
Maize–Soybean Rotation and Intercropping Increase Maize Yield by Influencing the Structure and Function of Rhizosphere Soil Fungal Communities
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Sample Collection
2.4. DNA Extraction and Sequencing
2.5. Statistical Analysis
3. Results
3.1. Maize Plant Growth
3.2. Maize Yield and Yield Components
3.3. Rhizosphere Soil Fungal Community
3.3.1. Alpha Diversity of Maize Rhizosphere Soil Fungal Community
3.3.2. NMDS of Maize Rhizosphere Soil Fungal Community
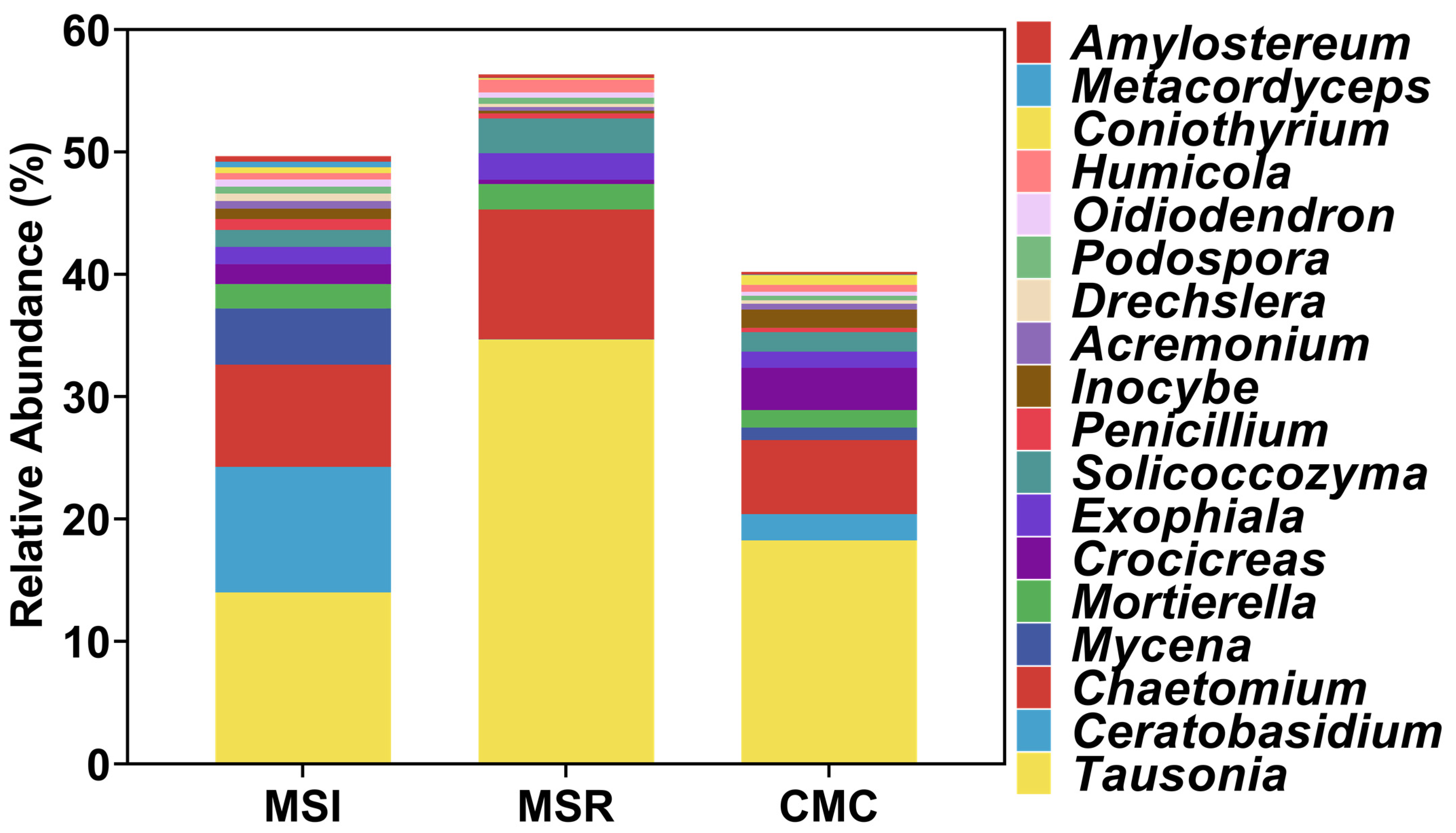
3.3.3. Fungal Community Composition in Maize Rhizosphere Soil under Different CropPing Systems
3.3.4. Co-Occurrence Network of the Maize Rhizosphere Soil Fungal Community
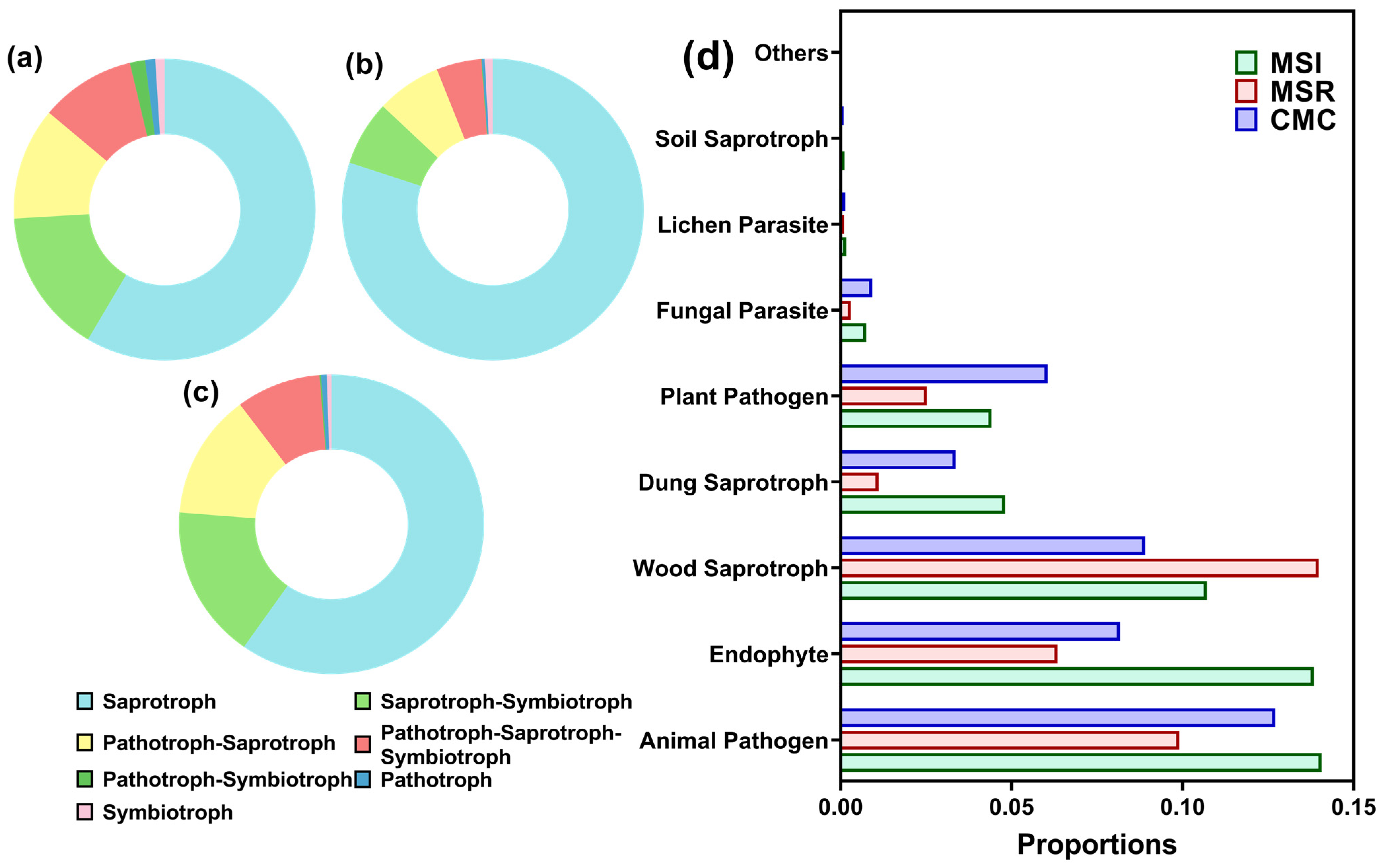
3.3.5. Prediction of Fungal Function in Rhizosphere Soil
3.3.6. Effects of Cropping System on Fungal Community Structure and Function and Maize Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiao, F.; Zhang, D.D.; Chen, Y.; Wu, J.H.; Zhang, J.Y. Effects of Long-Term Straw Returning and Nitrogen Fertilizer Reduction on Soil Microbial Diversity in Black Soil in Northeast China. Agronomy 2023, 13, 2036. [Google Scholar] [CrossRef]
- Li, R.P.; Zheng, J.Y.; Xie, R.Z.; Ming, B.; Peng, X.H.; Luo, Y.; Zheng, H.B.; Sui, P.X.; Wang, K.R.; Hou, P.; et al. Potential mechanisms of maize yield reduction under short-term no-tillage combined with residue coverage in the semi-humid region of Northeast China. Soil Tillage Res. 2022, 217, 10. [Google Scholar] [CrossRef]
- Du, G.M.; Yao, L.C.; Han, L.; Faye, B. What Should Be Learned from the Dynamic Evolution of Cropping Patterns in the Black Soil Region of Northeast China? A Case Study of Wangkui County, Heilongjiang Province. Land 2023, 12, 1574. [Google Scholar] [CrossRef]
- Zhang, H.Q. Effects of Soybean-Corn Rotation on Crop Yield, Economic Benefits, and Water Productivity in the Corn Belt of Northeast China. Sustainability 2023, 15, 1362. [Google Scholar] [CrossRef]
- Huang, Y.; Shang, M.Q.; Liu, T.T.; Wang, K.J. High-throughput methods for genome editing: The more the better. Plant Physiol. 2022, 188, 1731–1745. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.Y.; Ye, Q.X.; Huang, H.; Xia, L.L.; Tang, H.; Liu, X.Y.; Wu, J.W.; Li, Y.D.; Zhang, Y.Y.; Deng, L.J.; et al. Effects of grazing exclusion on soil microbial diversity and its functionality in grasslands: A meta-analysis. Front. Plant Sci. 2024, 15, 10. [Google Scholar] [CrossRef]
- Han, C.L.; Liang, D.F.; Zhou, W.D.; Xu, Q.Y.; Xiang, M.X.; Gu, Y.J.; Siddique, K.H.M. Soil, Plant, and Microorganism Interactions Drive Secondary Succession in Alpine Grassland Restoration. Plants 2024, 13, 1362. [Google Scholar] [CrossRef]
- Chen, D.M.; Huang, J.G.; Yuan, L. Positive effects of maize incorporation into the traditional tobacco cropping systems on soil nutrients, microbial biomass, enzyme activities, and bacterial compositions. Arch. Agron. Soil Sci. 2023, 69, 1674–1686. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, W.; Hou, K.X.; Zhang, J.W.; Du, Z.K.; Li, B.; Zhu, L.S. Ecological safety evaluation of chlorpyrifos on agricultural soil: Effects on soil microbes. Appl. Soil Ecol. 2023, 189, 11. [Google Scholar] [CrossRef]
- Zheng, B.C.; Chen, P.; Du, Q.; Yang, H.; Luo, K.; Wang, X.C.; Yang, F.; Yong, T.W.; Yang, W.Y. Soil Organic Matter, Aggregates, and Microbial Characteristics of Intercropping Soybean under Straw Incorporation and N Input. Agriculture 2022, 12, 1362. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, Y.X.; Li, T.; Chen, L.; Chen, Y.Q.; Sui, P. Changes in soil microbial biomass, diversity, and activity with crop rotation in cropping systems: A global synthesis. Appl. Soil Ecol. 2023, 186, 9. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, Y.D.; Zhang, K.; Jeong, J.; Zeng, Z.H.; Zang, H.D. Does crop rotation yield more in China? A meta-analysis. Field Crop. Res. 2020, 245, 107659. [Google Scholar] [CrossRef]
- Li, S.J.; Ye, S.; Liu, Z.Q.; Hassan, M.U.; Huang, G.Q.; Zhou, Q. How does intercropping contribute to soil organic carbon accumulation? A global synthesis. Agric. Ecosyst. Environ. 2024, 374, 10. [Google Scholar] [CrossRef]
- Xu, Z.; Li, C.J.; Zhang, C.C.; Yu, Y.; van der Werf, W.; Zhang, F.S. Intercropping maize and soybean increases efficiency of land and fertilizer nitrogen use; A meta-analysis. Field Crop. Res. 2020, 246, 107661. [Google Scholar] [CrossRef]
- Li, Q.M.; Zhang, D.; Zhang, J.Z.; Zhou, Z.J.; Pan, Y.; Yang, Z.H.; Zhu, J.H.; Liu, Y.H.; Zhang, L.F. Crop rotations increased soil ecosystem multifunctionality by improving keystone taxa and soil properties in potatoes. Front. Microbiol. 2023, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.J.; Li, S.; Baoyin, B.; Feng, Y.D.; Guo, D.Y.; Zhang, L.Q.; Gu, Y. Maize/Soybean Intercropping with Straw Return Increases Crop Yield by Influencing the Biological Characteristics of Soil. Microorganisms 2024, 12, 1108. [Google Scholar] [CrossRef] [PubMed]
- Faust, K. Open challenges for microbial network construction and analysis. Isme J. 2021, 15, 3111–3118. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Liu, Z.T.; Cheng, Z.R.; Wang, Q.; Hu, W.; Chen, B.L.; Meng, Y.L.; Zhou, Z.G. The trade-off between root growth redundancy and premature senescence under different straw returning modes affects boll formation and seedcotton yield. Eur. J. Agron. 2024, 156, 12. [Google Scholar] [CrossRef]
- Mwila, M.; Silva, J.V.; Kalala, K.; Simutowe, E.; Ngoma, H.; Nyagumbo, I.; Mataa, M.; Thierfelder, C. Do rotations and intercrops matter? Opportunities for intensification and diversification of maize-based cropping systems in Zambia. Field Crop. Res. 2024, 314, 13. [Google Scholar] [CrossRef]
- Tarekegne, A.; Wegary, D.; Cairns, J.E.; Zaman-Allah, M.; Beyene, Y.; Negera, D.; Teklewold, A.; Tesfaye, K.; Jumbo, M.B.; Das, B.; et al. Genetic gains in early maturing maize hybrids developed by the International Maize and Wheat Improvement Center in Southern Africa during 2000-2018. Front. Plant Sci. 2024, 14, 17. [Google Scholar] [CrossRef]
- Li, B.B.; Chen, X.M.; Deng, T.; Zhao, X.; Li, F.; Zhang, B.C.; Wang, X.; Shen, S.; Zhou, S.L. Timing effect of high temperature exposure on the plasticity of internode and plant architecture in maize. J. Integr. Agric. 2024, 23, 551–565. [Google Scholar] [CrossRef]
- Bian, D.H.; Jia, G.P.; Cai, L.J.; Ma, Z.Y.; Eneji, A.E.; Cui, Y.H. Effects of tillage practices on root characteristics and root lodging resistance of maize. Field Crop. Res. 2016, 185, 89–96. [Google Scholar] [CrossRef]
- Benitez, M.S.; Ewing, P.M.; Osborne, S.L.; Lehman, R.M. Rhizosphere microbial communities explain positive effects of diverse crop rotations on maize and soybean performance. Soil Biol. Biochem. 2021, 159, 13. [Google Scholar] [CrossRef]
- Liu, L.; Han, Q.L.; Liu, C.M.; Yang, J.; Liu, L.; Wu, J.R.; Li, C.Y. The Dynamics of Soil Microbe Metabolic Function Diversity in the Root-Zone of Maize-Soybean Intercropping. Int. J. Agric. Biol. 2019, 21, 639–647. [Google Scholar] [CrossRef]
- Nurgi, N.; Tana, T.; Dechassa, N.; Alemayehu, Y.; Tesso, B. Effects of planting density and variety on productivity of maize-faba bean intercropping system. Heliyon 2023, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Faghihinia, M.; Jansa, J.; Halverson, L.J.; Staddon, P.L. Hyphosphere microbiome of arbuscular mycorrhizal fungi: A realm of unknowns. Biol. Fertil. Soils 2023, 59, 17–34. [Google Scholar] [CrossRef]
- He, D.X.; Yao, X.D.; Zhang, P.Y.; Liu, W.B.; Huang, J.X.; Sun, H.X.; Wang, N.; Zhang, X.J.; Wang, H.Y.; Zhang, H.J.; et al. Effects of continuous cropping on fungal community diversity and soil metabolites in soybean roots. Microbiol. Spectr. 2023, 11, e01786-23. [Google Scholar] [CrossRef]
- Gahagan, A.C.; Shi, Y.C.; Radford, D.; Morrison, M.J.; Gregorich, E.; Aris-Brosou, S.; Chen, W. Long-Term Tillage and Crop Rotation Regimes Reshape Soil-Borne Oomycete Communities in Soybean, Corn, and Wheat Production Systems. Plants 2023, 12, 2338. [Google Scholar] [CrossRef]
- Wang, Q.F.; Zhou, D.P.; Chu, C.B.; Zhao, Z.; Ma, M.C.; Wu, S.H. The choice of rice rotation system affects the composition of the soil fungal community and functional traits. Heliyon 2024, 10, 12. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Majchrzak, L.; Borowiak, K.; Wolna-Maruwka, A.; Waraczewska, Z.; Budka, A.; Gaj, R. The Influence of Tillage and Cover Cropping on Soil Microbial Parameters and Spring Wheat Physiology. Agronomy 2020, 10, 200. [Google Scholar] [CrossRef]
- Shi, Y.X.; He, Y.S.; Zheng, Y.X.; Liu, X.X.; Wang, S.Z.; Xiong, T.E.; Wen, T.; Duan, H.; Liao, X.L.; Cui, Q.R.; et al. Characteristics of the phyllosphere microbial community and its relationship with major aroma precursors during the tobacco maturation process. Front. Plant Sci. 2024, 15, 14. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, A.Z.; Chen, X.W.; Zhang, S.X.; Zhang, Y.; McLaughlin, N.B.; Gao, Y.; Jia, S.X. The impact of cropping system, tillage and season on shaping soil fungal community in a long-term field trial. Eur. J. Soil Biol. 2021, 102, 10. [Google Scholar] [CrossRef]
- Nieves-Campos, E.I.; Méndez-Bravo, A.; Pérez-Bautista, Y.; Llanderal-Mendoza, J.; Guevara-Avendano, E.; Solís-García, I.A.; Diyarza-Sandoval, N.A.; Contreras-Ramos, S.M.; Rodriguez-Campos, J.; Méndez-Bravo, A.; et al. Anti-oomycete activity and plant growth promoting properties of avocado fungal endophytes. Rhizosphere 2024, 31, 10. [Google Scholar] [CrossRef]
- Peltoniemi, K.; Velmala, S.; Lloret, E.; Ollio, I.; Hyvönen, J.; Liski, E.; Brandt, K.K.; Campillo-Cora, C.; Fritze, H.; Iivonen, S.; et al. Soil and climatic characteristics and farming system shape fungal communities in European wheat fields. Agric. Ecosyst. Environ. 2024, 370, 12. [Google Scholar] [CrossRef]
- Nicola, L.; Landínez-Torres, A.Y.; Zambuto, F.; Capelli, E.; Tosi, S. The Mycobiota of High Altitude Pear Orchards Soil in Colombia. Biology-Basel 2021, 10, 1002. [Google Scholar] [CrossRef]
- Lu, Q.S.; Hu, C.L.; Cai, L.N.; Wu, C.F.; Zhang, H.Q.; Wei, L.; Zhang, T.Y.; Hu, H.C.; Liu, S.; Lei, J.J.; et al. Changes in soil fungal communities after onset of wheat yellow mosaic virus disease. Front. Bioeng. Biotechnol. 2022, 10, 13. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Amplification System | PCR Amplification Procedure | ||
|---|---|---|---|
| Reagent Composition | Volume | Reaction | |
| DNA template | 30 ng | ① Pre-denaturation | 94 °C 2 min |
| Forward Primer (5 uM) | 1 μL | ② Denaturation | 94 °C 15 s |
| Reverse Primer (5 uM) | 1 μL | ③ Anneal | 55 °C 30 s |
| BSA (2 ng μL−1) | 3 μL | ④ Extension | 72 °C 60 s |
| 2x Taq Plus Master Mix | 12.5 μL | ⑤ Final extension | 72 °C 7 min |
| ddH2O | 7.5 μL | ③–⑤ Number of cycles | 30 |
| Total | 25 μL | ⑥ Reaction termination | 10 °C ∞ |
| Step | Instrument/Reagents | Manufacturer | Specification/Model/Lot Number |
|---|---|---|---|
| Amplicon extraction | MoBio PowerSoil DNA Isolation Kit (100) | QIAGEN (Frankfurt, Germany) | 100 times |
| Amplifier amplification | KAPA 2G Robust Hot Start Ready Mix | KAPA (Boston, MA, USA) | |
| ABI 9700 PCR | ABI (Guangzhou, China) | ||
| Amplicon purification | Agencourt® AMPure® XP | Beckman Coulter (Shanghai, China) | Dispense 45 mL/bottle, total 450 mL/bottle |
| Amplicon building | NEBNext Ultra II DNA Library Prep Kit | NEB (Beijing, China) | 96 reactions |
| Agencourt® AMPure® XP | Beckman Coulter | Dispense 45 mL/bottle, total 450 mL/bottle | |
| ABI 9700 PCR | ABI | ||
| Library quality control (instruments) | Bioanalyzer (Agilent 2100) | Agilent (Palo Alto, CA, USA) | DE13806339 |
| Biomolecule Analyzer (Labchip GX) | PerkinElmer (Shanghai, China) | ||
| ABI Qpcr | ABI | Step One Plus (Shanghai, China) (www.PuDi.cn) (accessed on 1 August 2024) | |
| Library quality control (reagents) | Agilent DNA 1000 Kit | Agilent (Palo Alto, CA, USA) | 300 samples |
| HT DNA-Extended Range LabChip | PerkinElmer (Shanghai, China) | ||
| KAPA Library Quantification Kit | KAPA | 500 times | |
| Sequencing (equipment) | High-throughput second-generation sequencer | Illumina (Beijing, China) | MiSeq (Shanghai, China) (www.PuDi.cn) (accessed on 1 August 2024) |
| Sequencing (reagents) | MiSeq® Reagent Kit v3 (600 cycle) (PE300) | Illumina | |
| MiSeq Reagent Kit v2 (500 cycle) | Illumina |
| Treatment | Node | Edge | Positive (%) | Negative (%) | Average Degree | Average Weighting | Modularity | Cluster Coefficient |
|---|---|---|---|---|---|---|---|---|
| MSI | 201 | 1114 | 57.95 | 42.05 | 11.14 | 10.36 | 0.68 | 0.65 |
| MSR | 199 | 1266 | 47.32 | 52.68 | 12.74 | 11.77 | 0.65 | 0.62 |
| CMC | 200 | 1309 | 59.11 | 40.89 | 13.03 | 12.04 | 0.63 | 0.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Yang, Y.; Zhao, Z.; Feng, Y.; Bate, B.; Wang, H.; Li, Q.; Cui, J. Maize–Soybean Rotation and Intercropping Increase Maize Yield by Influencing the Structure and Function of Rhizosphere Soil Fungal Communities. Microorganisms 2024, 12, 1620. https://doi.org/10.3390/microorganisms12081620
Zhang L, Yang Y, Zhao Z, Feng Y, Bate B, Wang H, Li Q, Cui J. Maize–Soybean Rotation and Intercropping Increase Maize Yield by Influencing the Structure and Function of Rhizosphere Soil Fungal Communities. Microorganisms. 2024; 12(8):1620. https://doi.org/10.3390/microorganisms12081620
Chicago/Turabian StyleZhang, Liqiang, Yuhan Yang, Zehang Zhao, Yudi Feng, Baoyin Bate, Hongyu Wang, Qiuzhu Li, and Jinhu Cui. 2024. "Maize–Soybean Rotation and Intercropping Increase Maize Yield by Influencing the Structure and Function of Rhizosphere Soil Fungal Communities" Microorganisms 12, no. 8: 1620. https://doi.org/10.3390/microorganisms12081620