
Development of the Intestinal Microbiota of Dairy Calves and Changes Associated with Cryptosporidium spp. Infection in Brazil
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sampling
2.3. DNA Extraction and Pathogen Detection
2.4. Statistical Analysis
3. Results
3.1. Incidence of Diseases
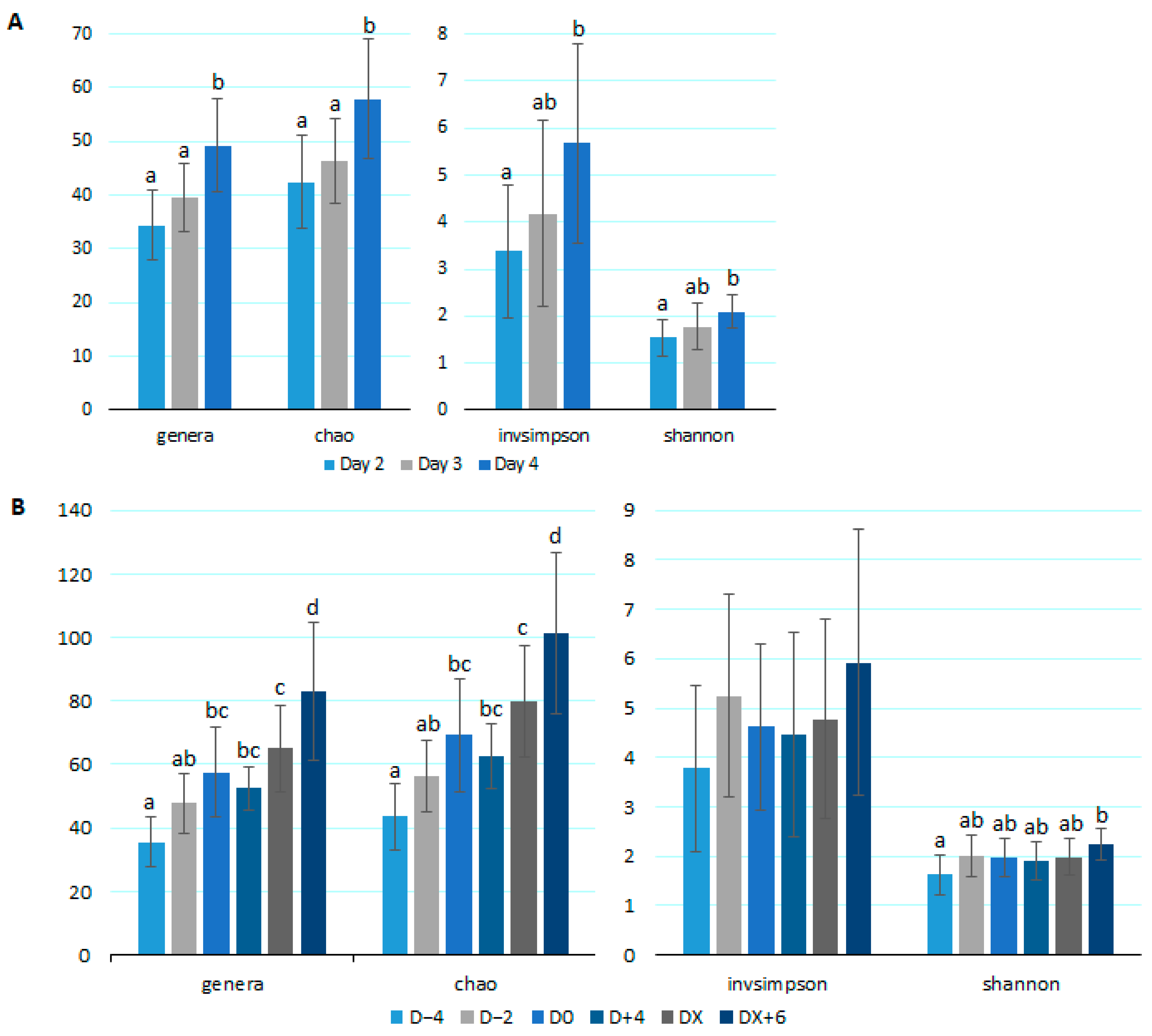
3.2. Microbiota Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; LeBlanc, S.J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Thomson, S.; Hamilton, C.A.; Hope, J.C.; Katzer, F.; Mabbott, N.A.; Morrison, L.J.; Innes, E.A. Bovine cryptosporidiosis: Impact, host-parasite interaction and control strategies. Vet. Res. 2017, 48, 42. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.; Martins, F.D.C.; Ladeia, W.A.; Cortela, I.B.; Valadares, M.F.; Matos, A.; Caldart, E.T.; Ayres, H.; Navarro, I.T.; Freire, R.L. Identification, molecular characterization and factors associated with occurrences of Cryptosporidium spp. in calves on dairy farms in Brazil. Rev. Bras. Parasitol. Vet. 2021, 30, e009621. [Google Scholar] [CrossRef] [PubMed]
- Meireles, M.V.; de Oliveira, F.P.; Teixeira, W.F.; Coelho, W.M.; Mendes, L.C. Molecular characterization of Cryptosporidium spp. in dairy calves from the state of Sao Paulo, Brazil. Parasitol. Res. 2011, 109, 949–951. [Google Scholar] [CrossRef]
- Santin, M. Clinical and subclinical infections with Cryptosporidium in animals. N. Z. Vet. J. 2013, 61, 1–10. [Google Scholar] [CrossRef]
- Thompson, R.C.; Smith, A. Zoonotic enteric protozoa. Vet. Parasitol. 2011, 182, 70–78. [Google Scholar] [CrossRef]
- Zambriski, J.A.; Nydam, D.V.; Wilcox, Z.J.; Bowman, D.D.; Mohammed, H.O.; Liotta, J.L. Cryptosporidium parvum: Determination of ID50 and the dose-response relationship in experimentally challenged dairy calves. Vet. Parasitol. 2013, 197, 104–112. [Google Scholar] [CrossRef]
- Arsenopoulos, K.; Theodoridis, A.; Papadopoulos, E. Effect of colostrum quantity and quality on neonatal calf diarrhoea due to Cryptosporidium spp. infection. Comp. Immunol. Microbiol. Infect. Dis. 2017, 53, 50–55. [Google Scholar] [CrossRef]
- Hartinger, T.; Pacifico, C.; Poier, G.; Terler, G.; Klevenhusen, F.; Zebeli, Q. Shift of dietary carbohydrate source from milk to various solid feeds reshapes the rumen and fecal microbiome in calves. Sci. Rep. 2022, 12, 12383. [Google Scholar] [CrossRef]
- Dill-McFarland, K.A.; Breaker, J.D.; Suen, G. Microbial succession in the gastrointestinal tract of dairy cows from 2 weeks to first lactation. Sci. Rep. 2017, 7, 40864. [Google Scholar] [CrossRef]
- Gomez, D.E.; Arroyo, L.G.; Costa, M.C.; Viel, L.; Weese, J.S. Characterization of the Fecal Bacterial Microbiota of Healthy and Diarrheic Dairy Calves. J. Vet. Intern. Med. 2017, 31, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Dall Agnol, A.M.; Lorenzetti, E.; Leme, R.A.; Ladeia, W.A.; Mainardi, R.M.; Bernardi, A.; Headley, S.A.; Freire, R.L.; Pereira, U.P.; Alfieri, A.F.; et al. Severe outbreak of bovine neonatal diarrhea in a dairy calf rearing unit with multifactorial etiology. Braz. J. Microbiol. 2021, 52, 2547–2553. [Google Scholar] [CrossRef] [PubMed]
- Cruvinel, L.B.; Ayres, H.; Zapa, D.M.B.; Nicaretta, J.E.; Couto, L.F.M.; Heller, L.M.; Bastos, T.S.A.; Cruz, B.C.; Soares, V.E.; Teixeira, W.F.; et al. Prevalence and risk factors for agents causing diarrhea (Coronavirus, Rotavirus, Cryptosporidium spp., Eimeria spp., and nematodes helminthes) according to age in dairy calves from Brazil. Trop. Anim. Health Prod. 2020, 52, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.Q.; Gomes, L.A.; Santos, I.S.; Alfieri, A.F.; Weese, J.S.; Costa, M.C. Fecal microbiota transplantation in puppies with canine parvovirus infection. J. Vet. Intern. Med. 2018, 32, 707–711. [Google Scholar] [CrossRef]
- Kim, H.S.; Whon, T.W.; Sung, H.; Jeong, Y.S.; Jung, E.S.; Shin, N.R.; Hyun, D.W.; Kim, P.S.; Lee, J.Y.; Lee, C.H.; et al. Longitudinal evaluation of fecal microbiota transplantation for ameliorating calf diarrhea and improving growth performance. Nat. Commun. 2021, 12, 161. [Google Scholar] [CrossRef]
- Slanzon, G.S.; Ridenhour, B.J.; Parrish, L.M.; Trombetta, S.C.; Moore, D.A.; Sischo, W.M.; McConnel, C.S. Effects of a farm-specific fecal microbial transplant (FMT) product on clinical outcomes and fecal microbiome composition in preweaned dairy calves. PLoS ONE 2022, 17, e0276638. [Google Scholar] [CrossRef]
- Islam, J.; Tanimizu, M.; Shimizu, Y.; Goto, Y.; Ohtani, N.; Sugiyama, K.; Tatezaki, E.; Sato, M.; Makino, E.; Shimada, T.; et al. Development of a rational framework for the therapeutic efficacy of fecal microbiota transplantation for calf diarrhea treatment. Microbiome 2022, 10, 31. [Google Scholar] [CrossRef]
- Gamsjager, L.; Cirone, K.M.; Schluessel, S.; Campsall, M.; Herik, A.; Lahiri, P.; Young, D.; Dufour, A.; Sapountzis, P.; Otani, S.; et al. Host innate immune responses and microbiome profile of neonatal calves challenged with Cryptosporidium parvum and the effect of bovine colostrum supplementation. Front. Cell Infect. Microbiol. 2023, 13, 1165312. [Google Scholar] [CrossRef]
- Ichikawa-Seki, M.; Motooka, D.; Kinami, A.; Murakoshi, F.; Takahashi, Y.; Aita, J.; Hayashi, K.; Tashibu, A.; Nakamura, S.; Iida, T.; et al. Specific increase of Fusobacterium in the faecal microbiota of neonatal calves infected with Cryptosporidium parvum. Sci. Rep. 2019, 9, 12517. [Google Scholar] [CrossRef]
- Dorbek-Kolin, E.; Husso, A.; Niku, M.; Loch, M.; Pessa-Morikawa, T.; Niine, T.; Kaart, T.; Iivanainen, A.; Orro, T. Faecal microbiota in two-week-old female dairy calves during acute cryptosporidiosis outbreak—Association with systemic inflammatory response. Res. Vet. Sci. 2022, 151, 116–127. [Google Scholar] [CrossRef]
- Mammeri, M.; Obregon, D.A.; Chevillot, A.; Polack, B.; Julien, C.; Pollet, T.; Cabezas-Cruz, A.; Adjou, K.T. Cryptosporidium parvum Infection Depletes Butyrate Producer Bacteria in Goat Kid Microbiome. Front. Microbiol. 2020, 11, 548737. [Google Scholar] [CrossRef]
- Hares, M.F.; Griffiths, B.E.; Johnson, F.; Nelson, C.; Haldenby, S.; Stewart, C.J.; Duncan, J.S.; Oikonomou, G.; Coombes, J.L. Specific pathway abundances in the neonatal calf faecal microbiome are associated with susceptibility to Cryptosporidium parvum infection: A metagenomic analysis. Anim. Microbiome 2023, 5, 43. [Google Scholar] [CrossRef]
- Xiao, L.; Alderisio, K.; Limor, J.; Royer, M.; Lal, A.A. Identification of species and sources of Cryptosporidium oocysts in storm waters with a small-subunit rRNA-based diagnostic and genotyping tool. Appl. Environ. Microbiol. 2000, 66, 5492–5498. [Google Scholar] [CrossRef]
- Herring, A.J.; Inglis, N.F.; Ojeh, C.K.; Snodgrass, D.R.; Menzies, J.D. Rapid diagnosis of rotavirus infection by direct detection of viral nucleic acid in silver-stained polyacrylamide gels. J. Clin. Microbiol. 1982, 16, 473–477. [Google Scholar] [CrossRef]
- Takiuchi, E.; Stipp, D.T.; Alfieri, A.F.; Alfieri, A.A. Improved detection of bovine coronavirus N gene in faeces of calves infected naturally by a semi-nested PCR assay and an internal control. J. Virol. Methods 2006, 131, 148–154. [Google Scholar] [CrossRef]
- Rahn, K.; De Grandis, S.A.; Clarke, R.C.; McEwen, S.A.; Galan, J.E.; Ginocchio, C.; Curtiss, R., 3rd; Gyles, C.L. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Malorny, B.; Hoorfar, J.; Bunge, C.; Helmuth, R. Multicenter validation of the analytical accuracy of Salmonella PCR: Towards an international standard. Appl. Environ. Microbiol. 2003, 69, 290–296. [Google Scholar] [CrossRef]
- Franck, S.M.; Bosworth, B.T.; Moon, H.W. Multiplex PCR for enterotoxigenic, attaching and effacing, and Shiga toxin-producing Escherichia coli strains from calves. J. Clin. Microbiol. 1998, 36, 1795–1797. [Google Scholar] [CrossRef]
- Andrade, G.I.; Coura, F.M.; Santos, E.L.; Ferreira, M.G.; Galinari, G.C.; Facury Filho, E.J.; de Carvalho, A.U.; Lage, A.P.; Heinemann, M.B. Identification of virulence factors by multiplex PCR in Escherichia coli isolated from calves in Minas Gerais, Brazil. Trop. Anim. Health Prod. 2012, 44, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Ishaq, S.L.; Bichi, E.; Olivo, S.K.; Lowe, J.; Aldridge, B.M. Biogeographical Differences in the Influence of Maternal Microbial Sources on the Early Successional Development of the Bovine Neonatal Gastrointestinal tract. Sci. Rep. 2018, 8, 3197. [Google Scholar] [CrossRef]
- Klein-Jobstl, D.; Quijada, N.M.; Dzieciol, M.; Feldbacher, B.; Wagner, M.; Drillich, M.; Schmitz-Esser, S.; Mann, E. Microbiota of newborn calves and their mothers reveals possible transfer routes for newborn calves’ gastrointestinal microbiota. PLoS ONE 2019, 14, e0220554. [Google Scholar] [CrossRef]
- Alipour, M.J.; Jalanka, J.; Pessa-Morikawa, T.; Kokkonen, T.; Satokari, R.; Hynonen, U.; Iivanainen, A.; Niku, M. The composition of the perinatal intestinal microbiota in cattle. Sci. Rep. 2018, 8, 10437. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Zambriski, J.A.; Nydam, D.V.; Bowman, D.D.; Bellosa, M.L.; Burton, A.J.; Linden, T.C.; Liotta, J.L.; Ollivett, T.L.; Tondello-Martins, L.; Mohammed, H.O. Description of fecal shedding of Cryptosporidium parvum oocysts in experimentally challenged dairy calves. Parasitol. Res. 2013, 112, 1247–1254. [Google Scholar] [CrossRef]
- Bellosa, M.L.; Nydam, D.V.; Liotta, J.L.; Zambriski, J.A.; Linden, T.C.; Bowman, D.D. A comparison of fecal percent dry matter and number of Cryptosporidium parvum oocysts shed to observational fecal consistency scoring in dairy calves. J. Parasitol. 2011, 97, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Operario, D.J.; Bristol, L.S.; Liotta, J.; Nydam, D.V.; Houpt, E.R. Correlation between diarrhea severity and oocyst count via quantitative PCR or fluorescence microscopy in experimental cryptosporidiosis in calves. Am. J. Trop. Med. Hyg. 2015, 92, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Santin, M.; Trout, J.M.; Xiao, L.; Zhou, L.; Greiner, E.; Fayer, R. Prevalence and age-related variation of Cryptosporidium species and genotypes in dairy calves. Vet. Parasitol. 2004, 122, 103–117. [Google Scholar] [CrossRef]
- Trotz-Williams, L.A.; Jarvie, B.D.; Martin, S.W.; Leslie, K.E.; Peregrine, A.S. Prevalence of Cryptosporidium parvum infection in southwestern Ontario and its association with diarrhea in neonatal dairy calves. Can. Vet. J. 2005, 46, 349–351. [Google Scholar]
- Fayer, R.; Gasbarre, L.; Pasquali, P.; Canals, A.; Almeria, S.; Zarlenga, D. Cryptosporidium parvum infection in bovine neonates: Dynamic clinical, parasitic and immunologic patterns. Int. J. Parasitol. 1998, 28, 49–56. [Google Scholar] [CrossRef]
- Weese, J.S.; Jelinski, M. Assessment of the Fecal Microbiota in Beef Calves. J. Vet. Intern. Med. 2017, 31, 176–185. [Google Scholar] [CrossRef]
- Bessegatto, J.A.; Lisboa, J.A.N.; Santos, B.P.; Curti, J.M.; Montemor, C.; Alfieri, A.A.; Mach, N.; Costa, M.C. Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture. Animals 2024, 14, 1447. [Google Scholar] [CrossRef]
- Mammeri, M.; Chevillot, A.; Thomas, M.; Julien, C.; Auclair, E.; Pollet, T.; Polack, B.; Vallee, I.; Adjou, K.T. Cryptosporidium parvum-Infected Neonatal Mice Show Gut Microbiota Remodelling Using High-Throughput Sequencing Analysis: Preliminary Results. Acta Parasitol. 2019, 64, 268–275. [Google Scholar] [CrossRef]
- Ras, R.; Huynh, K.; Desoky, E.; Badawy, A.; Widmer, G. Perturbation of the intestinal microbiota of mice infected with Cryptosporidium parvum. Int. J. Parasitol. 2015, 45, 567–573. [Google Scholar] [CrossRef]
- Jessop, E.; Li, L.; Renaud, D.L.; Verbrugghe, A.; Macnicol, J.; Gamsjager, L.; Gomez, D.E. Neonatal Calf Diarrhea and Gastrointestinal Microbiota: Etiologic Agents and Microbiota Manipulation for Treatment and Prevention of Diarrhea. Vet. Sci. 2024, 11, 108. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessegatto, J.A.; Lisbôa, J.A.N.; Martins, F.D.C.; Freire, R.L.; Facury Filho, E.J.; Alfieri, A.A.; Costa, M.C. Development of the Intestinal Microbiota of Dairy Calves and Changes Associated with Cryptosporidium spp. Infection in Brazil. Microorganisms 2024, 12, 1744. https://doi.org/10.3390/microorganisms12091744
Bessegatto JA, Lisbôa JAN, Martins FDC, Freire RL, Facury Filho EJ, Alfieri AA, Costa MC. Development of the Intestinal Microbiota of Dairy Calves and Changes Associated with Cryptosporidium spp. Infection in Brazil. Microorganisms. 2024; 12(9):1744. https://doi.org/10.3390/microorganisms12091744
Chicago/Turabian StyleBessegatto, José Antônio, Júlio Augusto Naylor Lisbôa, Felippe Danyel Cardoso Martins, Roberta Lemos Freire, Elias Jorge Facury Filho, Amauri Alcindo Alfieri, and Marcio C. Costa. 2024. "Development of the Intestinal Microbiota of Dairy Calves and Changes Associated with Cryptosporidium spp. Infection in Brazil" Microorganisms 12, no. 9: 1744. https://doi.org/10.3390/microorganisms12091744