
Akkermansia muciniphila Metabolite Inosine Inhibits Castration Resistance in Prostate Cancer
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Bacterial Culture
2.3. Mice
2.4. Human Study
2.5. Fecal Microbiota Transplantation Experiment
2.6. FITC-Dextran Permeability Experiment
- In vivo
- In vitro
2.7. Trans-Epithelial Electrical Resistance (TEER)
2.8. ELISA
2.9. Gene Expression Analysis
2.10. Protein Expression Analysis
2.11. Histological Staining and Analysis
2.12. 16. S rDNA Analysis
2.13. Untargeted Metabolomic Analysis
2.14. Statistical Analysis
3. Results
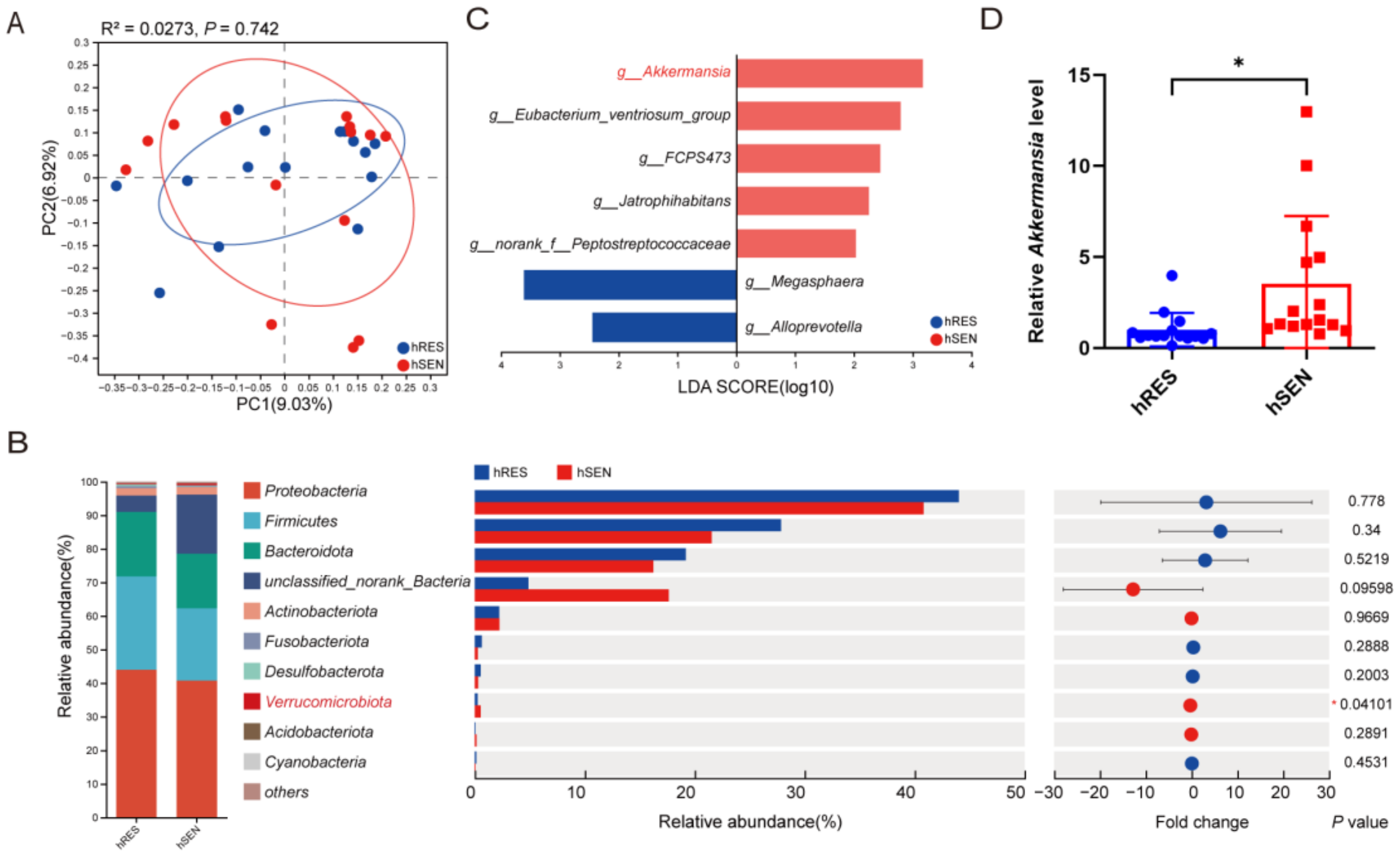
3.1. Differences in Gut Microbiota among hRES and hSEN PCa Patients
3.2. Gut Microbiota Influenced the Resistance to ADT against PCa
3.3. AKK Was Associated with Treatment Sensitivity to ADT
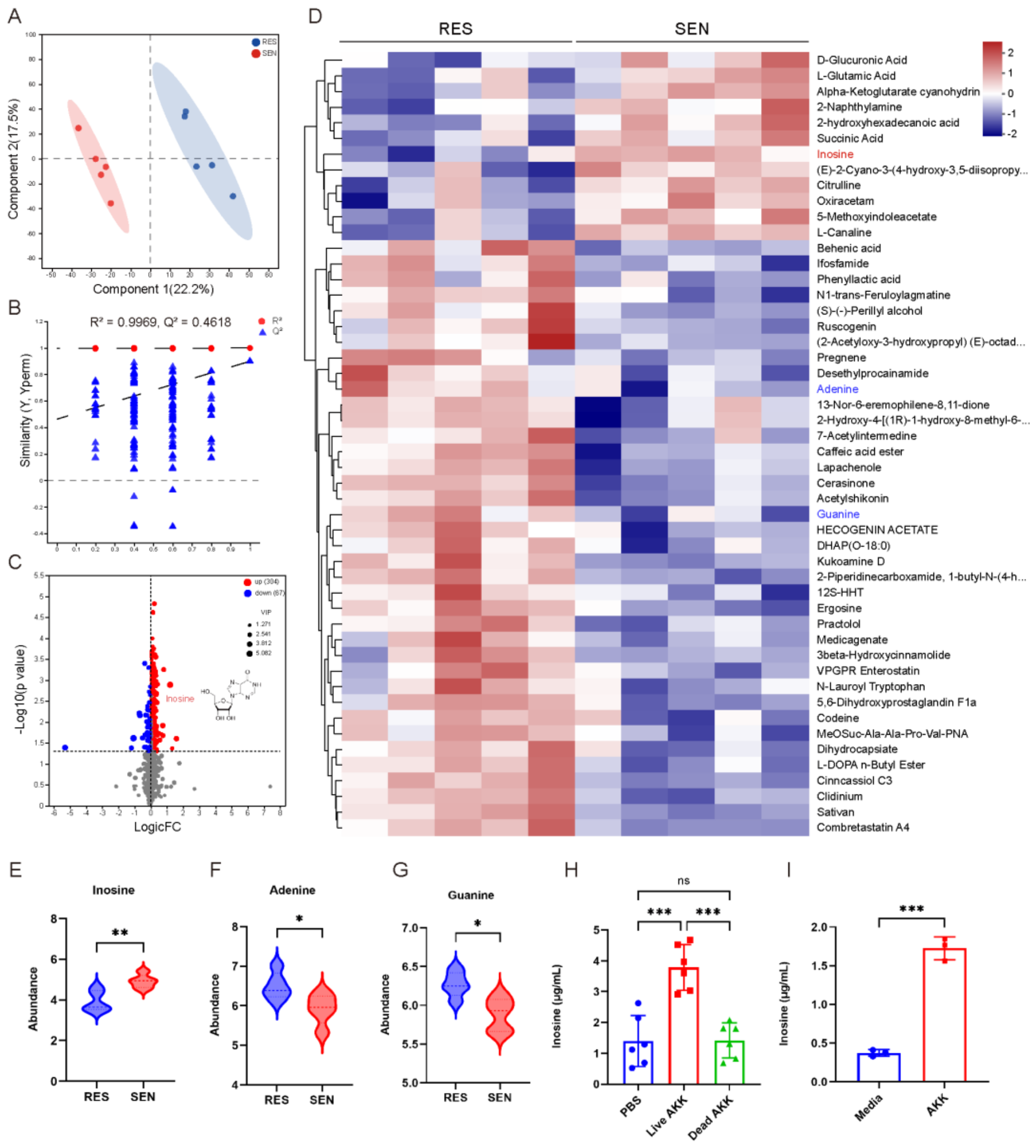
3.4. The Elevated Inosine Synthesis in SEN Group Was Associated with AKK
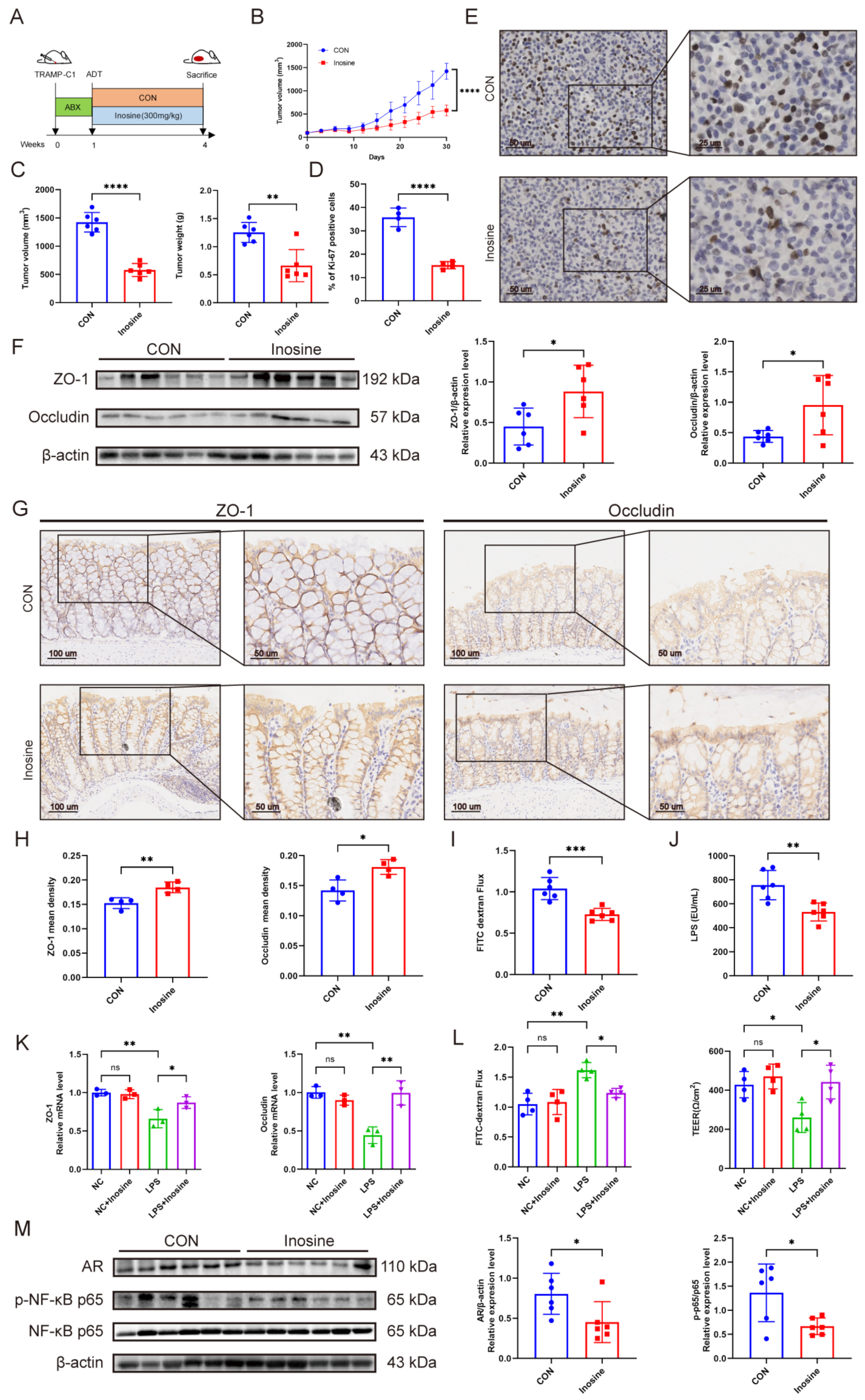
3.5. Intestinal Barrier Dysfunction and Elevated LPS Promoted Castration Resistance
3.6. The Metabolite Inosine Alleviated Intestinal Barrier Damage and Inhibited Castration Resistance
3.7. Predictive Model for CRPC Based on Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Burden of Disease 2019 Cancer Collaboration; Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life Years for 2022, 29 Cancer Groups From 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [PubMed]
- Rebello, R.J.; Oing, C.; Knudsen, K.E.; Loeb, S.; Johnson, D.C.; Reiter, R.E.; Gillessen, S.; Kwast, T.V.d.; Bristow, R.G. Prostate cancer. Nat. Rev. Dis. Primers 2021, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Desai, K.; McManus, J.M.; Sharifi, N. Hormonal Therapy for Prostate Cancer. Endocr. Rev. 2021, 42, 354–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, N.; Liu, Q.; Guo, J.; Pan, Q.; Cheng, B.; Xu, J.; Dong, B.; Yang, G.; Yang, B.; et al. Antiandrogen treatment induces stromal cell reprogramming to promote castration resistance in prostate cancer. Cancer Cell 2023, 41, 1345–1362.e9. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Song, X.-L.; Li, X.-A.; Chen, M.; Guo, J.; Yang, D.-H.; Chen, Z.; Zhao, S.-C. Current therapy and drug resistance in metastatic castration-resistant prostate cancer. Drug Resist. Updat. 2023, 68, 100962. [Google Scholar] [CrossRef] [PubMed]
- Pernigoni, N.; Zagato, E.; Calcinotto, A.; Troiani, M.; Mestre, R.P.; Calì, B.; Attanasio, G.; Troisi, J.; Minini, M.; Mosole, S.; et al. Commensal bacteria promote endocrine resistance in prostate cancer through androgen biosynthesis. Science 2021, 374, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Wu, K.; Long, Z.; Zhou, X.; Zhong, C.; Wang, S.; Lai, H.; Guo, Y.; Lv, D.; Lu, J.; et al. Gut dysbiosis promotes prostate cancer progression and docetaxel resistance via activating NF-κB-IL6-STAT3 axis. Microbiome 2022, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Fujita, K.; Motooka, D.; Hatano, K.; Fukae, S.; Kawamura, N.; Tomiyama, E.; Hayashi, Y.; Banno, E.; Takao, T.; et al. The gut microbiota associated with high-Gleason prostate cancer. Cancer Sci. 2021, 112, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.R.; Ha, G.; Hoff, A.M.; Wala, J.A.; Carrot-Zhang, J.; Whelan, C.W.; Haradhvala, N.J.; Freeman, S.S.; Reed, S.C.; Rhoades, J.; et al. Structural alterations driving castration-resistant prostate cancer revealed by linked-read genome sequencing. Cell 2018, 174, 433–447.e19. [Google Scholar] [CrossRef]
- Takeda, D.Y.; Spisák, S.; Seo, J.-H.; Bell, C.; O’Connor, E.; Korthauer, K.; Ribli, D.; Csabai, I.; Solymosi, N.; Szállási, Z.; et al. A somatically acquired enhancer of the androgen receptor is a noncoding driver in advanced prostate cancer. Cell 2018, 174, 422–432.e13. [Google Scholar] [CrossRef]
- Joseph, J.D.; Lu, N.; Qian, J.; Sensintaffar, J.; Shao, G.; Brigham, D.; Moon, M.; Maneval, E.C.; Chen, I.; Darimont, B.; et al. A Clinically Relevant Androgen Receptor Mutation Confers Resistance to Second-Generation Antiandrogens Enzalutamide and ARN-509. Cancer Discov. 2013, 3, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, E.S.; Lu, C.; Wang, H.; Luber, B.; Nakazawa, M.; Roeser, J.C.; Chen, Y.; Mohammad, T.A.; Chen, Y.; Fedor, H.L.; et al. AR-V7 and Resistance to Enzalutamide and Abiraterone in Prostate Cancer. N. Engl. J. Med. 2014, 371, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Cato, L.; de Tribolet-Hardy, J.; Lee, I.; Rottenberg, J.T.; Coleman, I.; Melchers, D.; Houtman, R.; Xiao, T.; Li, W.; Uo, T.; et al. ARv7 Represses Tumor-Suppressor Genes in Castration-Resistant Prostate Cancer. Cancer Cell 2019, 35, 401–413.e6. [Google Scholar] [CrossRef] [PubMed]
- Hakozaki, Y.; Yamada, Y.; Kawai, T.; Nakamura, M.; Takeshima, Y.; Iwaki, T.; Kinoshita, Y.; Fujii, Y.; Akiyama, Y.; Sato, Y. Time to castration resistance is a novel prognostic factor of cancer-specific survival in patients with nonmetastatic castration-resistant prostate cancer. Sci. Rep. 2022, 12, 16202. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Li, J.; Lyu, F.; Xiong, Q.; Gu, P.; Chen, Y.; Chen, M.; Bao, J.; Zhang, X.; Wei, R.; et al. Novel tripeptide RKH derived from Akkermansia muciniphila protects against lethal sepsis. Gut 2024, 73, 78–91. [Google Scholar] [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Hupp, T.; Duchnowska, R.; Marek-Trzonkowska, N.; Połom, K. Next-generation probiotics—Do they open new therapeutic strategies for cancer patients? Gut Microbes 2022, 14, 2035659. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, J.; Li, D.; Yang, H.; Chen, C.; Qin, M.; Wen, Z.; He, Z.; Xu, L. Akkermansia muciniphila: A potential target and pending issues for oncotherapy. Pharmacol. Res. 2023, 196, 106916. [Google Scholar] [CrossRef]
- Cross, T.-W.L.; Simpson, A.M.R.; Lin, C.-Y.; Hottmann, N.M.; Bhatt, A.P.; Pellock, S.J.; Nelson, E.R.; Loman, B.R.; Wallig, M.A.; Vivas, E.I.; et al. Gut microbiome responds to alteration in female sex hormone status and exacerbates metabolic dysfunction. Gut Microbes 2024, 16, 2295429. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, D.; Robinson, A.; Lagomarsino, V.N.; Matthews, L.T.; Hassan, R.; Perez, K.; Dan, W.; Yim, P.D.; Mixer, M.; Prochera, A.; et al. Diminished androgen levels are linked to irritable bowel syndrome and cause bowel dysfunction in mice. J. Clin. Investig. 2022, 132, e150789. [Google Scholar] [CrossRef]
- Scher, H.I.; Halabi, S.; Tannock, I.; Morris, M.; Sternberg, C.N.; Carducci, M.A.; Eisenberger, M.A.; Higano, C.; Bubley, G.J.; Dreicer, R.; et al. Design and end points of clinical trials for patients with progressive prostate cancer and castrate levels of testosterone: Recommendations of the Prostate Cancer Clinical Trials Working Group. J. Clin. Oncol. 2008, 26, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Bjartell, A.; Agarwal, N.; Chung, B.H.; Given, R.W.; Pereira de Santana Gomes, A.P.; Merseburger, A.S.; Özgüroğlu, M.; Soto, Á.J.; Uemura, H.; et al. Deep, rapid, and durable prostate-specific antigen decline with apalutamide plus androgen deprivation therapy is associated with longer survival and improved clinical outcomes in TITAN patients with metastatic castration-sensitive prostate cancer. Ann. Oncol. 2023, 34, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Xie, W.; D’Amico, A.V.; Ross, R.W.; Hu, J.C.; Pomerantz, M.; Regan, M.M.; Taplin, M.E.; Kantoff, P.W.; Sartor, O.; et al. Time to prostate-specific antigen nadir independently predicts overall survival in patients who have metastatic hormone-sensitive prostate cancer treated with androgen-deprivation therapy. Cancer 2009, 115, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, Y.; Wan, M.; Mei, F.; Wang, F.; Gu, P.; Zhang, X.; Wei, R.; Zeng, Y.; Zheng, H.; et al. Oral magnesium prevents acetaminophen-induced acute liver injury by modulating microbial metabolism. Cell Host Microbe 2024, 32, 48–62.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Qiu, Y.; Li, K.; Sun, Q.; Xie, M.; Huang, P.; Yu, Y.; Wang, B.; Xue, J.; Zhu, Z.; et al. Unraveling the impact of Lactobacillus spp. and other urinary microorganisms on the efficacy of mirabegron in female patients with overactive bladder. Front. Cell. Infect. Microbiol. 2022, 12, 1030315. [Google Scholar] [CrossRef]
- Qiu, Y.; Gao, Y.; Chen, C.; Xie, M.; Huang, P.; Sun, Q.; Zhou, Z.; Li, B.; Zhao, J.; Wu, P. Deciphering the influence of urinary microbiota on FoxP3+ regulatory T cell infiltration and prognosis in Chinese patients with non-muscle-invasive bladder cancer. Hum. Cell 2022, 35, 511–521. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Cassilly, C.D.; Liu, X.; Park, S.-M.; Tusi, B.K.; Chen, X.; Kwon, J.; Filipčík, P.; Bolze, A.S.; Liu, Z.; et al. Akkermansia muciniphila phospholipid induces homeostatic immune responses. Nature 2022, 608, 168–173. [Google Scholar] [CrossRef]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum polysaccharide modulates gut microbiota and immune cell function to inhibit inflammation and tumorigenesis in colon. Carbohydr. Polym. 2021, 267, 118231. [Google Scholar] [CrossRef]
- Yang, L.-Y.; Luo, Q.; Lu, L.; Zhu, W.-W.; Sun, H.-T.; Wei, R.; Lin, Z.F.; Wang, X.Y.; Wang, C.Q.; Lu, M.; et al. Increased neutrophil extracellular traps promote metastasis potential of hepatocellular carcinoma via provoking tumorous inflammatory response. J. Hematol. Oncol. 2020, 13, 3. [Google Scholar] [CrossRef]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef]
- Li, D.; Liu, R.; Wang, M.; Peng, R.; Fu, S.; Fu, A.; Le, J.; Yao, Q.; Yuan, T.; Chi, H.; et al. 3β-Hydroxysteroid dehydrogenase expressed by gut microbes degrades testosterone and is linked to depression in males. Cell Host Microbe 2022, 30, 329–339.e5. [Google Scholar] [CrossRef] [PubMed]
- Markle, J.G.M.; Frank, D.N.; Mortin-Toth, S.; Robertson, C.E.; Feazel, L.M.; Rolle-Kampczyk, U.; Von Bergen, M.; McCoy, K.D.; Macpherson, A.J.; Danska, J.S. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 2013, 339, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.-H.; Chou, C.-H.; Chen, Y.-L.; Wang, P.-H.; Brandon-Mong, G.-J.; Lee, T.-H.; Wu, T.-Y.; Li, P.-T.; Li, C.-W.; Lai, Y.-L.; et al. Circulating androgen regulation by androgen-catabolizing gut bacteria in male mouse gut. Gut Microbes 2023, 15, 2183685. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Yang, X.; Zhou, M.; Feng, L.; Ji, C.; Liang, J.; Zhang, B.; Shen, R.; Wang, L. Inhibition of inosine metabolism of the gut microbiota decreases testosterone secretion in the testis. mSystems 2024, 9, e0013824. [Google Scholar] [CrossRef] [PubMed]
- Busada, J.T.; Peterson, K.N.; Khadka, S.; Xu, X.; Oakley, R.H.; Cook, D.N.; Cidlowski, J.A. Glucocorticoids and androgens protect from gastric metaplasia by suppressing group 2021, 2 innate lymphoid cell activation. Gastroenterology 2021, 161, 637–652.e4. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, S.; Xu, Y.; Zhang, Y.; Ma, W.; Liang, C.; Chen, D.; Li, J. Androgen Maintains Intestinal Homeostasis by Inhibiting BMP Signaling via Intestinal Stromal Cells. Stem Cell Rep. 2020, 15, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. NF-kappaB as a critical link between inflammation and cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a000141. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin Is Associated With Higher Relative Abundance of Mucin-Degrading Akkermansia muciniphila and Several Short-Chain Fatty Acid-Producing Microbiota in the Gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef]
- He, K.-Y.; Lei, X.-Y.; Wu, D.-H.; Zhang, L.; Li, J.-Q.; Li, Q.-T.; Yin, W.-T.; Zhao, Z.-L.; Liu, H.; Xiang, X.-Y.; et al. Akkermansia muciniphila protects the intestine from irradiation-induced injury by secretion of propionic acid. Gut Microbes 2023, 15, 2293312. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.; Li, J.; Hao, G.; An, X.; Song, Y.; Wei, H.; Ge, W. Stachyose increases intestinal barrier through Akkermansia muciniphila and reduces gut inflammation in germ-free mice after human fecal transplantation. Food Res. Int. 2020, 137, 109288. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, X.; Guan, H.; Chen, S.; Fan, X.; Zhou, C.; Yang, H.; Xiao, W. A Novel Role of A2AR in the Maintenance of Intestinal Barrier Function of Enteric Glia from Hypoxia-Induced Injury by Combining with mGluR5. Front. Pharmacol. 2021, 12, 633403. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Pan, Y.; Guo, Y.; Zhu, Y.; Jin, H.; Gu, Y.; Li, C.; Wang, Y.; Lin, J.; Chen, Y.; et al. Symbiotic combination of Akkermansia muciniphila and inosine alleviates alcohol-induced liver injury by modulating gut dysbiosis and immune responses. Front. Microbiol. 2024, 15, 1355225. [Google Scholar] [CrossRef] [PubMed]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Yan, S.; Chang, J.; Hao, X.; Liu, J.; Tan, X.; Geng, Z.; Wang, Z. Berberine regulates short-chain fatty acid metabolism and alleviates the colitis-associated colorectal tumorigenesis through remodeling intestinal flora. Phytomedicine 2022, 102, 154217. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kuang, J.; Zhang, H.; Chen, W.; Zheng, X.; Wang, J.; Huang, F.; Ge, K.; Li, M.; Zhao, M.; et al. Bile Acid-Microbiome Interaction Promotes Gastric Carcinogenesis. Adv. Sci. 2022, 9, e2200263. [Google Scholar] [CrossRef] [PubMed]
- Gannon, P.O.; Lessard, L.; Stevens, L.-M.; Forest, V.; Bégin, L.R.; Minner, S.; Tennstedt, P.; Schlomm, T.; Mes-Masson, A.M.; Saad, F. Large-scale independent validation of the nuclear factor-kappa B p65 prognostic biomarker in prostate cancer. Eur. J. Cancer 2013, 49, 2441–2448. [Google Scholar] [CrossRef]
- McCall, P.; Bennett, L.; Ahmad, I.; MacKenzie, L.M.; Forbes, I.W.G.; Leung, H.Y.; Sansom, O.J.; Orange, C.; Seywright, M.; Underwood, M.A.; et al. NFκB signalling is upregulated in a subset of castrate-resistant prostate cancer patients and correlates with disease progression. Br. J. Cancer 2012, 107, 1554–1563. [Google Scholar] [CrossRef]
- Thomas-Jardin, S.E.; Dahl, H.; Nawas, A.F.; Bautista, M.; Delk, N.A. NF-κB signaling promotes castration-resistant prostate cancer initiation and progression. Pharmacol. Ther. 2020, 211, 107538. [Google Scholar] [CrossRef]
- Jin, R.; Yi, Y.; Yull, F.E.; Blackwell, T.S.; Clark, P.E.; Koyama, T.; Smith Jr, J.A.; Matusik, R.J. NF-κB gene signature predicts prostate cancer progression. Cancer Res. 2014, 74, 2763–2772. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Zaslavsky, A.; Fedele, G.; McLaughlin, S.K.; Reczek, E.E.; De Raedt, T.; Guney, I.; Strochlic, D.E.; MacConaill, L.E.; Beroukhim, R.; et al. An oncogene-tumor suppressor cascade drives metastatic prostate cancer by coordinately activating Ras and nuclear factor-kappaB. Nat. Med. 2010, 16, 286–294. [Google Scholar] [CrossRef]
- D’Amico, A.V.; Chen, M.-H.; Roehl, K.A.; Catalona, W.J. Preoperative PSA velocity and the risk of death from prostate cancer after radical prostatectomy. N. Engl. J. Med. 2004, 351, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Hamano, I.; Hatakeyama, S.; Narita, S.; Takahashi, M.; Sakurai, T.; Kawamura, S.; Hoshi, S.; Ishida, M.; Kawaguchi, T.; Ishidoya, S.; et al. Impact of nadir PSA level and time to nadir during initial androgen deprivation therapy on prognosis in patients with metastatic castration-resistant prostate cancer. World J. Urol. 2019, 37, 2365–2373. [Google Scholar] [CrossRef]
- Huang, S.-P.; Bao, B.-Y.; Wu, M.-T.; Choueiri, T.K.; Goggins, W.B.; Huang, C.-Y.; Pu, Y.-S.; Yu, C.-C.; Huang, C.-H. Impact of prostate-specific antigen (PSA) nadir and time to PSA nadir on disease progression in prostate cancer treated with androgen-deprivation therapy. Prostate 2011, 71, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Sternberg, C.N.; Efstathiou, E.; Fizazi, K.; Shen, Q.; Lin, X.; Sugg, J.; Steinberg, J.; Noerby, B.; De Giorgi, U.; et al. Nadir Prostate-specific Antigen as an Independent Predictor of Survival Outcomes: A Post Hoc Analysis of the PROSPER Randomized Clinical Trial. J. Urol. 2023, 209, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Derosa, L.; Routy, B.; Thomas, A.M.; Iebba, V.; Zalcman, G.; Friard, S.; Mazieres, J.; Audigier-Valette, C.; Moro-Sibilot, D.; Goldwasser, F.; et al. Intestinal Akkermansia muciniphila predicts clinical response to PD-1 blockade in patients with advanced non-small-cell lung cancer. Nat. Med. 2022, 28, 315–324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Data | Number (%), Median (IQR) | p Value | ||
|---|---|---|---|---|
| Total | hRES Group | hSEN Group | ||
| Number of patients | 30 | 15 | 15 | |
| PFS (months) | 24.5 (20–25.75) | 20 (17.5–22) | 26 (25–27.5) | <0.001 |
| Age (years) | 70 (65–74) | 72 (68–78.5) | 69 (63–72) | 0.033 |
| BMI (kg/m2) | 23.47 (23.03–24.59) | 23.25 (22.67–23.62) | 24.46 (23.27–25.42) | 0.026 |
| TNM | ||||
| T1 + 2 | 17 (56.7%) | 5 | 12 | 0.0253 |
| T3 + 4 | 13 (43.3%) | 10 | 3 | |
| N0 | 22 (73.3%) | 10 | 12 | 0.6817 |
| N1 | 8 (26.7%) | 5 | 3 | |
| M0 | 18 (60%) | 10 | 8 | 0.7104 |
| M1 | 12 (40%) | 5 | 7 | |
| Gleason score | 0.0996 | |||
| G1 (3 + 3) | 4 (13.3%) | 1 | 3 | |
| G2 (3 + 4) | 5 (16.7%) | 2 | 3 | |
| G3 (4 + 3) | 2 (6.7) | 2 | 0 | |
| G4 (8) | 6 (20%) | 1 | 5 | |
| G5 (9,10) | 13 (43.3%) | 9 | 4 | |
| PSA diagnosis (ng/mL) | 15.66 (11.26–38.83) | 17.11 (11.49–40.95) | 14.8 (11.38–29.34) | 0.339 |
| nPSA (ng/mL) | 0.27 (0.13–1.21) | 1.23 (0.549–2.15) | 0.13 (0.08–0.16) | <0.001 |
| Variable | PFS | |
|---|---|---|
| HR (95% CI) | p Value | |
| Age (years) | 1.1 (1–1.2) | 0.05 |
| BMI (kg/m2) | 0.77 (0.57–1) | 0.087 |
| TNM | 2.5 (1.3–4.8) | 0.0068 |
| Gleason | 1.3 (0.87–1.9) | 0.22 |
| PSA diagnose (ng/mL) | 1 (1–1) | 0.82 |
| nPSA (ng/mL) | 3 (1.8–5) | 0.00003 |
| AKK abundance (%) | 0.011 (0.00013–0.91) | 0.046 |
| Variable | PFS | |
|---|---|---|
| HR (95% CI) | p Value | |
| Age (years) | 1.04 (0.96–1.13) | 0.319 |
| TNM | 1.89 (0.88–4.00) | 0.102 |
| nPSA (ng/mL) | 2.39 (1.37–4.15) | 0.002 |
| AKK abundance (%) | 0.30 (0.02–3.86) | 0.355 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Li, L.; Yang, Q.; Xue, J.; Wang, B.; Xie, M.; Shangguan, W.; Zhu, Z.; Wu, P. Akkermansia muciniphila Metabolite Inosine Inhibits Castration Resistance in Prostate Cancer. Microorganisms 2024, 12, 1653. https://doi.org/10.3390/microorganisms12081653
Yu Y, Li L, Yang Q, Xue J, Wang B, Xie M, Shangguan W, Zhu Z, Wu P. Akkermansia muciniphila Metabolite Inosine Inhibits Castration Resistance in Prostate Cancer. Microorganisms. 2024; 12(8):1653. https://doi.org/10.3390/microorganisms12081653
Chicago/Turabian StyleYu, Yao, Leqian Li, Qishen Yang, Jingwen Xue, Benlin Wang, Ming Xie, Wentai Shangguan, Zhangrui Zhu, and Peng Wu. 2024. "Akkermansia muciniphila Metabolite Inosine Inhibits Castration Resistance in Prostate Cancer" Microorganisms 12, no. 8: 1653. https://doi.org/10.3390/microorganisms12081653