, 405–413. [Google Scholar] [CrossRef]
- Gond, S.K.; Torres, M.S.; Bergen, M.S.; Helsel, Z.; White, J.F. Induction of Salt Tolerance and Up-Regulation of Aquaporin Genes in Tropical Corn by Rhizobacterium Pantoea agglomerans. Lett. Appl. Microbiol. 2015, 60, 392–399. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, J.H.; Kang, B.R.; Rong, X.; Gardener, B.B.M.; Ji, H.J.; Park, C.S.; Kim, Y.C. Draft Genome Sequence of Pantoea ananatis B1-9, a Nonpathogenic Plant Growth-Promoting Bacterium. J. Bacteriol. 2012, 194, 729. [Google Scholar] [CrossRef]
- Montañez, A.; Abreu, C.; Gill, P.R.; Hardarson, G.; Sicardi, M. Biological Nitrogen Fixation in Maize (Zea mays L.) by 15N Isotope-Dilution and Identification of Associated Culturable Diazotrophs. Biol. Fertil. Soils 2009, 45, 253–263. [Google Scholar] [CrossRef]
- Dutkiewicz, J.; Mackiewicz, B.; Lemieszek, M.K.; Golec, M.; Milanowski, J. Pantoea agglomerans: A Mysterious Bacterium of Evil and Good. Part IV. Beneficial Effects. Ann. Agric. Environ. Med. 2016, 23, 206–222. [Google Scholar] [CrossRef]
- Sammer, U.F.; Reiher, K.; Spiteller, D.; Wensing, A.; Völksch, B. Assessment of the Relevance of the Antibiotic 2-Amino-3-(Oxirane-2,3-Dicarboxamido)-Propanoyl-Valine from Pantoea agglomerans Biological Control Strains against Bacterial Plant Pathogens. Microbiologyopen 2012, 1, 438–449. [Google Scholar] [CrossRef]
- Pusey, P.L.; Stockwell, V.O.; Reardon, C.L.; Smits, T.H.M.; Duffy, B. Antibiosis Activity of Pantoea agglomerans Biocontrol Strain E325 against Erwinia amylovora on Apple Flower Stigmas. Phytopathology 2011, 101, 1234–1241. [Google Scholar] [CrossRef]
- Cañamás, T.P.; Viñas, I.; Abadias, M.; Usall, J.; Torres, R.; Teixidó, N. Acid Tolerance Response Induced in the Biocontrol Agent Pantoea agglomerans CPA-2 and Effect on Its Survival Ability in Acidic Environments. Microbiol. Res. 2009, 164, 438–450. [Google Scholar] [CrossRef]
- Deroo, W.; De Troyer, L.; Dumoulin, F.; De Saeger, S.; De Boevre, M.; Vandenabeele, S.; De Gelder, L.; Audenaert, K. A Novel in Planta Enrichment Method Employing Fusarium graminearum-Infected Wheat Spikes to Select for Competitive Biocontrol Bacteria. Toxins 2022, 14, 222. [Google Scholar] [CrossRef]
- Brady, C.; Cleenwerck, I.; Venter, S.; Vancanneyt, M.; Swings, J.; Coutinho, T. Phylogeny and Identification of Pantoea Species Associated with Plants, Humans and the Natural Environment Based on Multilocus Sequence Analysis (MLSA). Syst. Appl. Microbiol. 2008, 31, 447–460. [Google Scholar] [CrossRef]
- Gaoa, J.; Xue, J.; Yan, H.; Tong, S.; Khan, M.S.; Wang, L.; Mao, X.; Zhang, X.; Sun, J. Pantoea endophytica sp. Nov., Novel Endophytic Bacteria Isolated from Maize Planting in Different Geographic Regions of Northern China. Syst. Appl. Microbiol. 2019, 42, 488–494. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, K.; Liao, H.; Hector, S.B.; Shi, X.; Li, J.; Liu, B.; Xu, T.; Tong, C.; Liu, X.; et al. Genomic and Secretomic Insight into Lignocellulolytic System of an Endophytic Bacterium Pantoea ananatis Sd-1. Biotechnol. Biofuels 2016, 9, 25. [Google Scholar] [CrossRef]
- Sarhan, M.S.; Hamza, M.A.; Youssef, H.H.; Patz, S.; Becker, M.; ElSawey, H.; Nemr, R.; Daanaa, H.S.A.; Mourad, E.F.; Morsi, A.T.; et al. Culturomics of the Plant Prokaryotic Microbiome and the Dawn of Plant-Based Culture Media—A Review. J. Adv. Res. 2019, 19, 15–27. [Google Scholar] [CrossRef]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. PlasmidSPAdes: Assembling Plasmids from Whole Genome Sequencing Data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef]
- Rahman, N.A.; Wan Rosli, W.I. Nutritional Compositions and Antioxidative Capacity of the Silk Obtained from Immature and Mature Corn. J. King Saud. Univ. Sci. 2014, 26, 119–127. [Google Scholar] [CrossRef]
- Ha, A.W.; Kang, H.J.; Kim, S.L.; Kim, M.H.; Kim, W.K. Acute and Subacute Toxicity Evaluation of Corn Silk Extract. Prev. Nutr. Food Sci. 2018, 23, 70. [Google Scholar] [CrossRef]
- Hasanudin, K.; Hashim, P.; Mustafa, S. Corn Silk (Stigma maydis) in Healthcare: A Phytochemical and Pharmacological Review. Molecules 2012, 17, 9697–9715. [Google Scholar] [CrossRef]
- Nassar, F.S.; Alsahlawi, A.M.; E Abdellatif, H.A.; Kamel, N.N.; Abbas, A.O. Dietary Corn Silk (Stigma maydis) Extract Supplementation Modulate Production Performance, Immune Response and Redox Balance in Corticosterone-Induced Oxidative Stress Broilers. Animals 2023, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Rios, V.L.; Brettingham, D.J.L.; Raizada, M. Bacteria Existing in Pre-Pollinated Styles (Silks) Can Defend the Exposed Male Gamete Fertilization Channel of Maize against an Environmental Fusarium Pathogen. Front. Plant Sci. 2023, 14, 1292109. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Rios, V.L.; Brettingham, D.J.L.; Raizada, M. Maize Pollen Carry Bacteria That Suppress a Fungal Pathogen That Enters through the Male Gamete Fertilization Route. Front. Plant Sci. 2024, 14, 1286199. [Google Scholar] [CrossRef]
- Wu, G.; Chen, J.; Shi, X.; Kim, J.S.; Xia, J.; Zhang, L. Impacts of Global Climate Warming on Meteorological and Hydrological Droughts and Their Propagations. Earths Future 2022, 10, e2021EF002542. [Google Scholar] [CrossRef]
- Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.A.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global Changes in Drought Conditions under Different Levels of Warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
- Van Loon, J.; Speratti, A.B.; Gabarra, L.; Govaerts, B.; Batán, E.; de Mexico, E. Precision for Smallholder Farmers: A Small-Scale-Tailored Variable Rate Fertilizer Application Kit. Agriculture 2018, 8, 48. [Google Scholar] [CrossRef]
- Thar, S.P.; Farquharson, R.J.; Ramilan, T.; Coggins, S.; Chen, D. Recommended vs. Practice: Smallholder Fertilizer Decisions in Central Myanmar. Agriculture 2021, 11, 65. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, H.; Pan, X.; Jin, W.; Li, X. Aberrant Meiotic Modulation Partially Contributes to the Lower Germination Rate of Pollen Grains in Maize (Zea mays L.) under Low Nitrogen Supply. Plant Cell Physiol. 2017, 58, 342–353. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, M.E.H.; Raizada, M.N. The Microbiome of Fertilization-Stage Maize Silks (Style) Encodes Genes and Expresses Traits That Potentially Promote Survival in Pollen/Style Niches and Host Reproduction. Microorganisms 2024, 12, 1473. https://doi.org/10.3390/microorganisms12071473
Thompson MEH, Raizada MN. The Microbiome of Fertilization-Stage Maize Silks (Style) Encodes Genes and Expresses Traits That Potentially Promote Survival in Pollen/Style Niches and Host Reproduction. Microorganisms. 2024; 12(7):1473. https://doi.org/10.3390/microorganisms12071473
Chicago/Turabian StyleThompson, Michelle E. H., and Manish N. Raizada. 2024. "The Microbiome of Fertilization-Stage Maize Silks (Style) Encodes Genes and Expresses Traits That Potentially Promote Survival in Pollen/Style Niches and Host Reproduction" Microorganisms 12, no. 7: 1473. https://doi.org/10.3390/microorganisms12071473