
Evaluation of Resistance Induction Promoted by Bioactive Compounds of Pseudomonas aeruginosa LV Strain against Asian Soybean Rust
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production and Extract of Microbial Bioactive Compounds
2.2. Field Experiment
2.2.1. Experimental Area
2.2.2. Weed Control
2.2.3. Effect of F4A Fraction on Asian Soybean Rust
2.3. Metabolomic Analysis
2.3.1. Sample Preparation
2.3.2. Metabolomic Analysis
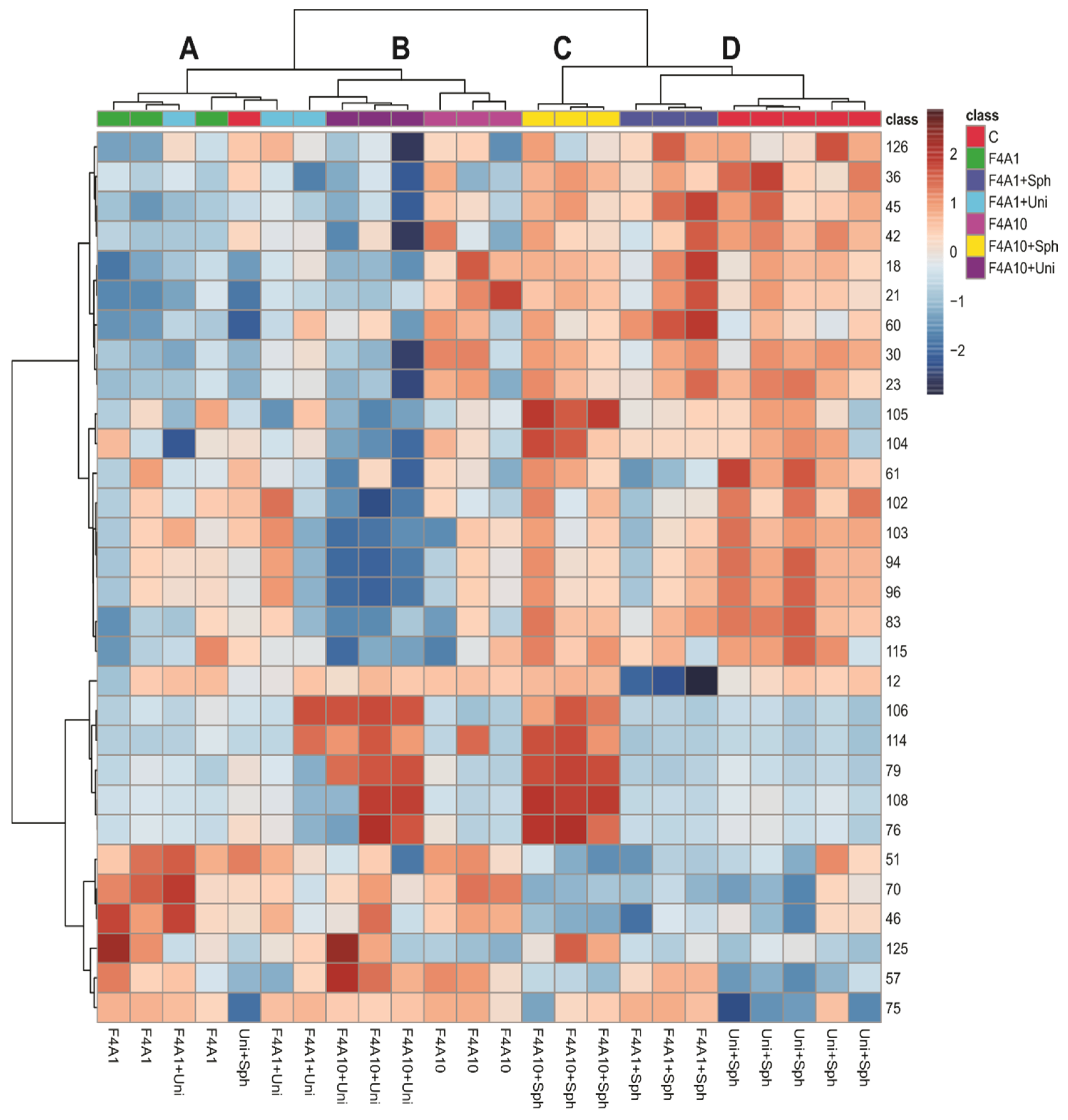
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conab. Acompanhamento da Safra Brasileira de Grãos (Monitoring the Brazilian Grain Harvest). 2023. Available online: http://www.conab.gov.br (accessed on 12 July 2024).
- Philis, G.; Gracey, E.O.; Gansel, L.C.; Fet, A.M.; Rebours, C. Comparing the primary energy and phosphorus consumption of soybean and seaweed-based aquafeed proteins—A material and substance flow analysis. Clean. Prod. 2018, 200, 1142–1153. [Google Scholar] [CrossRef]
- Langenbach, C.; Campe, R.; Beyer, S.F.; Mueller, A.N.; Conrath, U. Fighting Asian soybean rust. Front. Plant Sci. 2016, 7, 797. [Google Scholar] [CrossRef]
- Hoppe, H.H.; Koch, E. Defense reactions in host and nonhost plants against the soybean rust fungus (Phakopsora pachyrhizi Syd.). J. Phytopathol. 1989, 125, 77–88. [Google Scholar] [CrossRef]
- Koch, E.; Ebrahim Nesbat, F.; Hoppe, H. Light and electron microscopic studies on the development of soybean rust (Phakopsora pachyrhizi Syd.) in susceptible soybean leaves. Phytopathol. Z. 1983, 106, 302–320. [Google Scholar] [CrossRef]
- Vittal, R.; Paul, C.; Hill, C.B.; Hartman, G.L. Characterization and quantification of fungal colonization of Phakopsora pach-yrhizi in soybean genotypes. Phytopathology 2014, 104, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Hartman, G.L.; Rupe, J.C.; Sikora, E.J.; Domier, L.L.; Davis, J.A.; Steffey, K.L. (Eds.) Compendium of Soybean Diseases and Pests, 5th ed.; The American Phytopathological Society: Saint Paul, MN, USA, 2015. [Google Scholar] [CrossRef]
- Zambolim, L.; Juliatti, F.C.; Guerra, W. How to cope with the vulnerability of site specific fungicides on the control of Asian soybean rust. Int. J. Res. Agron. 2021, 4, 14–25. [Google Scholar] [CrossRef]
- Nascimento, R.S.M.; Ferreira, L.R.; Zambolim, L.; Parreira, D.F.; da Costa, Y.K.S.; Damascena, J.F.; Cecon, P.R. Spray mixture volume in the control of Asian soybean rust. Crop Prot. 2021, 146, 105662. [Google Scholar] [CrossRef]
- FAO. Pesticides Use, Pesticides Trade and Pesticides Indicators. 2022. Available online: https://www.fao.org/documents/card/en?details=cc0918en (accessed on 23 April 2024).
- Godoy, C.V.; Flausino, A.M.; Santos, L.C.M.; del Ponte, E.M. Eficiência do controle da ferrugem asiática da soja em função do momento de aplicação sob condições de epidemia em Londrina, PR (Efficiency of Asian soybean rust control depending on the time of application under epidemic conditions in Londrina, PR). Trop. Plant Pathol. 2009, 34, 56–61. [Google Scholar] [CrossRef]
- Godoy, C.V.; Seixas, C.D.S.; Soares, R.M.; Marcelino-Guimarães, F.C.; Meyer, M.C.; Costamilan, L.M. Asian soybean rust in Brazil: Past, present, and future. Pesqui. Agropecu. Bras. 2016, 51, 407–421. [Google Scholar] [CrossRef]
- Delaney, M.; ArchMiller, A.A.; Delaney, D.P.; Wilson, A.E.; Sikora, E.J. Effectiveness of Fungicide on Soybean Rust in the Southeastern United States: A Meta-Analysis. Sustainability 2018, 10, 1784. [Google Scholar] [CrossRef]
- Scherm, H.; Christiano, R.S.C.; Esker, P.D.; Del Ponte, E.M.; Godoy, C.V. Quantitative review of fungicide efficacy trials for managing soybean rust in Brazil. Crop Prot. 2009, 28, 774–782. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 1, 315. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant Systemic Acquired Resistance by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Fritig, B.; Heitz, T.; Legrand, M. Antimicrobial proteins in induced plant defense. Curr. Opin. Immunol. 1998, 10, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Van Strien, E.A. The families of Pathogenesis-Related Proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Barka, E.A. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia Sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced Systemic Resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Castaldi, S.; Masi, M.; Sautua, F.; Cimmino, A.; Isticato, R.; Carmona, M.; Tuzi, A.; Evidente, A. Pseudomonas fluorescens showing antifungal activity against Macrophomina phaseolina, a severe pathogenic fungus of soybean, produces phenazine as the main active metabolite. Biomolecules 2021, 11, 1728. [Google Scholar] [CrossRef]
- Morales, D.K.; Jacobs, N.J.; Rajamani, S.; Krishnamurthy, M.; Cubillos-Ruiz, J.R.; Hogan, D.A. Antifungal mechanisms by which a novel Pseudomonas aeruginosa phenazine toxin kills Candida albicans in biofilms. Mol. Microbiol. 2010, 78, 1379–1392. [Google Scholar] [CrossRef]
- Clough, S.E.; Jousset, A.; Elphinstone, J.G.; Friman, V.P. Combining in vitro and in vivo screening to identify efficient Pseu-domonas biocontrol strains against the phytopathogenic bacterium Ralstonia solanacearum. Microbiologyopen 2022, 11, e1283. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D. Combined use of Trichoderma and beneficial bacteria (mainly Bacillus and Pseudomonas): Development of microbial synergistic bio-inoculants in sustainable agriculture. Biol. Control 2022, 176, 105100. [Google Scholar] [CrossRef]
- Simionato, A.S.; Navarro, M.O.P.; Barazetti, A.R.; dos Santos, I.M.O.; Spago, F.R.; Chryssafidis, A.L.; Andrade, G. Strategies for biological control and antagonisms. Plant-Microbe Interact. Agro-Ecol. Perspect. 2017, 2, 221–244. [Google Scholar] [CrossRef]
- Cardozo, V.F.; Oliveira, A.G.; Nishio, E.K.; Perugini, M.R.E.; Andrade, C.G.T.J.; Silveira, W.D.; Durán, N.; Andrade, G.; Kobayashi, R.K.T.; Nakazato, G. Antibacterial activity of extracellular compounds produced by a Pseudomonas strain against methicillin-resistant Staphylococcus aureus (MRSA) strains. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.G.; Spago, F.R.; Simionato, A.S.; Navarro, M.O.P.; Da Silva, C.S.; Barazetti, A.R.; Cely, M.V.T.; Tischer, C.A.; San Martin, J.A.B.; De Jesus Andrade, C.G.T.; et al. Bioactive organocopper compound from Pseudomonas aeruginosa inhibits the growth of Xanthomonas citri subsp. citri. Front. Microbiol. 2016, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.P.; Oliveira, A.G., Jr.; Beranger, J.P.O.; Góis, C.G.; Vasconcellos, F.C.S.; Martin, J.A.B.S.; Andrade, C.G.T.J.; Mello, J.C.P.; Andrade, G. Activity of extracellular compounds of Pseudomonas sp. against Xanthomonas axonopodis in vitro and bacterial leaf blight in Eucalyptus. Trop. Plant Pathol. 2012, 37, 233–238. [Google Scholar] [CrossRef]
- Pistori, J.F.; Simionato, A.S.; Navarro, M.O.P.; Andreata, M.F.L.; Santos, I.M.O.; Meneguim, L.; Leite Junior, R.P.; Oliveira, A.G.; Andrade, G. Low-molecular-weight metabolites produced by Pseudomonas aeruginosa as an alternative to control Huanglongbing in Citrus sinensis Cv. Valencia. Trop. Plant Pathol. 2018, 43, 289–296. [Google Scholar] [CrossRef]
- Munhoz, L.D.; Fonteque, J.P.; Santos, I.M.O.; Navarro, M.O.P.; Simionato, A.S.; Goya, E.T.; Rezende, M.I.; Balbi-Peña, M.I.; de Oliveira, A.G.; Andrade, G. Control of bacterial stem rot on tomato by extracellular bioactive compounds produced by Pseudomonas aeruginosa LV strain. Cogent Food Agric. 2017, 3, 1282592. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources 2006: A Framework for International Classification, Correlation and Communication; World Soil Resources Reports; Cambridge University Press: Cambridge, UK, 2006; Volume 43. [Google Scholar] [CrossRef]
- Köppen, W. Die wärmezonen der Erde, nach der dauer der heissen, gemässigten und kalten zeit und nach der wirkung der wärme auf die organische welt betrachtet (The heat zones of the Earth are considered according to the duration of the hot, temperate and cold periods and the effect of heat on the organic world). Meteorol. Z. 1884, 215–226. Available online: http://koeppen-geiger.vu-wien.ac.at/pdf/Koppen_1884.pdf (accessed on 12 July 2024).
- IDR-Paraná. Dados Meteorológicos Históricos e Atuais (Historical and Current Weather Data). Available online: https://www.idrparana.pr.gov.br/Pagina/Dados-Meteorologicos-Historicos-e-Atuais (accessed on 28 March 2024).
- Cely, M.V.T.; Siviero, M.A.; Emiliano, J.; Spago, F.R. Inoculation of Schizolobium parahyba with mycorrhizal fungi and plant growth-promoting rhizobacteria increases wood yield under field conditions. Front. Plant Sci. 2016, 7, 1708. [Google Scholar] [CrossRef]
- Vukics, V.; Guttman, A. Structural characterization of flavonoid glycosides by multi-stage mass spectrometry. Mass Spectrom. Rev. 2010, 29, 1–16. [Google Scholar] [CrossRef]
- Macedo, A.L.; Boaretto, A.G.; Silva, A.N.; Maia, D.S.; Siqueira, J.M.; Silva, D.B.; Carollo, C.A. Evaluation of the Effect of Brazilian Savanna (Cerrado) Seasons in Flavonoids and Alkaloids Accumulation: The Case of Duguetia furfuracea. J. Braz. Chem. Soc. 2021, 32, 1840–1850. [Google Scholar] [CrossRef]
- Savarino, P.; Demeyer, M.; Decroo, C.; Colson, E.; Gerbaux, P. Mass spectrometry analysis of saponins. Mass Spectrom. Rev. 2023, 42, 954–983. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yu, Y.; Zhou, Z.; Zhou, J.M. Plant pattern-recognition receptors controlling innate immunity. Sci. China Life Sci. 2016, 59, 878–888. [Google Scholar] [CrossRef]
- Gullino, M.L.; Tinivella, F.; Garibaldi, A.; Kemmitt, G.M.; Bacci, L.; Sheppard, B. Mancozeb: Past, present, and future. Plant Dis. 2010, 94, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, M.N.; Coleman, T.; Giang, P.D.; Wang, R.; Bernhardt, P.V.; De Voss, J.J.; Bell, S.G. To be, or not to be, an inhibitor: A comparison of azoles interactions with and oxidation by a Cytochrome P450 enzyme. Inorg. Chem. 2022, 61, 236–245. [Google Scholar] [CrossRef]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Monk, B.C.; Tyndall, J.D.A. Structural insights into binding of the antifungal drug fluconazole to Saccharomyces cerevisiae lanosterol 14α-demethylase. Antimicrob. Agents Chemother. 2015, 59, 4982–4989. [Google Scholar] [CrossRef]
- Pontzen, R.; Scheinpflug, H. Effects of triazole fungicides on sterol biosynthesis during spore germination of Botrytis cinerea, Venturia inaequalis and Puccinia graminis f. sp. Tritici. Neth. J. Plant Pathol. 1989, 95, 151–160. [Google Scholar] [CrossRef]
- Twizeyimana, M.; Hartman, G.L. Sensitivity of Phakopsora pachyrhizi isolates to fungicides and reduction of fungal infection based on fungicide and timing of application. Plant Dis. 2017, 101, 121–128. [Google Scholar] [CrossRef]
- Venancio, W.S.; Rodrigues, M.A.T.; Begliomini, E.; Souza, N.L. Efeitos fisiológicos de fungicida do grupo das estrobirulinas sobre plantas (Physiological effects of fungicides from the strobirulin group on plants). Publ. UEPG Ci. Exatas Terra Ci. Agr. Eng. 2003, 9, 59–68. [Google Scholar]
- Barazetti, A.R.; Simionato, A.S.; Navarro, M.O.P.; dos Santos, I.M.O.; Modolon, F.; de Lima Andreata, M.F.; Liuti, G.; Cely, M.V.T.; Chryssafidi, A.L.; Dealis, M.L.; et al. Formulations of arbuscular mycorrhizal fungi inoculum applied to soybean and corn plants under controlled and field conditions. Appl. Soil Ecol. 2019, 142, 25–33. [Google Scholar] [CrossRef]
- Beckers, G.J.; Conrath, U. Priming for stress resistance: From the lab to the field. Curr. Opin. Plant Biol. 2007, 10, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Herms, S.; Seehaus, K.; Koehle, H.; Conrath, U. A Strobilurin fungicide enhances the resistance of tobacco against Tobacco mosaic virus and Pseudomonas syringae Pv Tabaci. Plant Physiol. 2002, 130, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Skandalis, N.; Dimopoulou, A.; Beri, D.; Tzima, A.; Malandraki, I.; Theologidis, I.; Bitivanos, S.; Varveri, C.; Klitsinaris, T.; Vassilakos, N. Effect of pyraclostrobin application on viral and bacterial diseases of tomato. Plant Dis. 2016, 100, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Trdá, L.; Janda, M.; Macková, D.; Pospíchalová, R.; Dobrev, P.I.; Burketová, L.; Matušinsky, P. Dual mode of the saponin aescin in plant protection: Antifungal agent and plant defense elicitor. Front. Plant Sci. 2019, 10, 1448. [Google Scholar] [CrossRef] [PubMed]
- Perry, E.K.; Newman, D.K. Prevalence and correlates of phenazine resistance in culturable bacteria from a dryland wheat field. Appl. Environ. Microbiol. 2022, 88, e0232021. [Google Scholar] [CrossRef] [PubMed]
- Pierson, L.S.; Pierson, E.A. Metabolism and function of phenazines in bacteria: Impacts on the behavior of bacteria in the environment and biotechnological processes. Appl. Microbiol. Biotechnol. 2010, 86, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- De Vleesschauwer, D.; Cornelis, P.; Höfte, M. Redox-active pyocyanin secreted by Pseudomonas aeruginosa 7NSK2 triggers systemic resistance to Magnaporthe grisea but enhances Rhizoctonia solani susceptibility in rice. Mol. Plant Microbe Interact. 2006, 19, 1406–1419. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, I.M.O.; Abe, V.Y.; de Carvalho, K.; Barazetti, A.R.; Simionato, A.S.; de Almeida Pega, G.E.; Matis, S.H.; Cano, B.G.; Cely, M.V.T.; Marcelino-Guimarães, F.C.; et al. Secondary metabolites of Pseudomonas aeruginosa LV strain decrease Asian soybean rust severity in experimentally infected plants. Plants 2021, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.; da Graça, J.P.; Porto, C.; Martin do Prado, R.; Hoffmann-Campo, C.B.; Meyer, M.C.; de Oliveira Nunes, E.; Pilau, E.J. Unraveling Asian soybean rust metabolomics using mass spectrometry and molecular networking approach. Sci. Rep. 2020, 10, 138. [Google Scholar] [CrossRef]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Kim, H.J.; Suh, H.J.; Kim, J.H.; Park, S.; Joo, Y.C.; Kim, J.S. Antioxidant activity of glyceollins derived from soybean elicited with Aspergillus sojae. J. Agric. Food Chem. 2010, 58, 11633–11638. [Google Scholar] [CrossRef]
- Kim, H.J.; Lim, J.-S.; Kim, W.-K.; Kim, J.-S. Soyabean glyceollins: Biological effects and relevance to human health. Proc. Nutr. Soc. 2012, 71, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Li, N.; Dong, L.; Zhang, D.; Fan, S.; Jiang, L.; Wang, X.; Xu, P.; Zhang, S. Overexpression of soybean isoflavone reductase (GmIFR) enhances resistance to Phytophthora sojae in soybean. Front. Plant Sci. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tu, Y.C.; Lian, T.W.; Hung, J.T.; Yen, J.H.; Wu, M.J. Distinctive antioxidant and antiinflammatory effects of flavonols. J. Agric. Food Chem. 2006, 54, 9798–9804. [Google Scholar] [CrossRef]
- Heijnen, C.G.M.; Haenen, G.R.M.M.; Van Acker, F.A.A.; Van Der Vijgh, W.J.F.; Bast, A. Flavonoids as peroxynitrite scavengers: The role of the hydroxyl groups. Toxicol. Vitr. 2001, 15, 3–6. [Google Scholar] [CrossRef]
- Fidelis, Q.C.; Faraone, I.; Russo, D.; Aragão Catunda, F.E., Jr.; Vignola, L.; de Carvalho, M.G.; de Tommasi, N.; Milella, L. Chemical and biological insights of Ouratea hexasperma (A. St.-Hil.) Baill.: A source of bioactive compounds with multifunctional properties. Nat. Prod. Res. 2018, 33, 1500–1503. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Min, C.W.; Kramer, K.; Agrawal, G.K.; Rakwal, R.; Park, K.H.; Wang, Y.; Finkemeier, I.; Kim, S.T. A multi-omics analysis of Glycine Max leaves reveals alteration in flavonoid and isoflavonoid metabolism upon ethylene and abscisic acid treatment. Proteomics 2018, 18, e1700366. [Google Scholar] [CrossRef]
- Lynch, D.V.; Dunn, T.M. An introduction to plant sphingolipids and a review of recent advances in understanding their metabolism and function. New Phytol. 2004, 161, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Luttgeharm, K.D.; Chen, M.; Mehra, A.; Cahoon, R.E.; Markham, J.E.; Cahoon, E.B. Overexpression of Arabidopsis ceramide synthases differentially affects growth, sphingolipid metabolism, programmed cell death, and mycotoxin resistance. Plant Physiol. 2015, 169, 1108–1117. [Google Scholar] [CrossRef]
- Hörtensteiner, S.; Wüthrich, K.L.; Matile, P.; Ongania, K.H.; Kräutler, B. The key step in chlorophyll breakdown in higher plants: Cleavage of pheophorbide a macrocycle by a monooxygenase. J. Biol. Chem. 1998, 273, 15335–15339. [Google Scholar] [CrossRef]
- Kinney, A.J. Development of genetically engineered soybean oils for food applications. J. Food Lipids 1996, 3, 273–292. [Google Scholar] [CrossRef]
- Honna, P.T.; Pagliarini-Fuganti, R.; Ferreira, L.C.; Molinari, M.D.C.; Marin, S.R.R.; de Oliveira, M.C.N.; Farias, J.R.B.; Neumaier, N.; Mertz-Henning, L.M.; Kanamori, N.; et al. Molecular, physiological, and agronomical characterization, in green-house and in field conditions, of soybean plants genetically modified with AtGolS2 gene for drought tolerance. Mol. Breed. 2016, 36, 157. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | Compound | UV (nm) | MF | MS (m/z) [M + H]+ | MS/MS |
|---|---|---|---|---|---|---|
| 13 | 16.03 | O-hexosyl daidzein | 272, 330 sh | C21H20O9 | 417.1169 | 255, 227, 199 |
| 15 | 17.06 | O-deoxyhexosyl di-hexosyl kaempferol | 265, 348 | C33H40O20 | 757.2171 | 287 |
| 17 | 17.40 | O-deoxyhexosyl di-hexosyl kaempferol | 265, 346 | C33H40O20 | 757.2163 | 287 |
| 18 | 17.71 | O-di-hexosyl kaempferol | 265, 345 | C27H30O16 | 611.1602 | 449, 287 |
| 19 | 17.75 | O-di-deoxyhexosyl kaempferol | 265, 345 | C21H20O11 | 449.1088 | 271 |
| 21 | 18.00 | O-di-hexosyl luteolin | 268, 335 | C27H30O16 | 611.1607 | 287 |
| 22 | 18.25 | NI | 280 | C13H20O2 | 209.1529 | - |
| 23 | 18.35 | O-di-deoxyhexosyl hexosyl kaempferol | 265, 347 | C33H40O19 | 741.2251 | 595, 449, 287 |
| 24 | 18.72 | O-di-deoxyhexosyl hexosyl kaempferol | 265, 348 | C33H40O19 | 741.2245 | 595, 449, 287 |
| 25 | 19.03 | O-hexosyl-malonyl daidzein | 275, 330 sh | C24H22O12 | 503.1203 | 255 |
| 26 | 19.11 | O-deoxyhexosyl hexosyl luteolin | 266, 338 | C27H30O15 | 595.1678 | 449, 287 |
| 27 | 19.21 | O-hexosyl genistein | 260, 330 sh | C21H20O10 | 433.1139 | 271 |
| 29 | 19.75 | O-deoxyhexosyl hexosyl kaempferol | 265, 348 | C27H30O15 | 595.1655 | 449, 287, 153 |
| 30 | 20.28 | O-hexosyl kaempferol | 265, 347 | C21H20O11 | 449.1073 | 287 |
| 33 | 20.76 | O-deoxyhexosyl hexosyl kaempferol | 265, 349 | C27H30O15 | 595.1650 | 449, 287, 241, 165 |
| 34 | 21.09 | O-hexosyl kaempferol | 265, 349 | C21H20O11 | 449.1090 | 287 |
| 36 | 21.35 | O-hexosyl-malonyl glycitin | 270, 330 sh | C25H24O13 | 533.1274 | 371, 285 |
| 38 | 21.65 | O-hexosyl apigenin | 268, 335 | C21H20O10 | 433.1147 | 271 |
| 39 | 21.85 | O-pentosyl luteolin | 265, 332 | C20H18O10 | 419.0981 | 287 |
| 40 | 22.20 | O-hexosyl-malonyl genistein | 275, 330 sh | C24H22O13 | 519.1113 | 271 |
| 42 | 23.38 | NI | - | C22H32O10 | 457.2076 | 439, 191, 173 |
| 43 | 23.48 | O-hexosyl-malonyl genistein | 259,330 sh | C24H22O13 | 519.1131 | 271 |
| 44 | 23.79 | NI | - | C22H32O10 | 457.2061 | 439, 191, 173 |
| 45 | 23.85 | O-hexosyl-malonyl naringenin | 280, 330 sh | C24H24O13 | 521.1286 | 273, 153 |
| 48 | 24.99 | O-hexosyl-malonyl apigenin | 267, 334 | C24H22O13 | 519.1126 | 271 |
| 49 | 25.40 | O-hexosyl-malonyl genistein | 275, 330 sh | C24H22O13 | 519.1131 | 271 |
| 53 | 27.19 | O-hexosyl-malonyl afrormosin | 278, 330 sh | C26H26O13 | 547.1498 | 299 |
| 56 | 27.96 | O-hexosyl-malonyl afrormosin | 257, 320 sh | C26H26O13 | 547.1478 | 299 |
| 57 | 28.52 | Genistein | 278, 330 sh | C15H10O5 | 271.0611 | 271, 215 |
| 59 | 29.04 | O-hexosyl-malonyl afrormosin | 280, 328 sh | C26H26O13 | 547.1489 | 299 |
| 60 | 29.35 | Apigenin | 267, 335 | C15H10O5 | 271.0609 | 271, 243, 229, 153 |
| 64 | 30.78 | NI | - | C20H30O5 | 351.2153 | 275 |
| 67 | 32.52 | NI | - | C18H23NO2 | 286.1785 | - |
| 68 | 32.68 | NI | - | C18H25NO2 | 288.1956 | 270, 172, 159 |
| 71 | 32.91 | NI | 280 | C10H18N2O5 | 247.1300 | - |
| 72 | 33.05 | Saponin | - | C51H80O21 | 1029.5243 | |
| 76 | 33.22 | NI | 280, 320 | C16H21NO | 244.1693 | 172, 159 |
| 77 | 33.36 | Sphingolipid (phytosphingosine isomer) | - | C18H39NO3 | 318.2995 | 256 |
| 82 | 33.66 | Saponin (soyaponin ag isomer) | - | C54H84O22 | 1085.5510 | 599, 581, 563, 463, 405, 365, 217, 203 |
| 85 | 33.72 | Saponin | - | C51H80O21 | 1029.5256 | |
| 86 | 33.87 | Soyasaponin I | - | C48H78O18 | 943.5258 | 599, 581, 441, 423, 405, 383, 365, 315, 247, 217, 203 |
| 89 | 34.25 | Saponin | C51H80O21 | 1029.5272 | ||
| 91 | 34.50 | Saponin | - | C55H72O13 | 941.5099 | 597, 439, 421, 410, 381, 313, 273, 245, 219 |
| 94 | 34.84 | Saponin (soyasaponin βg) | - | C54H84O21 | 1069.5546 | 923, 761, 725, 599, 581, 567, 441, 423, 217 |
| 95 | 34.87 | NI | - | C20H28O4 | 333.2047 | - |
| 96 | 34.87 | Saponin (soyasaponin βg) | - | C54H84O21 | 1069.5546 | 923, 761, 725, 599, 581, 567, 441, 423, 217 |
| 100 | 35.26 | Pterocarpan | 280, 325 sh | C21H20O4 | 337.1430 | 279, 267 |
| 103 | 35.46 | Saponin | - | C48H74O17 | 923.4965 | - |
| 105 | 35.89 | Fatty acid derivative | - | C21H36O4 | 353.2691 | - |
| 111 | 36.78 | Fatty acid derivative | - | C18H28O2 | 277.2164 | - |
| 112 | 36.91 | Fatty acid derivative | - | C18H28O2 | 277.2152 | - |
| 115 | 37.33 | Fatty acid derivative | - | C21H36O4 | 353.2691 | - |
| 119 | 40.84 | Eicosatetraenoic acid | - | C20H32O2 | 305.2496 | - |
| 122 | 41.38 | Octadecatrienoic acid | - | C18H30O2 | 279.2295 | - |
| 125 | 42.34 | Fatty acid derivative | - | C21H36O4 | 353.2694 | - |
| 129 | 43.34 | Pheophorbide A | 280, 410 | C35H36N4O5 | 593.2751 | 565, 533, 461 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barazetti, A.R.; Dealis, M.L.; Basso, K.R.; Silva, M.C.D.; Alves, L.d.C.; Parra, M.E.A.; Simionato, A.S.; Cely, M.V.T.; Macedo, A.L.; Silva, D.B.; et al. Evaluation of Resistance Induction Promoted by Bioactive Compounds of Pseudomonas aeruginosa LV Strain against Asian Soybean Rust. Microorganisms 2024, 12, 1576. https://doi.org/10.3390/microorganisms12081576
Barazetti AR, Dealis ML, Basso KR, Silva MCD, Alves LdC, Parra MEA, Simionato AS, Cely MVT, Macedo AL, Silva DB, et al. Evaluation of Resistance Induction Promoted by Bioactive Compounds of Pseudomonas aeruginosa LV Strain against Asian Soybean Rust. Microorganisms. 2024; 12(8):1576. https://doi.org/10.3390/microorganisms12081576
Chicago/Turabian StyleBarazetti, André Riedi, Mickely Liuti Dealis, Kawany Roque Basso, Maria Clara Davis Silva, Leonardo da Cruz Alves, Maria Eugênia Alcântara Parra, Ane Stéfano Simionato, Martha Viviana Torres Cely, Arthur Ladeira Macedo, Denise Brentan Silva, and et al. 2024. "Evaluation of Resistance Induction Promoted by Bioactive Compounds of Pseudomonas aeruginosa LV Strain against Asian Soybean Rust" Microorganisms 12, no. 8: 1576. https://doi.org/10.3390/microorganisms12081576