
Review of the Proteomics and Metabolic Properties of Corynebacterium glutamicum


Abstract
:1. Introduction
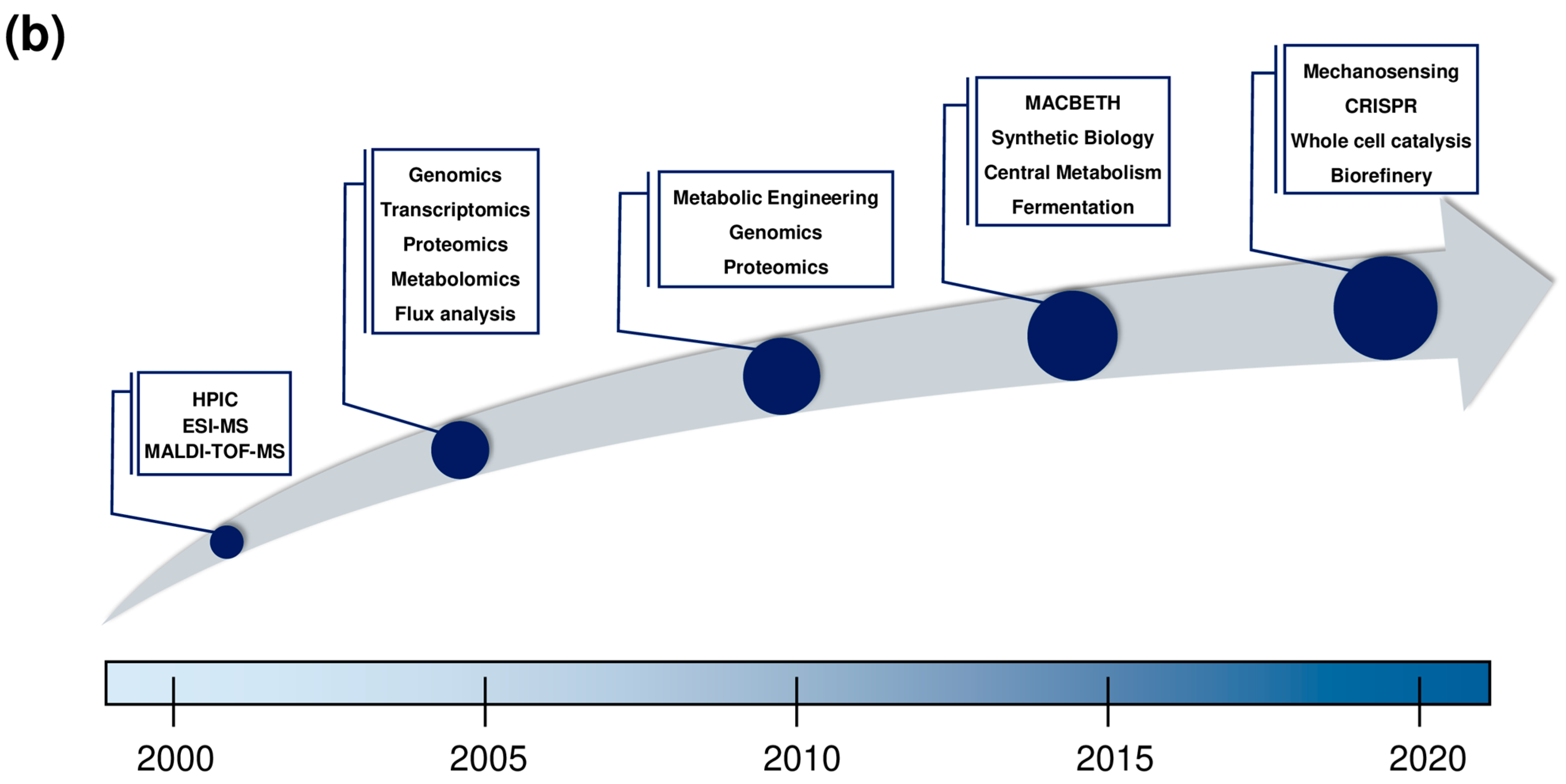
2. Corynebacterium glutamicum Research Timeline and Overview
3. Importance of Proteome Research and Its Impact on Future Research
3.1. Use of C. glutamicum for the Mass Production of Useful Proteins
3.2. Use of C. glutamicum as a Biosensor and Diagnostic Tool
3.3. Biological Purification
3.4. Association between Central Carbon Metabolism and Proteomics
4. Central Carbon Metabolism Research and Carbon Source Characteristics
4.1. Metabolic Characteristics According to Carbon Source Type
4.2. Metabolite Production Based on Carbon Sources
5. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinoshita, S.; Udaka, S.; Shimono, M. Studies on the Amino Acid Fermentation. J. Gen. Appl. Microbiol. 1957, 3, 193–205. [Google Scholar] [CrossRef]
- Kinoshita, S.; Nakayama, K.; Akita, S. Taxonomical study of glutamic acid accumulating bacteria, Micrococcus glutamicus nov. sp. J. Agric. Chem. Soc. Jpn. 1958, 22, 176–185. [Google Scholar] [CrossRef]
- Abe, S.; Takayama, K.-I.; Kinoshita, S. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Li, T.-H. The Production of Glutamic Acid by Fermentation. Master’s Thesis, University of Missouri at Rolla, Rolla, MO, USA, 1965. [Google Scholar]
- Hermann, T. Industrial production of amino acids by coryneform bacteria. J. Biotechnol. 2003, 104, 155–172. [Google Scholar] [CrossRef]
- Siewe, R.M.; Weil, B.; Burkovski, A.; Eikmanns, B.J.; Eikmanns, M.; Kramer, R. Functional and genetic characterization of the (methyl)ammonium uptake carrier of Corynebacterium glutamicum. J. Biol. Chem. 1996, 271, 5398–5403. [Google Scholar] [CrossRef]
- Hermann, T.; Wersch, G.; Uhlemann, E.-M.; Schmid, R.; Burkovski, A. Mapping and identification of Corynebacterium glutamicum proteins by two-dimensional gel electrophoresis and microsequencing. Electrophoresis 1998, 19, 3217–3221. [Google Scholar] [CrossRef] [PubMed]
- Peter, H.; Burkovski, A.; Kramer, R. Osmo-sensing by N- and C-terminal extensions of the glycine betaine uptake system BetP of Corynebacterium glutamicum. J Biol Chem 1998, 273, 2567–2574. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.; de Graaf, A.A.; Eggeling, L.; Mollney, M.; Wiechert, W.; Sahm, H. In vivo quantification of parallel and bidirectional fluxes in the anaplerosis of Corynebacterium glutamicum. J. Biol. Chem. 2000, 275, 35932–35941. [Google Scholar] [CrossRef]
- Rübenhagen, R.; Rönsch, H.; Jung, H.; Krämer, R.; Morbach, S. Osmosensor and osmoregulator properties of the betaine carrier BetP from Corynebacterium glutamicum in proteoliposomes. J. Biol. Chem. 2000, 275, 735–741. [Google Scholar] [CrossRef]
- Nakayama, K. Development of L-Lysine Production Technologies. J. Synth. Org. Chem. 1977, 35, 64–72. [Google Scholar] [CrossRef]
- Nakayama, K.; Araki, K.; Kase, H. Microbial Production of Essential Amino Acids with Corynebacterium Glutamicum Mutants. In Nutritional Improvement of Food and Feed Proteins; Friedman, M., Ed.; Springer: Boston, MA, USA, 1978; pp. 649–661. [Google Scholar]
- Follettie, M.; Shin, H.; Sinskey, A. Organization and regulation of the Corynebacterium glutamicum hom-thrB and thrC loci. Mol. Microbiol. 1988, 2, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Louw, M.E. Studies on the Improvement of Lysine Production in the Genera Brevibacterium and Corynebacterium. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 1983. [Google Scholar]
- Ebbighausen, H.; Weil, B.; Krämer, R. Isoleucine excretion in Corynebacterium glutamicum: Evidence for a specific efflux carrier system. Appl. Microbiol. Biotechnol. 1989, 31, 184–190. [Google Scholar] [CrossRef]
- Luntz, M.G.; Zhdanova, N.I.; Bourd, G.I. Transport and excretion of L-lysine in Corynebacterium glutamicum. Microbiology 1986, 132, 2137–2146. [Google Scholar] [CrossRef]
- Lee, J.H.; Wendisch, V.F. Production of amino acids—Genetic and metabolic engineering approaches. Bioresour. Technol. 2017, 245, 1575–1587. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. Systems and synthetic metabolic engineering for amino acid production—The heartbeat of industrial strain development. Curr. Opin. Biotechnol. 2012, 23, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Kitagawa, K.; Jo, S.J.; Song, Y.; Taguchi, S. Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) in recombinant Corynebacterium glutamicum using propionate as a precursor. J. Biotechnol. 2011, 152, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Rittmann, D.; Wendisch, V.F.; Bott, M.; Sahm, H. Global expression profiling and physiological characterization of Corynebacterium glutamicum grown in the presence of L-valine. Appl. Environ. Microbiol. 2003, 69, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature-triggered glutamate production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef] [PubMed]
- Polen, T.; Schluesener, D.; Poetsch, A.; Bott, M.; Wendisch, V.F. Characterization of citrate utilization in Corynebacterium glutamicum by transcriptome and proteome analysis. FEMS Microbiol. Lett. 2007, 273, 109–119. [Google Scholar] [CrossRef]
- Tsuge, Y.; Yamaguchi, A. Physiological characteristics of Corynebacterium glutamicum as a cell factory under anaerobic conditions. Appl. Microbiol. Biotechnol. 2021, 105, 6173–6181. [Google Scholar] [CrossRef]
- Yang, L.; Li, J.; Zhang, Y.; Chen, L.; Ouyang, Z.; Liao, D.; Zhao, F.; Han, S. Characterization of the enzyme kinetics of EMP and HMP pathway in Corynebacterium glutamicum: Reference for modeling metabolic networks. Front. Bioeng. Biotechnol. 2023, 11, 1296880. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, D.; Cai, M.; Zhang, H.; Pan, X.; You, J.; Zhang, X.; Xu, M.; Rao, Z. Combining protein and metabolic engineering strategies for high-level production of L-theanine in Corynebacterium glutamicum. Bioresour. Technol. 2024, 394, 130200. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Sun, W.; Leng, G.; Li, D.; Feng, C.; Tian, Z.; Wang, X. Enhanced fermentative γ-aminobutyric acid production by a metabolic engineered Corynebacterium glutamicum. Biotechnol. Bioprocess Eng. 2024, 29, 129–140. [Google Scholar] [CrossRef]
- Mutz, M.; Bruning, V.; Brusseler, C.; Muller, M.F.; Noack, S.; Marienhagen, J. Metabolic engineering of Corynebacterium glutamicum for the production of anthranilate from glucose and xylose. Microb. Biotechnol. 2024, 17, e14388. [Google Scholar] [CrossRef]
- Luckie, B.A.; Kashyap, M.; Pearson, A.N.; Chen, Y.; Liu, Y.; Valencia, L.E.; Carrillo Romero, A.; Hudson, G.A.; Tao, X.B.; Wu, B.; et al. Development of Corynebacterium glutamicum as a monoterpene production platform. Metab. Eng. 2024, 81, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Elkasaby, T.; Hanh, D.D.; Kahar, P.; Kawaguchi, H.; Sazuka, T.; Kondo, A.; Ogino, C. Utilization of sweet sorghum juice as a carbon source for enhancement of itaconic acid production in engineered Corynebacterium glutamicum. Enzym. Microb. Technol. 2024, 172, 110345. [Google Scholar] [CrossRef] [PubMed]
- Benninghaus, L.; Schwardmann, L.S.; Jilg, T.; Wendisch, V.F. Establishment of synthetic microbial consortia with Corynebacterium glutamicum and Pseudomonas putida: Design, construction, and application to production of gamma-glutamylisopropylamide and l-theanine. Microb. Biotechnol. 2024, 17, e14400. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Q.; Liu, H.; Wang, Y.; Zheng, Z.; Zhang, Y.; Tan, T. Engineering Corynebacterium glutamicum for the efficient production of N-acetylglucosamine. Bioresour. Technol. 2023, 390, 129865. [Google Scholar] [CrossRef]
- Desiderato, C.K.; Muller, C.; Schretzmeier, A.; Hasenauer, K.M.; Gnannt, B.; Supple, B.; Reiter, A.; Steier, V.; Oldiges, M.; Eikmanns, B.J.; et al. Optimized recombinant production of the bacteriocin garvicin Q by Corynebacterium glutamicum. Front. Microbiol. 2023, 14, 1254882. [Google Scholar] [CrossRef]
- Huang, D.; Wang, X.; Liu, W.B.; Ye, B.C. Remodeling metabolism of Corynebacterium glutamicum for high-level dencichine production. Bioresour. Technol. 2023, 388, 129800. [Google Scholar] [CrossRef]
- Schmollack, M.; Werner, F.; Huber, J.; Kiefer, D.; Merkel, M.; Hausmann, R.; Siebert, D.; Blombach, B. Metabolic engineering of Corynebacterium glutamicum for acetate-based itaconic acid production. Biotechnol. Biofuels Bioprod. 2022, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Baritugo, K.A.; Kim, H.T.; David, Y.; Choi, J.I.; Hong, S.H.; Jeong, K.J.; Choi, J.H.; Joo, J.C.; Park, S.J. Metabolic engineering of Corynebacterium glutamicum for fermentative production of chemicals in biorefinery. Appl. Microbiol. Biotechnol. 2018, 102, 3915–3937. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Jeong, K.J. Advances in Synthetic Biology Tools and Engineering of Corynebacterium glutamicum as a Platform Host for Recombinant Protein Production. Biotechnol. Bioprocess Eng. 2023, 28, 962–976. [Google Scholar] [CrossRef]
- Lv, X.; Li, Y.; Xiu, X.; Liao, C.; Xu, Y.; Liu, Y.; Li, J.; Du, G.; Liu, L. CRISPR genetic toolkits of classical food microorganisms: Current state and future prospects. Biotechnol. Adv. 2023, 69, 108261. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Nakagawa, S. The Corynebacterium glutamicum genome: Features and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 2003, 62, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.; Anand, U.; Jha, N.K.; Korzeniewska, E.; Bontempi, E.; Prockow, J.; Dey, A. The soil bacterium, Corynebacterium glutamicum, from biosynthesis of value-added products to bioremediation: A master of many trades. Environ. Res. 2022, 213, 113622. [Google Scholar] [CrossRef] [PubMed]
- Groussac, E.; Ortiz, M.; François, J. Improved protocols for quantitative determination of metabolites from biological samples using high performance ionic-exchange chromatography with conductimetric and pulsed amperometric detection. Enzym. Microb. Technol. 2000, 26, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, J.; Bathe, B.; Bartels, D.; Bischoff, N.; Bott, M.; Burkovski, A.; Dusch, N.; Eggeling, L.; Eikmanns, B.J.; Gaigalat, L.; et al. The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of L-aspartate-derived amino acids and vitamins. J. Biotechnol. 2003, 104, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Hermann, T.; Pfefferle, W.; Baumann, C.; Busker, E.; Schaffer, S.; Bott, M.; Sahm, H.; Dusch, N.; Kalinowski, J.; Pühler, A.; et al. Proteome analysis of Corynebacterium glutamicum. Electrophoresis 2001, 22, 1712–1723. [Google Scholar] [CrossRef]
- Bendt, A.K.; Burkovski, A.; Schaffer, S.; Bott, M.; Farwick, M.; Hermann, T. Towards a phosphoproteome map of Corynebacterium glutamicum. Proteomics 2003, 3, 1637–1646. [Google Scholar] [CrossRef]
- Schluesener, D.; Fischer, F.; Kruip, J.; Rogner, M.; Poetsch, A. Mapping the membrane proteome of Corynebacterium glutamicum. Proteomics 2005, 5, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Haussmann, U.; Qi, S.W.; Wolters, D.; Rogner, M.; Liu, S.J.; Poetsch, A. Physiological adaptation of Corynebacterium glutamicum to benzoate as alternative carbon source—A membrane proteome-centric view. Proteomics 2009, 9, 3635–3651. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A. Systems biology: Its practice and challenges. Cell 2005, 121, 511–513. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F.; Bott, M.; Kalinowski, J.; Oldiges, M.; Wiechert, W. Emerging Corynebacterium glutamicum systems biology. J. Biotechnol. 2006, 124, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Ludke, A.; Kramer, R.; Burkovski, A.; Schluesener, D.; Poetsch, A. A proteomic study of Corynebacterium glutamicum AAA+ protease FtsH. BMC Microbiol. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Fanous, A.; Weiss, W.; Gorg, A.; Jacob, F.; Parlar, H. A proteome analysis of the cadmium and mercury response in Corynebacterium glutamicum. Proteomics 2008, 8, 4976–4986. [Google Scholar] [CrossRef] [PubMed]
- Silberbach, M.; Schafer, M.; Huser, A.T.; Kalinowski, J.; Puhler, A.; Kramer, R.; Burkovski, A. Adaptation of Corynebacterium glutamicum to ammonium limitation: A global analysis using transcriptome and proteome techniques. Appl. Environ. Microbiol. 2005, 71, 2391–2402. [Google Scholar] [CrossRef]
- Dong, X.; Quinn, P.J.; Wang, X. Metabolic engineering of Escherichia coli and Corynebacterium glutamicum for the production of L-threonine. Biotechnol. Adv. 2011, 29, 11–23. [Google Scholar] [CrossRef]
- Rehm, N.; Burkovski, A. Engineering of nitrogen metabolism and its regulation in Corynebacterium glutamicum: Influence on amino acid pools and production. Appl. Microbiol. Biotechnol. 2011, 89, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Wittmann, C. Bio-based production of chemicals, materials and fuels—Corynebacterium glutamicum as versatile cell factory. Curr. Opin. Biotechnol. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Berlec, A.; Strukelj, B. Current state and recent advances in biopharmaceutical production in Escherichia coli, yeasts and mammalian cells. J. Ind. Microbiol. Biotechnol. 2013, 40, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Buschke, N.; Becker, J.; Schafer, R.; Kiefer, P.; Biedendieck, R.; Wittmann, C. Systems metabolic engineering of xylose-utilizing Corynebacterium glutamicum for production of 1,5-diaminopentane. Biotechnol. J. 2013, 8, 557–570. [Google Scholar] [CrossRef]
- Franzel, B.; Trotschel, C.; Ruckert, C.; Kalinowski, J.; Poetsch, A.; Wolters, D.A. Adaptation of Corynebacterium glutamicum to salt-stress conditions. Proteomics 2010, 10, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Nesvera, J.; Patek, M. Tools for genetic manipulations in Corynebacterium glutamicum and their applications. Appl. Microbiol. Biotechnol. 2011, 90, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, H.; Inui, M.; Yukawa, H. Transcriptional regulators of multiple genes involved in carbon metabolism in Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 114–125. [Google Scholar] [CrossRef]
- Patek, M.; Holatko, J.; Busche, T.; Kalinowski, J.; Nesvera, J. Corynebacterium glutamicum promoters: A practical approach. Microb. Biotechnol. 2013, 6, 103–117. [Google Scholar] [CrossRef]
- Kulis-Horn, R.K.; Persicke, M.; Kalinowski, J. Histidine biosynthesis, its regulation and biotechnological application in Corynebacterium glutamicum. Microb. Biotechnol. 2014, 7, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Blombach, B.; Gauttam, R.; Eikmanns, B.J. The RamA regulon: Complex regulatory interactions in relation to central metabolism in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2018, 102, 5901–5910. [Google Scholar] [CrossRef]
- Choi, J.W.; Jeon, E.J.; Jeong, K.J. Recent advances in engineering Corynebacterium glutamicum for utilization of hemicellulosic biomass. Curr. Opin. Biotechnol. 2019, 57, 17–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Xiao, S.; Qi, W.; Xu, J.; Yuan, Z.; Wang, Z. Engineering Corynebacterium glutamicum Mutants for 3-Methyl-1-butanol Production. Biochem. Genet. 2019, 57, 443–454. [Google Scholar] [CrossRef]
- Lee, C.-S.; Nam, J.-Y.; Son, E.-S.; Kwon, O.-c.; Han, W.; Cho, J.-Y.; Park, Y.-J. Next-generation sequencing-based genome-wide mutation analysis of L-lysine-producing Corynebacterium glutamicum ATCC 21300 strain. J. Microbiol. 2012, 50, 860–863. [Google Scholar] [CrossRef]
- Kim, H.-I.; Nam, J.-Y.; Cho, J.-Y.; Lee, C.-S.; Park, Y.-J. Next-generation sequencing-based transcriptome analysis of L-lysine-producing Corynebacterium glutamicum ATCC 21300 strain. J. Microbiol. 2013, 51, 877–880. [Google Scholar] [CrossRef]
- Cheng, F.; Luozhong, S.; Yu, H.; Guo, Z. Biosynthesis of Chondroitin in Engineered Corynebacterium glutamicum. J. Microbiol. Biotechnol. 2019, 29, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Yu, H.; Stephanopoulos, G. Engineering Corynebacterium glutamicum for high-titer biosynthesis of hyaluronic acid. Metab. Eng. 2019, 55, 276–289. [Google Scholar] [CrossRef]
- Hirasawa, T.; Kim, J.; Shirai, T.; Furusawa, C.; Shimizu, H. Molecular mechanisms and metabolic engineering of glutamate overproduction in Corynebacterium glutamicum. In Reprogramming Microbial Metabolic Pathways; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2012; Volume 64, pp. 261–281. [Google Scholar] [CrossRef]
- Eggeling, L.; Bott, M. A giant market and a powerful metabolism: L-lysine provided by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2015, 99, 3387–3394. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, Y.; Hasunuma, T.; Kondo, A. Recent advances in the metabolic engineering of Corynebacterium glutamicum for the production of lactate and succinate from renewable resources. J. Ind. Microbiol. Biotechnol. 2015, 42, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, D.; Xiong, B.; Zhang, C.; Bi, C. Engineering Corynebacterium glutamicum for violacein hyper production. Microb. Cell Factories 2016, 15, 148. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Quinn, P.J. Production of L-valine from metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2018, 102, 4319–4330. [Google Scholar] [CrossRef]
- Zha, J.; Zang, Y.; Mattozzi, M.; Plassmeier, J.; Gupta, M.; Wu, X.; Clarkson, S.; Koffas, M.A.G. Metabolic engineering of Corynebacterium glutamicum for anthocyanin production. Microb. Cell Factories 2018, 17, 143. [Google Scholar] [CrossRef]
- Wang, X. Strategy for improving L-isoleucine production efficiency in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2019, 103, 2101–2111. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Xu, J.Z.; Zhang, W.G. Metabolic engineering of l-leucine production in Escherichia coli and Corynebacterium glutamicum: A review. Crit. Rev. Biotechnol. 2019, 39, 633–647. [Google Scholar] [CrossRef]
- Kondoh, M.; Hirasawa, T. L-Cysteine production by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2019, 103, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.H.; Zhou, N.Y.; Liu, S.J. Degradation and assimilation of aromatic compounds by Corynebacterium glutamicum: Another potential for applications for this bacterium? Appl. Microbiol. Biotechnol. 2012, 95, 77–89. [Google Scholar] [CrossRef]
- Kogure, T.; Inui, M. Recent advances in metabolic engineering of Corynebacterium glutamicum for bioproduction of value-added aromatic chemicals and natural products. Appl. Microbiol. Biotechnol. 2018, 102, 8685–8705. [Google Scholar] [CrossRef]
- Becker, J.; Rohles, C.M.; Wittmann, C. Metabolically engineered Corynebacterium glutamicum for bio-based production of chemicals, fuels, materials, and healthcare products. Metab. Eng. 2018, 50, 122–141. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, F.; Wendisch, V.F. Transport and metabolic engineering of the cell factory Corynebacterium glutamicum. FEMS Microbiol. Lett. 2018, 365, fny166. [Google Scholar] [CrossRef]
- Trotschel, C.; Albaum, S.P.; Poetsch, A. Proteome turnover in bacteria: Current status for Corynebacterium glutamicum and related bacteria. Microb. Biotechnol. 2013, 6, 708–719. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, W.; Zhao, Z.; Dai, X.; Yang, Y.; Bai, Z. Protein secretion in Corynebacterium glutamicum. Crit. Rev. Biotechnol. 2017, 37, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, A.; Lindner, S.N.; Wendisch, V.F. Metabolic engineering of Corynebacterium glutamicum aimed at alternative carbon sources and new products. Comput. Struct. Biotechnol. J. 2012, 3, e201210004. [Google Scholar] [CrossRef]
- Zhao, N.; Qian, L.; Luo, G.; Zheng, S. Synthetic biology approaches to access renewable carbon source utilization in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2018, 102, 9517–9529. [Google Scholar] [CrossRef]
- Judith Becker, C.W. Industrial Microorganisms: Corynebacterium glutamicum. In Industrial Biotechnology; Wittmann, C., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; Volume 2. [Google Scholar]
- Lu, L.; Mao, Y.; Kou, M.; Cui, Z.; Jin, B.; Chang, Z.; Wang, Z.; Ma, H.; Chen, T. Engineering central pathways for industrial-level (3R)-acetoin biosynthesis in Corynebacterium glutamicum. Microb. Cell Factories 2020, 19, 102. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, J.Z.; Wang, B.; Rao, Z.M.; Zhang, W.G. L-valine production in Corynebacterium glutamicum based on systematic metabolic engineering: Progress and prospects. Amino Acids 2021, 53, 1301–1312. [Google Scholar] [CrossRef]
- Sheng, Q.; Wu, X.Y.; Xu, X.; Tan, X.; Li, Z.; Zhang, B. Production of l-glutamate family amino acids in Corynebacterium glutamicum: Physiological mechanism, genetic modulation, and prospects. Synth. Syst. Biotechnol. 2021, 6, 302–325. [Google Scholar] [CrossRef]
- Mhatre, A.; Shinde, S.; Jha, A.K.; Rodriguez, A.; Wardak, Z.; Jansen, A.; Gladden, J.M.; George, A.; Davis, R.W.; Varman, A.M. Corynebacterium glutamicum as an Efficient Omnivorous Microbial Host for the Bioconversion of Lignocellulosic Biomass. Front. Bioeng. Biotechnol. 2022, 10, 827386. [Google Scholar] [CrossRef]
- Kuhn, E.M.; Chen, X.; Tucker, M.P. Deacetylation and mechanical refining (DMR) and deacetylation and dilute acid (DDA) pretreatment of corn stover, switchgrass, and a 50:50 corn stover/switchgrass blend. ACS Sustain. Chem. Eng. 2020, 8, 6734–6743. [Google Scholar] [CrossRef]
- Wu, X.; Liu, J.; Liu, D.; Yuwen, M.; Koffas, M.A.G.; Zha, J. Biosynthesis of eriodictyol from tyrosine by Corynebacterium glutamicum. Microb. Cell Factories 2022, 21, 86. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y. Corynebacterium glutamicum Mechanosensing: From Osmoregulation to L-Glutamate Secretion for the Avian Microbiota-Gut-Brain Axis. Microorganisms 2021, 9, 201. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sun, P.; Wei, G.; Zhu, Y.; Li, J.; Liu, Y.; Chen, J.; Deng, Y. Efficient biosynthesis of creatine by whole-cell catalysis from guanidinoacetic acid in Corynebacterium glutamicum. Synth. Syst. Biotechnol. 2024, 9, 99–107. [Google Scholar] [CrossRef]
- Xu, G.; Wang, J.; Shen, J.; Zhu, Y.; Liu, W.; Chen, Y.; Zha, J.; Zhang, X.; Zhang, X.; Shi, J.; et al. Enhanced poly-gamma-glutamic acid synthesis in Corynebacterium glutamicum by reconstituting PgsBCA complex and fermentation optimization. Metab. Eng. 2024, 81, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Shi, F.; Xiang, Y.; Lai, W.; Ji, G. Establishment of CRISPR-Cpf1-assisted gene editing tool and engineering of 4-hydroxyisoleucine biosynthesis in Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2023, 39, 266. [Google Scholar] [CrossRef]
- Kyriakopoulos, S.; Kontoravdi, C. Analysis of the landscape of biologically-derived pharmaceuticals in Europe: Dominant production systems, molecule types on the rise and approval trends. Eur. J. Pharm. Sci. 2013, 48, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, D.; Tadele, L.R.; Lilge, L.; Henkel, M.; Hausmann, R. High-level recombinant protein production with Corynebacterium glutamicum using acetate as carbon source. Microb. Biotechnol. 2022, 15, 2744–2757. [Google Scholar] [CrossRef]
- Srivastava, P.; Deb, J.K. Construction of fusion vectors of corynebacteria: Expression of glutathione-S-transferase fusion protein in Corynebacterium acetoacidophilum ATCC 21476. FEMS Microbiol. Lett. 2002, 212, 209–216. [Google Scholar] [CrossRef]
- Rietschel, E.T.; Brade, H. Bacterial Endotoxins. Sci. Am. 1992, 267, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.F.; Hochstein, H.D.; Seligmann, E.B. The Limulus test for endotoxin (Pyrogen) in radiopharmaceuticals and biologicals. Bull. Parenter. Drug Assoc. 1972, 26, 153–162. [Google Scholar] [PubMed]
- Bobrovsky, P.A.; Kharlampieva, D.D.; Kirillin, S.A.; Brovina, K.A.; Grafskaia, E.N.; Lazarev, V.N.; Manuvera, V.A. Upregulation of YciM Expression Reduces Endotoxin Contamination of Recombinant Proteins Produced in Escherichia coli Cells. Biochemistry 2023, 88, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Mamat, U.; Wilke, K.; Bramhill, D.; Schromm, A.B.; Lindner, B.; Kohl, T.A.; Corchero, J.L.; Villaverde, A.; Schaffer, L.; Head, S.R.; et al. Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Factories 2015, 14, 57. [Google Scholar] [CrossRef]
- Sun, M.; Gao, X.; Zhao, Z.; Li, A.; Wang, Y.; Yang, Y.; Liu, X.; Bai, Z. Enhanced production of recombinant proteins in Corynebacterium glutamicum by constructing a bicistronic gene expression system. Microb. Cell Factories 2020, 19, 113. [Google Scholar] [CrossRef]
- Vertès, A.A. Protein Secretion Systems of Corynebacterium glutamicum. In Corynebacterium glutamicum: Biology and Biotechnology; Yukawa, H., Inui, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 351–389. [Google Scholar]
- Liu, X.; Yang, Y.; Zhang, W.; Sun, Y.; Peng, F.; Jeffrey, L.; Harvey, L.; McNeil, B.; Bai, Z. Expression of recombinant protein using Corynebacterium glutamicum: Progress, challenges and applications. Crit. Rev. Biotechnol. 2016, 36, 652–664. [Google Scholar] [CrossRef]
- Shen, J.; Chen, J.; Jensen, P.R.; Solem, C. A novel genetic tool for metabolic optimization of Corynebacterium glutamicum: Efficient and repetitive chromosomal integration of synthetic promoter-driven expression libraries. Appl. Microbiol. Biotechnol. 2017, 101, 4737–4746. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Liu, J.; Guo, Y.; Fan, L.; Ni, X.; Zheng, X.; Wang, M.; Zheng, P.; Sun, J.; et al. MACBETH: Multiplex automated Corynebacterium glutamicum base editing method. Metab. Eng. 2018, 47, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Wang, Y.; Yu, P.; Zheng, P.; Wang, M. Optimization of base editing in Corynebacterium glutamicum. Chin. J. Biotechnol. 2020, 36, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Fanous, A.; Hecker, M.; Gorg, A.; Parlar, H.; Jacob, F. Corynebacterium glutamicum as an indicator for environmental cobalt and silver stress—A proteome analysis. J. Environ. Sci. Health B 2010, 45, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Fanous, A.; Weiland, F.; Luck, C.; Gorg, A.; Friess, A.; Parlar, H. A proteome analysis of Corynebacterium glutamicum after exposure to the herbicide 2,4-dichlorophenoxy acetic acid (2,4-D). Chemosphere 2007, 69, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.Y.; Kim, J.; Park, G.; Kim, H.J.; Yang, J.; Seo, S.W. Synthetic biology tools for engineering Corynebacterium glutamicum. Comput. Struct. Biotechnol. J. 2023, 21, 1955–1965. [Google Scholar] [CrossRef] [PubMed]
- Harst, A.; Albaum, S.P.; Bojarzyn, T.; Trotschel, C.; Poetsch, A. Proteomics of FACS-sorted heterogeneous Corynebacterium glutamicum populations. J. Proteom. 2017, 160, 1–7. [Google Scholar] [CrossRef]
- Schulte, J.; Baumgart, M.; Bott, M. Development of a single-cell GlxR-based cAMP biosensor for Corynebacterium glutamicum. J. Biotechnol. 2017, 258, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, B.; Liu, Y.M.; Yang, K.Q.; Liu, S.J. New Intracellular Shikimic Acid Biosensor for Monitoring Shikimate Synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2018, 7, 591–601. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, P.; Xu, Z.; Kondo, A.; Wittmann, C.; Wendisch, V.F. Editorial: Engineering Corynebacterium glutamicum Chassis for Synthetic Biology, Biomanufacturing, and Bioremediation. Front. Bioeng. Biotechnol. 2022, 10, 923145. [Google Scholar] [CrossRef]
- Lei, M.; Peng, X.; Sun, W.; Zhang, D.; Wang, Z.; Yang, Z.; Zhang, C.; Yu, B.; Niu, H.; Ying, H.; et al. Nonsterile l-Lysine Fermentation Using Engineered Phosphite-Grown Corynebacterium glutamicum. ACS Omega 2021, 6, 10160–10167. [Google Scholar] [CrossRef]
- Liu, J.; Xu, J.Z.; Rao, Z.M.; Zhang, W.G. Industrial production of L-lysine in Corynebacterium glutamicum: Progress and prospects. Microbiol. Res. 2022, 262, 127101. [Google Scholar] [CrossRef] [PubMed]
- Gießelmann, G.; Dietrich, D.; Jungmann, L.; Kohlstedt, M.; Jeon, E.J.; Yim, S.S.; Sommer, F.; Zimmer, D.; Mühlhaus, T.; Schroda, M.; et al. Metabolic Engineering of Corynebacterium glutamicum for High-Level Ectoine Production: Design, Combinatorial Assembly, and Implementation of a Transcriptionally Balanced Heterologous Ectoine Pathway. Biotechnol. J. 2019, 14, 1800417. [Google Scholar] [CrossRef]
- Plassmeier, J.; Barsch, A.; Persicke, M.; Niehaus, K.; Kalinowski, J. Investigation of central carbon metabolism and the 2-methylcitrate cycle in Corynebacterium glutamicum by metabolic profiling using gas chromatography-mass spectrometry. J. Biotechnol. 2007, 130, 354–363. [Google Scholar] [CrossRef]
- Auchter, M.; Cramer, A.; Huser, A.; Ruckert, C.; Emer, D.; Schwarz, P.; Arndt, A.; Lange, C.; Kalinowski, J.; Wendisch, V.F.; et al. RamA and RamB are global transcriptional regulators in Corynebacterium glutamicum and control genes for enzymes of the central metabolism. J. Biotechnol. 2011, 154, 126–139. [Google Scholar] [CrossRef]
- Zhang, X.; Lai, L.; Xu, G.; Zhang, X.; Shi, J.; Koffas, M.A.G.; Xu, Z. Rewiring the Central Metabolic Pathway for High-Yield l-Serine Production in Corynebacterium glutamicum by Using Glucose. Biotechnol. J. 2019, 14, e1800497. [Google Scholar] [CrossRef]
- Wittmann, C.; Kiefer, P.; Zelder, O. Metabolic fluxes in Corynebacterium glutamicum during lysine production with sucrose as carbon source. Appl. Environ. Microbiol. 2004, 70, 7277–7287. [Google Scholar] [CrossRef]
- Muffler, A.; Bettermann, S.; Haushalter, M.; Hörlein, A.; Neveling, U.; Schramm, M.; Sorgenfrei, O. Quantitative determination of metabolic fluxes during coutilization of two carbon sources: Comparative analyses with Corynebacterium glutamicum during growth on acetate and/or glucose. J. Bacteriol. 2000, 182, 3088–3096. [Google Scholar] [CrossRef]
- Sauer, U.; Lasko, D.R.; Fiaux, J.; Hochuli, M.; Glaser, R.; Szyperski, T.; Wüthrich, K.; Bailey, J.E. Metabolic Flux Ratio Analysis of Genetic and Environmental Modulations of Escherichia coli Central Carbon Metabolism. J. Bacteriol. 1999, 181, 6679–6688. [Google Scholar] [CrossRef] [PubMed]
- Cocaign-Bousquet, M.; Guyonvarch, A.; Lindley, N.D. Growth Rate-Dependent Modulation of Carbon Flux through Central Metabolism and the Kinetic Consequences for Glucose-Limited Chemostat Cultures of Corynebacterium glutamicum. Appl. Environ. Microbiol. 1996, 62, 429–436. [Google Scholar] [CrossRef]
- Marx, A.; Graaf, A.A.d.; Wiechert, W.; Eggeling, L.; Sahm, H. Determination of the Fluxes in the Central Metabolism of Corynebacterium glutamicum by Nuclear Mag net i c Resonance Spectroscopy Combined with Metabolite Balancing. Biotechnol. Bioeng. 1995, 49, 111–129. [Google Scholar] [CrossRef]
- Takaç, S.; Çalık, G.; Mavituna, F.; Dervakos, G. Metabolic flux distribution for the optimized production of l-glutamate. Enzym. Microb. Technol. 1998, 23, 286–300. [Google Scholar] [CrossRef]
- Marx, A.; Striegel, K.; Graaf, A.A.d.; Sahm, H.; Eggeling, L. Response of the Central Metabolism of Corynebacterium glutamicum to Different Flux Burdens. Biotechnol. Bioeng. 1997, 56, 168–180. [Google Scholar] [CrossRef]
- Hirasawa, T.; Wachi, M. Glutamate Fermentation-2: Mechanism of L-Glutamate Overproduction in Corynebacterium glutamicum. Adv. Biochem. Eng. Biotechnol. 2017, 159, 57–72. [Google Scholar] [CrossRef]
- Noack, S.; Voges, R.; Gatgens, J.; Wiechert, W. The linkage between nutrient supply, intracellular enzyme abundances and bacterial growth: New evidences from the central carbon metabolism of Corynebacterium glutamicum. J. Biotechnol. 2017, 258, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, S. Comparative analysis of Corynebacterium glutamicum genomes: A new perspective for the industrial production of amino acids. BMC Genom. 2017, 18, 940. [Google Scholar] [CrossRef] [PubMed]
- Kolb, A.; Busby, S.; Buc, H.; Garges, S.; Adhya, S. Transcriptional regulation by cAMP and its receptor protein. Annu. Rev. Biochem. 1993, 62, 749–795. [Google Scholar] [CrossRef] [PubMed]
- De Crombrugghe, B.; Busby, S.; Buc, H. Cyclic AMP Receptor Protein: Role in Transcription Activation. Science 1984, 224, 831–838. [Google Scholar] [CrossRef]
- Han, S.O.; Inui, M.; Yukawa, H. Expression of Corynebacterium glutamicum glycolytic genes varies with carbon source and growth phase. Microbiology 2007, 153, 2190–2202. [Google Scholar] [CrossRef]
- Muffler, A.; Bettermann, S.; Haushalter, M.; Hörlein, A.; Neveling, U.; Schramm, M.; Sorgenfrei, O. Genome-wide transcription profiling of Corynebacterium glutamicum after heat shock and during growth on acetate and glucose†. J. Bacteriol. 2002, 98, 255–268. [Google Scholar] [CrossRef]
- Han, S.O.; Inui, M.; Yukawa, H. Effect of carbon source availability and growth phase on expression of Corynebacterium glutamicum genes involved in the tricarboxylic acid cycle and glyoxylate bypass. Microbiology 2008, 154, 3073–3083. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Vertès, A.A.; Okino, S.; Inui, M.; Yukawa, H. Engineering of a xylose metabolic pathway in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2006, 72, 3418–3428. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; He, Z.-n.; Shi, Z.-p.; Enock, M. Improving the stability of glutamate fermentation by Corynebacterium glutamicum via supplementing sorbitol or glycerol. Bioresour. Bioprocess. 2015, 2, 9. [Google Scholar] [CrossRef]
- Schmid, R.; Uhlemann, E.-M.; Nolden, L.; Wersch, G.; Hecker, R.; Hermann, T.; Marx, A.; Burkovski, A. Response to nitrogen starvation in Corynebacterium glutamicum. FEMS Microbiol. Lett. 2000, 187, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Chen, Y.; Gong, A.D. Development of a defined medium for Corynebacterium glutamicum using urea as nitrogen source. 3 Biotech 2021, 11, 405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, J.; Jin, Z.; Xia, X.; Zhang, W. Improvement of acetyl-CoA supply and glucose utilization increases l-leucine production in Corynebacterium glutamicum. Biotechnol. J. 2022, 17, e2100349. [Google Scholar] [CrossRef]
- Wang, J.Y.; Rao, Z.M.; Xu, J.Z.; Zhang, W.G. Enhancing beta-alanine production from glucose in genetically modified Corynebacterium glutamicum by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2021, 105, 9153–9166. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, K.; Osamura, T.; Takahashi, F. A 4-hydroxybenzoate 3-hydroxylase mutant enables 4-amino-3-hydroxybenzoic acid production from glucose in Corynebacterium glutamicum. Microb. Cell Factories 2023, 22, 168. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jin, H.; Liu, W.; Wang, Q.; Qi, Q. Engineering Corynebacterium glutamicum to produce 5-aminolevulinic acid from glucose. Microb. Cell Factories 2015, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Kallscheuer, N.; Marienhagen, J. Corynebacterium glutamicum as platform for the production of hydroxybenzoic acids. Microb. Cell Factories 2018, 17, 70. [Google Scholar] [CrossRef]
- Guo, Y.-S.; Yang, Z.-D.; Huang, J.-S.; Gao, J.-Y.; Chen, X.-P.; Cheng, H.; Zhang, P.-J.; Su, H.-H. Efficient and economical biosynthesis of high-purity isomaltulose from sugar industrial waste molasses using an engineered Corynebacterium glutamicum strain. Green Chem. 2022, 24, 4050–4060. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Dou, W.; Xu, X.; Xu, Z. Enhancement of L-Serine Production by Corynebacterium glutamicum SYPS-062 Directly from Sucrose. Trop. J. Pharm. Res. 2015, 13, 2011–2014. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Bu, Y.F.; Liu, J.Z. Production of L-ornithine from sucrose and molasses by recombinant Corynebacterium glutamicum. Folia Microbiol. 2015, 60, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Acton, Q.A. Aminobenzoic Acids—Advances in Research and Application: 2013 Edition: ScholarlyBrief; ScholarlyEditions: Atlanta, GA, USA, 2013. [Google Scholar]
- Li, Z.; Dong, Y.; Liu, Y.; Cen, X.; Liu, D.; Chen, Z. Systems metabolic engineering of Corynebacterium glutamicum for high-level production of 1,3-propanediol from glucose and xylose. Metab. Eng. 2022, 70, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Jiang, Y.; Li, Z.; Wang, F.; Wu, X.Y. Recent Progress on Chemical Production From Non-food Renewable Feedstocks Using Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2020, 8, 606047. [Google Scholar] [CrossRef] [PubMed]
- Uhde, A.; Youn, J.W.; Maeda, T.; Clermont, L.; Matano, C.; Kramer, R.; Wendisch, V.F.; Seibold, G.M.; Marin, K. Glucosamine as carbon source for amino acid-producing Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2013, 97, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, F.J. Organic Forms of Soil Nitrogen. In Nitrogen in Agricultural Soils; Wiley: Hoboken, NJ, USA, 1982; pp. 67–122. [Google Scholar]
- Appuhn, A.; Joergensen, R.G.; Raubuch, M.; Scheller, E.; Wilke, B. The automated determination of glucosamine, galactosamine, muramic acid, and mannosamine in soil and root hydrolysates by HPLC. J. Plant Nutr. Soil Sci. 2004, 167, 17–21. [Google Scholar] [CrossRef]
- Parsons, J. Chemistry and distribution of amino sugars in soils and soil organisms. In Soil Biochemistry; CRC Press: Boca Raton, FL, USA, 2021; pp. 197–228. [Google Scholar]
- Nguyen, A.Q.; Schneider, J.; Reddy, G.K.; Wendisch, V.F. Fermentative production of the diamine putrescine: System metabolic engineering of Corynebacterium glutamicum. Metabolites 2015, 5, 211–231. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | C. glutamicum | E. coli |
|---|---|---|
| Morphology | Rod-shaped bacterium | Rod-shaped bacterium |
| Staining properties | Gram (+) | Gram (−) |
| Genome size | 3.28 Mbp | 4.6 Mbp |
| Habitat environment | Soil and plants | Human and animal intestines |
| Growing environment | Mainly aerobic | Facultative anaerobic |
| Bacterial endotoxin | X | O |
| Necessary to form an inclusion body upon recovery | X | O |
| Expression protein secretion form | High solubility | Low solubility |
| Plasmid vector optimization | O | O |
| Protein secretion pathway study | O | O |
| Safety | GRAS | Most are non-pathogenic * |
| Industrial application | Amino acid biosynthesis, various compounds biosynthesis, metabolic engineering | Protein production, antibody production, gene cloning, metabolic engineering |
| Carbon Source | Production | Reference |
|---|---|---|
| Glucose | L-serine | [139] |
| β-alanine | [142] | |
| 4-amino-3-hydroxybenzoic acid | [143] | |
| 5-aminolevulinic acid | [144] | |
| N-acetylglucosamine | [31] | |
| Putrescine | [156] | |
| γ-aminobutyric acid | [26] | |
| L-cysteine | [76] | |
| Violacein | [71] | |
| Glucose and xylose | 1,3-propanediol | [150] |
| Anthranilate | [27] | |
| Xylose | Cadaverin | [55] |
| Sucrose | L-serine | [147] |
| L-ornithine | [148] | |
| Acetate | Itaconic acid | [34] |
| Recombinant protein | [97] | |
| Glucosamine | L-lysine, putrescine | [152] |
| Molasses | Isomaltulose | [146] |
| Itaconic acid | [29] | |
| Starch and lignocellulose | Lactate, succinate | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Lim, S. Review of the Proteomics and Metabolic Properties of Corynebacterium glutamicum. Microorganisms 2024, 12, 1681. https://doi.org/10.3390/microorganisms12081681
Park J, Lim S. Review of the Proteomics and Metabolic Properties of Corynebacterium glutamicum. Microorganisms. 2024; 12(8):1681. https://doi.org/10.3390/microorganisms12081681
Chicago/Turabian StylePark, Juhwan, and Sooa Lim. 2024. "Review of the Proteomics and Metabolic Properties of Corynebacterium glutamicum" Microorganisms 12, no. 8: 1681. https://doi.org/10.3390/microorganisms12081681