
The Effects of Mixed Robinia pseudoacacia and Quercus variabilis Plantation on Soil Bacterial Community Structure and Nitrogen-Cycling Gene Abundance in the Southern Taihang Mountain Foothills

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Experimental Design
2.3. Determination of Soil Physicochemical Properties
2.4. Soil DNA Extraction and Sequencing
2.5. Real-Time Fluorescent Quantitative PCR Analysis
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Properties of Soils from Different Forest Types
3.2. Diversity of Bacterial Communities in Different Forest Soils
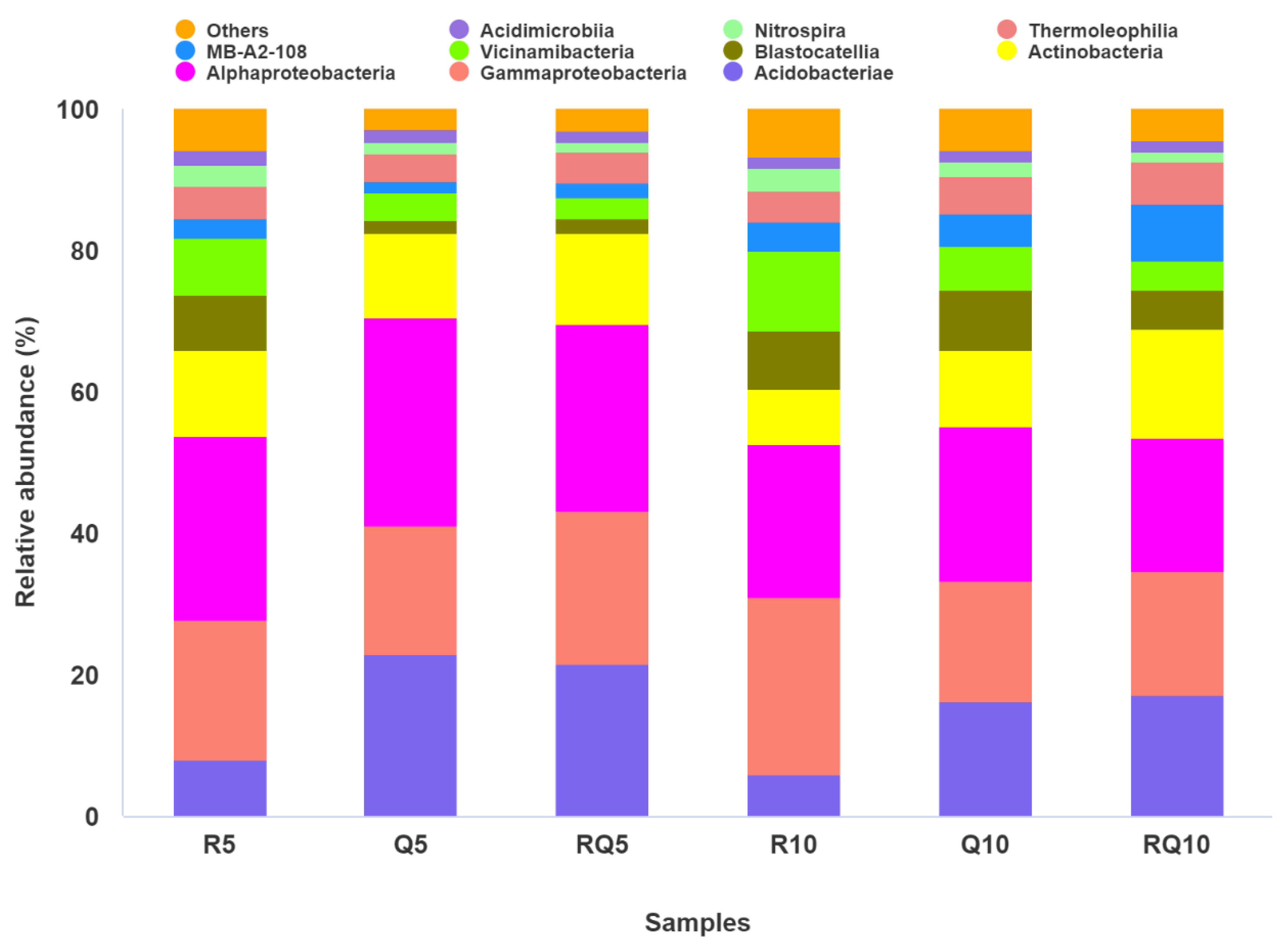
3.3. Composition and Structure of Soil Bacterial Communities in Different Forest Stands
3.4. Ecological Function Prediction of Bacterial Communities in Different Forest Types
3.5. Abundance of Nitrogen-Cycling Functional Genes in Different Forest Types
4. Discussion
4.1. Physicochemical Properties of Soil in Different Forest Stands
4.2. Bacterial Community Diversity in Different Forest Stands
4.3. Soil Bacterial Community Composition of Different Forest Types
4.4. Nitrogen Cycle Functional Gene Abundance in Different Forest Types
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Heijden, M.G.A.V.D.; Bardgett, R.D.; Straalen, N.M.V. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Comerford, N.B.; Franzluebbers, A.J.; Stromberger, M.E.; Morris, L.; Markewitz, D.; Moore, R. Assessment and evaluation of soil ecosystem services. Soil Horiz. 2013, 54, 1–14. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Sharma, B.; Nagpal, S.; Kumar, A.; Tiwari, S.; Nair, R.M. Plant growth-promoting rhizobacteria: Salt stress alleviators to improve crop productivity for sustainable agriculture development. Front. Plant Sci. 2023, 13, 1101862. [Google Scholar] [CrossRef]
- Preece, C.; Verbruggen, E.; Liu, L.; Weedon, J.T.; Peñuelas, J. Effects of past and current drought on the composition and diversity of soil microbial communities. Soil Biol. Biochem. 2019, 131, 28–39. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and Soil Microbial Community: A Review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Sofo, A.; Mininni, A.N.; Ricciuti, P. Soil Macrofauna: A key Factor for Increasing Soil Fertility and Promoting Sustainable Soil Use in Fruit Orchard Agrosystems. Agronomy 2020, 10, 456. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef] [PubMed]
- Nacke, H.; Goldmann, K.; Schöning, I.; Pfeiffer, B.; Kaiser, K.; Castillo-Villamizar, G.A.; Schrumpf, M.; Buscot, F.; Daniel, R.; Wubet, T. Fine spatial scale variation of soil microbial communities under European Beech and Norway Spruce. Front. Microbiol. 2016, 7, 2067. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H.; del Río, M.; Ammer Ch Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; Fabrika, M.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2015, 134, 927–947. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 2015, 90, 444–466. [Google Scholar] [CrossRef]
- Jin, X.; Liu, Y.; Hu, W.; Wang, G.; Kong, Z.; Wu, L.; Ge, G. Soil Bacterial and Fungal Communities and the Associated Nutrient Cycling Responses to Forest Conversion after Selective Logging in a Subtropical Forest of China. For. Ecol. Manag. 2019, 444, 308–317. [Google Scholar] [CrossRef]
- Nie, H.; Qin, T.; Yan, D.; Lv, X.; Wang, J.; Huang, Y.; Lv, Z.; Liu, S.; Liu, F. How do tree species characteristics affect the bacterial community structure of subtropical natural mixed forests? Sci. Total Environ. 2021, 764, 144633. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; He, R. Soil microbial community characteristics under different vegetation types at the national nature reserve of Xiaolongshan Mountains, Northwest China. Ecol. Inform. 2020, 55, 101020. [Google Scholar] [CrossRef]
- Aleklett, K.; Leff, J.W.; Fierer, N.; Hart, M. Wild plant species growing closely connected in a subalpine meadow host distinct root-associated bacterial communities. PeerJ 2015, 2015, e804. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef]
- Yao, F.; Yang, S.; Wang, Z.; Wang, X.; Ye, J.; Wang, X.; DeBruyn, J.M.; Feng, X.; Jiang, Y.; Li, H. Microbial Taxa Distribution Is Associated with Ecological Trophic Cascades along an Elevation Gradient. Front. Microbiol. 2017, 8, 2071. [Google Scholar] [CrossRef]
- Benizri, E.; Amiaud, B. Relationship between plants and soil microbial communities in fertilized grasslands. Soil Biol. Biochem. 2005, 37, 2055–2064. [Google Scholar] [CrossRef]
- de Deyn, G.B.; van der, P.W.H. Linking aboveground and belowground diversity. Trends Ecol. Evol. 2005, 20, 625–633. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Buma, D.S.; de Boer, W.; Klinkhamer, P.G.L.; van Veen, J.A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie Leeuwenhoek 2002, 81, 509. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef]
- Thamdrup, B. New pathways and processes in the global nitrogen cycle. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 407–428. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Feng, Y.; Wang, X.; Yu, Q.; Zhang, F.; Yang, G.; Ren, G.; Han, X.; Wang, X.; Ren, C. Soil microbial nitrogen-cycling gene abundances in response to crop diversification: A meta-analysis. Sci. Total Environ. 2022, 838, 156621. [Google Scholar] [CrossRef]
- Rosenkranz, S.; Wilcke, W.; Eisenhauer, N.; Oelmann, Y. Net ammonification as influenced by plant diversity in experimental grasslands. Soil Biol. Biochem. 2012, 48, 78–87. [Google Scholar] [CrossRef]
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G.W.; Prosser, J.I.; Schuster, S.; Schleper, C. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 2006, 442, 806. [Google Scholar] [CrossRef] [PubMed]
- Sintes, E.; Bergauer, K.; de Corte, D.; Yokokawa, T.; Herndl, G.J. Archaeal amoA gene diversity points to distinct biogeography of ammonia-oxidizing Crenarchaeota in the ocean. Environ. Microbiol. 2013, 15, 1647–1658. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Nicol, G.W.; Neufeld, J.D. Differential responses of soil ammonia-oxidizing Archaea and Bacteria to temperature and depth under two different land uses. Soil Biol. Biochem. 2018, 120, 272–282. [Google Scholar] [CrossRef]
- Merloti, L.F.; Mendes, L.W.; Pedrinho, A.; De Souza, L.F.; Ferrari, B.M.; Tsai, S.M. Forest-to-Agriculture Conversion in Amazon Drives Soil Microbial Communities and N-cycle. Soil Biol. Biochem. 2019, 137, 107567. [Google Scholar] [CrossRef]
- Martínez-Espinosa, C.; Sauvage, S.; Al Bitar, A.; Green, P.; Vörösmarty, C.; Sánchez-Pérez, J. Denitrification in wetlands: A review towards a quantification at global scale. Sci. Total Environ. 2021, 754, 142398. [Google Scholar] [CrossRef] [PubMed]
- Coyne, M.S. Denitrification in soil. In Soil Nitrogen Uses and Environmental Impacts; Lal, R., Steward, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 95–140. [Google Scholar]
- Schulz, S.; Kölbl, A.; Ebli, M.; Buegger, F.; Schloter, M.; Fiedler, S. Field-scale pattern of denitrifying microorganisms and N2O emission rates indiate a high potential for complete denitrification in an agriculturally used organic soil. Microb. Ecol. 2017, 74, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Orellana, L.H.; Rodriguez-R, L.M.; Higgins, S.; Chee-Sanford, J.C.; Sanford, R.A.; Ritalahti, K.M.; Löffler, F.E.; Konstantinidis, K.T. Detecting Nitrous Oxide Reductase (nosZ) Genes in Soil Metagenomes: Method Development and Implications for the Nitrogen Cycle. mBio 2014, 5, e01193-14. [Google Scholar] [CrossRef]
- Nugroho, R.A.; Röling, W.F.M.; Laverman, A.M.; Zoomer, H.R.; Verhoef, H.A. Presence of Nitrosospira cluster 2 bacteria corresponds to N transformation rates in nine acid Scots pine forest soils. FEMS Microbiol. Ecol. 2005, 53, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Mintie, A.T.; Heichen, R.S.; Cromack, K.; Myrold, D.D.; Bottomley, P.J. Ammonia-Oxidizing Bacteria along Meadow-to-Forest Transects in the Oregon Cascade Mountains. Appl. Environ. Microbiol. 2003, 69, 3129–3136. [Google Scholar] [CrossRef]
- Avrahami, S.; Conrad, R. Cold-temperature climate: A factor for selection of ammonia oxidizers in upland soil? Can. J. Microbiol. 2005, 51, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Avrahami, S.; Liesack, W.; Conrad, R. Effects of temperature and fertilizer on activity and community structure of soil ammonia oxidizers. Environ. Microbiol. 2003, 5, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.A.; Bottomley, P.J.; Myrold, D.D. Community composition of ammonia oxidizing bacteria and archaea in soils under stands of red alder and Douglas fir in Oregon. Environ. Microbiol. 2008, 10, 2956. [Google Scholar]
- Boyle, S.A.; Rich, J.J.; Bottomley, P.J.; Cromack, K.; Myrold, D.D. Reciprocal transfer effects on denitrifying community composition and activity at forest and meadow sites in the Cascade Mountains of Oregon. Soil Biol. Biochem. 2006, 38, 870–878. [Google Scholar] [CrossRef]
- Bottomley, P.J.; Taylor, A.E.; Boyle, S.A.; McMahon, S.K.; Rich, J.J.; Cromack Jr, K.; Myrold, D.D. Responses of nitrification and ammonia-oxidizing bacteria to reciprocal transfers of soil between adjacent coniferous forest and meadow vegetation in the Cascade Mountains of Oregon. Microb. Ecol. 2004, 48, 500–508. [Google Scholar] [CrossRef]
- Rich, J.J.; Heichen, R.S.; Bottomley, P.J.; Cromack Jr., K.; Myrold, D.D. Community Composition and Functioning of Denitrifying Bacteria from Adjacent Meadow and Forest Soils. Appl. Environ. Microbiol. 2003, 69, 5974–5982. [Google Scholar] [CrossRef]
- Rachid, C.T.C.C.; Balieiro, F.C.; Peixoto, R.S.; Pinheiro, Y.A.S.; Piccolo, M.C.; Chaer, G.M.; Rosado, A.S. Mixed Plantations Can Promote Microbial Integration and Soil Nitrate Increases with Changes in the N Cycling Genes. Soil Biol. Biochem. 2013, 66, 146–153. [Google Scholar] [CrossRef]
- Balieiro, F.D.C.; Pereira, M.G.; Alves, B.J.R.; Resende, A.S.D.; Franco, A.A. Soil carbon and nitrogen in pasture soil reforested with Eucalyptus and Guachapele. Rev. Bras. Ci. Solo 2008, 32, 1253–1260. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. On the success and failure of mixed-species tree plantations: Lessons learned from a model system of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]
- Khanna, P.K. Comparison of growth and nutrition of young monocultures and mixed stands of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 1997, 94, 105–113. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Bouillet, J.-P.; Gonçalves, J.L.M.; Silva, E.V.; Jourdan, C.; Cunha, M.C.S.; Moreira, M.R.; Saint-André, L.; Maquère, V.; Nouvellon, Y.; et al. Mixed-species plantations of Acacia mangium and Eucalyptus grandis in Brazil. For. Ecol. Manag. 2008, 255, 3905–3917. [Google Scholar] [CrossRef]
- Shibata, H.; Urakawa, R.; Toda, H.; Inagaki, Y.; Tateno, R.; Koba, K.; Nakanishi, A.; Fukuzawa, K.; Yamasaki, A. Changes in nitrogen transformation in forest soil representing the climate gradient of the Japanese archipelago. J. For. Res. 2011, 16, 374–385. [Google Scholar] [CrossRef]
- Jia, Y.; Liao, Z.; Chew, H.; Wang, L.; Lin, B.; Chen, C.; Lu, G.; Lin, Z. Effect of Pennisetum giganteum z.x. lin mixed nitrogen-fixing bacterial fertilizer on the growth, quality, soil fertility and bacterial community of pakchoi (Brassica chinensis L.). PLoS ONE 2020, 15, e0228709. [Google Scholar] [CrossRef]
- Liang, Y.; Pan, F.; He, X.; Chen, X.; Su, Y. Effect of vegetation types on soil arbuscular mycorrhizal fungi and nitrogen-fixing bacterial communities in a karst region. Environ. Sci. Pollut. Res. 2016, 23, 18482–18491. [Google Scholar] [CrossRef]
- Hou, G.; Bi, H.; Wei, X.; Wang, N.; Cui, Y.; Zhao, D.; Ma, X.; Wang, S. Optimal configuration of stand structures in a low-efficiency Robinia pseudoacacia forest based on a comprehensive index of soil and water conservation ecological benefits. Ecol. Indic. 2020, 114, 106308. [Google Scholar] [CrossRef]
- Meng, X.; Fan, S.; Dong, L.; Li, K.; Li, X. Response of Understory Plant Diversity to Soil Physical and Chemical Properties in Urban Forests in Beijing, China. Forests 2023, 14, 571. [Google Scholar] [CrossRef]
- Wang, N.; Bi, H.X.; Cui, Y.H.; Zhao, D.Y.; Hou, G.R.; Yun, H.Y.; Liu, Z.H.; Lan, D.Y.; Jin, C. Optimization of stand structure in Robinia pseudoacacia Linn. based on soil and water conservation improvement function. Ecol. Indic. 2022, 136, 108671. [Google Scholar] [CrossRef]
- Wei, H.; Peng, C.; Liu, X.; Chen, D.; Li, Y.; Wang, M.; Yang, B.; Song, H.; Li, Q.; Jiang, L.; et al. Contrasting Soil Bacterial Community, Diversity, and Function in Two Forests in China. Front. Microbiol. 2018, 9, 1693. [Google Scholar] [CrossRef]
- Wei, X.; Liang, W. Multifactor relationships between stand structure and soil and water conservation functions of Robinia pseudoacacia L. in the Loess Region. PLoS ONE 2019, 14, e0219499. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liang, W.J. Regulation of stand density alters forest structure and soil moisture during afforestation with Robinia pseudoacacia L. and Pinus tabulaeformis Carr. on the Loess Plateau. For. Ecol. Manag. 2021, 491, 119196. [Google Scholar] [CrossRef]
- Bremner, J.M.; Jenkinson, D.S. Determination of organic carbon in soil. J. Soil Sci. 1960, 11, 394–402. [Google Scholar] [CrossRef]
- Li, K.; Zhao, Y.; Yuan, X.; Zhao, H.; Wang, Z.; Li, S.; Malhi, S.S. Comparison of Factors Affecting Soil Nitrate Nitrogen and Ammonium Nitrogen Extraction. Commun. Soil Sci. Plant Anal. 2012, 43, 571–588. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Cornfield, A.H. Ammonia released on treating soils with N sodium hydroxide as a possible means of predicting the nitrogen-supplying power of soils. Nature 1960, 87, 260–261. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate (No. 939); Department of Agriculture Circular: Washington, DC, USA, 1954. [Google Scholar]
- Lu, D.; Li, C.; Sokolwski, E.; Magen, H.; Chen, X.; Wang, H.; Zhou, J. Crop Yield and Soil Available Potassium Changes as Affected by Potassium Rate in Rice–Wheat Systems. Field Crops Res. 2017, 214, 38–44. [Google Scholar] [CrossRef]
- Sparling, G.; West, A. Modifications to the flmigation-extraction technique to permit simultaneous extraction and estimation of soil microbial c and n. Commun. Soil Sci. Plant Anal. 1988, 19, 327–344. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass carbon. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Yao, H.; Gao, Y.; Nicol, G.W.; Campbell, C.D.; Prosser, J.I.; Zhang, L.; Han, W.; Singh, B.K. Links between ammonia oxidizer community structure, abundance, and nitrification potential in acidic soils. Appl. Environ. Microbiol. 2011, 77, 4618–4625. [Google Scholar] [CrossRef]
- Pell, M.; Stenberg, B.; Stenström, J.; Torstensson, L. Potential Denitrification Activity Assay in Soil—With or without Chloramphenicol? Soil Biol. Biochem. 1996, 28, 393–398. [Google Scholar] [CrossRef]
- Muchane, M.N.; Sileshi, G.W.; Gripenberg, S.; Jonsson, M.; Pumarino, L.; Barrios, E. Agroforestry boosts soil health in the humid and sub-humid tropics: A meta-analysis. Agric. Ecosyst. Environ. 2020, 295, 106899. [Google Scholar] [CrossRef]
- Van Eerd, L.L.; Congreves, K.A.; Hayes, A.; Verhallen, A.; Hooker, D.C. Long-term tillage and crop rotation effects on soil quality, organic carbon, and total nitrogen. Can. J. Soil Sci. 2014, 94, 303–315. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, R.; Fu, L.; Tao, S.; Bao, J. Effects of orchard grass on soil fertility and nutritional status of fruit trees in Korla fragrant pear orchard. Horticulturae 2023, 9, 903. [Google Scholar] [CrossRef]
- Cierjacks, A.; Kowarik, I.; Joshi, J.; Hempel, S.; Ristow, M.; Von der Lippe, M.; Weber, E. Biological Flora of the British Isles: Robinia pseudoacacia. J. Ecol. 2013, 101, 1623–1640. [Google Scholar] [CrossRef]
- De Marco, A.; Arena, C.; Giordano, M.; De Santo, A. Impact of the invasive tree black locus ton soil properties of Mediterranean Stone pine-holm oak forests. Plant Soil 2013, 372, 473–486. [Google Scholar] [CrossRef]
- Dancer, W.S.; Peterson, L.A.; Chesters, G. Ammonification and nitrification of N as influenced by soil pH and previous N treatment. Soil Sci. Soc. Am. Proc. 1973, 37, 67–69. [Google Scholar] [CrossRef]
- Lehtovirta-Morley, L.E.; Sayavedra-Soto, L.A.; Gallois, N.; Schouten, S.; Stein, L.Y.; Prosser, J.I.; Nicol, G.W. Identifying Potential Mechanisms Enabling Acidophily in the Ammonia-Oxidizing Archaeon “Candidatus Nitrosotalea devanaterra”. Appl. Environ. Microbiol. 2016, 82, 2608–2619. [Google Scholar] [CrossRef]
- He, J.-Z.; Hu, H.-W.; Zhang, L.-M. Current Insights into the Autotrophic Thaumarchaeal Ammonia Oxidation in Acidic Soils. Soil Biol. Biochem. 2012, 55, 146–154. [Google Scholar] [CrossRef]
- Zhang, K.; Cheng, X.; Dang, H.; Ye, C.; Zhang, Q. Soil nitrogen and denitrification potential as affected by land use and stand age following agricultural abandonment in a headwater catchment. Soil Use Manag. 2012, 28, 361–369. [Google Scholar] [CrossRef]
- Wang, R.; Feng, Q.; Liao, T.; Zheng, X.; Butterbach-Bahl, K.; Zhang, W.; Jin, C. Effects of nitrate concentration on the denitrification potential of a calcic cambisol and its fractions of N2, N2O and NO. Plant Soil 2013, 363, 175–189. [Google Scholar] [CrossRef]
- Li, N.; Shao, T.Y.; Zhu, T.S.; Long, X.H.; Gao, X.M.; Liu, Z.P.; Shao, H.B.; Rengel, Z. Vegetation succession influences soil carbon sequestration in coastal alkali-saline soils in southeast China. Sci. Rep. 2018, 8, 12. [Google Scholar] [CrossRef]
- Dickinson, C.H.; Pugh, G.J.F. Biology of Plant Litter Decomposition; Academic Press: London, UK, 1974; Volume 2, pp. 245–775. [Google Scholar]
- Li, Q.X.; Jia, Z.Q.; Zhu, Y.J.; Wang, Y.S.; Li, H.; Yang, D.F.; Zhao, X.B. Spatial heterogeneity of soil nutrients after the establishment of Caragana intermedia plantation on sand dunes in alpine sandy land of the Tibet Plateau. PLoS ONE 2015, 10, e0124456. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, H.; Li, G.; Ma, W.; Wu, J.; Gong, Y.; Xu, G. Vegetation Degradation Impacts Soil Nutrients and Enzyme Activities in Wet Meadow on the Qinghai-Tibet Plateau. Sci. Rep. 2020, 10, 21271. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, B.; Ma, Y.; Sun, H. Differences in bacterial community structure and potential functions among eucalyptus plantations with different ages and species of trees. Appl. Soil. Ecol. 2020, 149, 103515. [Google Scholar] [CrossRef]
- An, S.-S.; Cheng, Y.; Huang, Y.-M.; Liu, D. Effects of Revegetation on Soil Microbial Biomass, Enzyme Activities, and Nutrient Cycling on the Loess Plateau in China. Restor. Ecol. 2013, 21, 600–607. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil Moisture Is the Major Factor Influencing Microbial Community Structure and Enzyme Activities across Seven Biogeoclimatic Zones in Western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Dai, H.; Dong, B.; Yang, Z.; Yuan, Y.; Tan, Y.; Huang, Y.; Zhang, X. Mixed Plantations Improve Soil Bacterial Similarity by Reducing Heterogeneous Environmental Selection. Forests 2023, 14, 1341. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, W.; Li, X.; Wu, J. A Global Meta-Analysis of the Impacts of Tree Plantations on Biodiversity. Glob. Ecol. Biogeogr. 2022, 31, 576–587. [Google Scholar] [CrossRef]
- Hooper, D.U.; Bignell, D.E.; Brown, V.K.; Brussard, L.; Dangerfield, J.M.; Wall, D.H.; Wardle, D.A.; Coleman, D.C.; Giller, K.E.; Lavelle, P.J.B. Interactions between Aboveground and Belowground Biodiversity in Terrestrial Ecosystems: Patterns, Mechanisms, and Feedbacks. Bioscience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Osler, G.H.R.; Campbell, C.D.; Neilson, R.; Burslem, D.F.R.P.; Van Der Wal, R. The Enigma of Soil Animal Species Diversity Revisited: The Role of Small-Scale Heterogeneity. PLoS ONE 2010, 5, e11567. [Google Scholar] [CrossRef]
- Monard, C.; Vandenkoornhuyse, P.; Bot, B.; Binet, F. Relationship between bacterial diversity and function under biotic control: The soil pesticide degraders as a case study. ISME J. 2011, 5, 1048–1056. [Google Scholar] [CrossRef]
- Nioh, I. Comparison of microflora of Yezo spruce (Picea jezoensis (Sieb. et Zucc.) Carr.) roots in the healthy and deleterious forests. Jpn. J. For. Environ. 1994, 36, 51–56. [Google Scholar]
- Hartman, W.H.; Richardson, C.J.; Vilgalys, R.; Bruland, G.L. Environmental and anthropogenic controls over bacterial communities in wetland soils. Proc. Natl. Acad. Sci. USA 2008, 105, 17842–17847. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.M.; Zhao, M.X.; Cong, J.; Qi, Q.; Xiao, Y.; Cong, W.; Deng, Y.; Zhou, J.Z.; Zhang, Y.G. Soil pH exerts stronger impacts than vegetation type and plant diversity on soil bacterial community composition in subtropical broad-leaved forests. Plant Soil 2020, 450, 273–286. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Abundance and diversity of bacterial, archaeal, and fungal communities along an altitudinal gradient in alpine forest soils: What are the driving factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Sariyildiz, T.; Anderson, J.; Kucuk, M. Effects of tree species and topography on soil chemistry, litter quality, and decomposition in northeast turkey. Soil Biol. Biochem. 2005, 37, 1695–1706. [Google Scholar] [CrossRef]
- Liu, T.; Wu, X.; Li, H.; Alharbi, H.; Wang, J.; Dang, P.; Chen, X.; Kuzyakov, Y.; Yan, W. Soil organic matter, nitrogen and pH driven change in bacterial community following forest conversion. For. Ecol. Manag. 2020, 477, 118473. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Li, Z.A.; Shao, Y.; Xu, G.; Liu, Z.; Zhou, L.; Fu, S. Dicranopteris-dominated understory as major driver of intensive forest ecosystem in humid subtropical and tropical region. Soil Biol. Biochem. 2012, 49, 78–87. [Google Scholar] [CrossRef]
- Feng, Y.; Grogan, P.; Caporaso, J.G.; Zhang, H.; Lin, X.; Knight, R.; Chu, H. PH is a good predictor of the distribution of anoxygenic purple phototrophic bacteria in Arctic soils. Soil Biol. Biochem. 2014, 74, 193–200. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil PH Drives the Spatial Distribution of Bacterial Communities along Elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil ph as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Kaiser, M.; Kleber, M.; Berhe, A.A. How air-drying and rewetting modify soil organic matter characteristics: An assessment to improve data interpretation and inference. Soil Biol. Biochem. 2015, 80, 324–340. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Shen, C.; Ni, Y.; Liang, W.; Wang, J.; Chu, H. Distinct soil bacterial communities along a small-scale elevational gradient in alpine tundra. Front. Microbiol. 2015, 6, 582. [Google Scholar] [CrossRef] [PubMed]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil Water Content and Organic Carbon Availability Are Major Determinants of Soil Microbial Community Composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Armas, C.; Padilla, F.; Pugnaire, F. Climatic change and rainfall patterns: Effects on semi-arid plant communities of the Iberian Southeast. J. Arid. Environ. 2011, 75, 1302–1309. [Google Scholar] [CrossRef]
- Butenschoen, O.; Scheu, S.; Eisenhauer, N. Interactive effects of warming, soil humidity and plant diversity on litter decomposition and microbial activity. Soil Biol. Biochem. 2011, 43, 1902–1907. [Google Scholar] [CrossRef]
- Schnürer, J.; Clarholm, M.; Boström, S.; Rosswall, T. Effects of moisture on soil microorganisms and nematodes: A field experiment. Microb. Ecol. 1986, 12, 217–230. [Google Scholar] [CrossRef]
- Sarathchandra, S.; Ghani, A.; Yeates, G.; Burch, G.; Cox, N. Effect of nitrogen and phosphate fertilisers on microbial and nematode diversity in pasture soils. Soil Biol. Biochem. 2001, 33, 953–964. [Google Scholar] [CrossRef]
- Gu, S.; Hu, Q.; Cheng, Y.; Bai, L.; Liu, Z.; Xiao, W.; Gong, Z.; Wu, Y.; Feng, K.; Deng, Y.; et al. Application of organic fertilizer improves microbial community diversity and alters microbial network structure in tea (Camellia sinensis) plantation soils. Soil Tillage Res. 2019, 195, 104356. [Google Scholar] [CrossRef]
- Soares, M.; Rousk, J. Microbial growth and carbon use efficiency in soil: Links to fungal-bacterial dominance, SOC-quality and stoichiometry. Soil Biol. Biochem. 2019, 131, 195–205. [Google Scholar] [CrossRef]
- Peng, C.; Lai, S.; Luo, X.; Lu, J.; Huang, Q.; Chen, W. Effects of long term rice straw application on the microbial communities of rapeseed rhizosphere in a paddyupland rotation system. Sci. Total Environ. 2016, 557–558, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Torres, I.F.; Bastida, F.; Hernández, T.; Bombach, P.; Richnow, H.H.; García, C. The role of lignin and cellulose in the carbon-cycling of degraded soils under semiarid climate and their relation to microbial biomass. Soil Biol. Biochem. 2014, 75, 152–160. [Google Scholar] [CrossRef]
- He, Z.L.; Yang, X.E.; Baligar, V.C.; Calvert, D.V. Microbiological and biochemical indexing systems for assessing quality of acid soils. Adv. Agron. 2003, 78, 89–138. [Google Scholar]
- Wang, R.; Filley, T.R.; Xu, Z.; Wang, X.; Li, M.H.; Zhang, Y.; Luo, W.; Jiang, Y. Coupled Response of Soil Carbon and Nitrogen Pools and Enzyme Activities to Nitrogen and Water Addition in a Semi-Arid Grassland of Inner Mongolia. Plant Soil 2014, 381, 323–336. [Google Scholar] [CrossRef]
- Deng, J.J.; Yin, Y.; Zhu, W.X.; Zhou, Y.B. Variations in soil bacterial community diversity and structures among different revegetation types in the Baishilazi Nature Reserve. Front. Microbiol. 2018, 9, 2874. [Google Scholar] [CrossRef]
- Sun, H.; Terhonen, E.; Koskinen, K.; Paulin, L.; Kasanen, R.; Asiegbu, F.O. Bacterial diversity and community structure along different peat soils in boreal forest. Appl. Soil Ecol. 2014, 74, 37–45. [Google Scholar] [CrossRef]
- Zeng, Q.; Dong, Y.; An, S. Bacterial community responses to soils along a latitudinal and vegetation gradient on the Loess Plateau, China. PLoS ONE 2016, 11, e0152894. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.; Wang, D.; Heenan, M.; Xu, Z. Short-term responses of soil nitrogen mineralization, nitrification and desnitrification to prescribed burning in a suburban forest ecosystem of subtropical Australia. Sci. Total Environ. 2018, 642, 879–886. [Google Scholar] [CrossRef]
- Wei, X.; Bi, H.; Liang, W.; Hou, G.; Kong, L.; Zhou, Q. Relationship between Soil Characteristics and Stand Structure of Robinia pseudoacacia L. and Pinus tabulaeformis Carr. Mixed Plantations in the Caijiachuan Watershed: An Application of Structural Equation Modeling. Forests 2018, 9, 124. [Google Scholar] [CrossRef]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Zhao, L.; Xu, S.J., Jr.; Liu, Y.Z.; Liu, H.Y.; Cheng, G.D. Soil moisture effect on bacterial and fungal community in Beilu River (Tibetan Plateau) permafrost soils with different vegetation types. J. Appl. Microbiol. 2013, 114, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.A.; Beletsky, A.V.; Rakitin, A.L.; Kadnikov, V.V.; Philippov, D.A.; Mardanov, A.V.; Ravin, N.V.; Dedysh, S.N. Closely Located but Totally Distinct: Highly Contrasting Prokaryotic Diversity Patterns in Raised Bogs and Eutrophic Fens. Microorganisms 2020, 8, 484. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Wang, Y.; Shi, T.; Zhang, X.; Huang, G.; Gong, J. Intertidal zonation affects diversity and functional potentials of bacteria in surface sediments: A case study of the Golden Bay mangrove, China. Appl. Soil Ecol. 2018, 130, 159–168. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Wang, J.; Liu, G.; Wang, G.; Guo, L.; Peng, S. Decreased temporary turnover of bacterial communities along soil depth gradient during a 35-year grazing exclusion period in a semiarid grassland. Geoderma 2019, 351, 49–58. [Google Scholar] [CrossRef]
- Mushinski, R.M.; Zhou, Y.; Gentry, T.J.; Boutton, T.W. Bacterial Metataxonomic Profile and Putative Functional Behavior Associated with C and N Cycle Processes Remain Altered for Decades after Forest Harvest. Soil. Biol. Biochem. 2018, 119, 184–193. [Google Scholar] [CrossRef]
- Chen, L.; Brookes, P.C.; Xu, J.; Zhang, J.; Zhang, C.; Zhou, X.; Luo, Y. Structural and functional differentiation of the root-associated bacterial microbiomes of perennial ryegrass. Soil Biol. Biochem. 2016, 98, 1–10. [Google Scholar]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar]
- Pankratov, T.A.; Ivanova, A.O.; Dedysh, S.; Liesack, W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ. Microbiol. 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- Radajewski, S.; Webster, G.; Reay, D.S.; Morris, S.A.; Ineson, P.; Nedwell, D.B.; Prosser, J.I.; Murrell, J.C. Identification of active methylotroph populations in an acidic forest soil by stable-isotope probing. Microbiology 2002, 148, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.P.; Gischkat, S.; Reiche, M.; Akob, D.M.; Hallberg, K.B.; Kusel, K. Ecophysiology of Fe-cycling bacteria in acidic sediments. Appl. Environ. Microb. 2010, 76, 8174–8183. [Google Scholar] [CrossRef]
- Bryant, D.A.; Costas, A.M.G.; Maresca, J.A.; Chew, A.G.M.; Klatt, C.G.; Bateson, M.M.; Tallon, L.J.; Hostetler, J.; Nelson, W.C.; Heidelberg, J.F.; et al. Candidatus Chloracidobacterium thermophilum: An aerobic phototrophic acidobacterium. Science 2007, 317, 523–526. [Google Scholar] [PubMed]
- Ivanova, A.A.; Zhelezova, A.D.; Chernov, T.I.; Dedysh, S.N. Linking ecology and systematics of acidobacteria: Distinct habitat preferences of the Acidobacteriia and Blastocatellia in tundra soils. PLoS ONE 2020, 15, e0230157. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.; Yang, L.Z.; Zhao, L.P.; Yin, S.X. Actinobacterial community structure in soils receiving long-term organic and inorganic amendments. Appl. Environ. Microbiol. 2008, 74, 526–530. [Google Scholar] [PubMed]
- Ma, B.; Stirling, E.; Liu, Y.; Zhao, K.; Zhou, J.; Singh, B.K.; Tang, C.; Dahlgren, R.A.; Xu, J. Soil Biogeochemical Cycle Couplings Inferred from a Function-Taxon Network. Research 2021, 2021, 7102769. [Google Scholar]
- Singh, B.K.; Munro, S.; Potts, J.M.; Millard, P. Influence of grass species and soil type on rhizosphere microbial community structure in grassland soils. Appl. Soil Ecol. 2007, 36, 147–155. [Google Scholar]
- Pajares, S.; Bohannan, B.J. Ecology of nitrogen fixing, nitrifying, and denitrifying microorganisms in tropical forest soils. Front. Microbiol. 2016, 7, 1045. [Google Scholar] [CrossRef]
- Yousuf, B.; Keshri, J.; Mishra, A.; Jha, B. Application of targeted metagenomics to explore abundance and diversity of CO2-fixing bacterial community using cbbL gene from the rhizosphere of Arachis hypogaea. Gene 2012, 506, 18–24. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.B.; Wu, X.; Huang, T.; Ran, W.; Li, D.C.; Shen, Q.R.; Li, Q.; Zhang, R.F. Swine manure and quicklime have different impacts on chemical properties and composition of bacterial communities of an acidic soil. Appl. Soil Ecol. 2016, 100, 38–44. [Google Scholar] [CrossRef]
- Han, S.; Huang, Q.; Chen, W. Partitioning nitrospira community structure and co-occurrence patterns in a long-term inorganic and organic fertilization soil. J. Soils Sed. 2021, 21, 1099–1108. [Google Scholar] [CrossRef]
- Koch, H.; van Kessel, M.A.H.J.; Lucker, S. Complete nitrification: Insights into the ecophysiology of comammox Nitrospira. Appl. Microbiol. Biotechnol. 2019, 103, 177–189. [Google Scholar] [CrossRef]
- Saghaï, A.; Banjeree, S.; Degrune, F.; Edlinger, A.; García-Palacios, P.; Garland, G.; Heijden, M.G.A.; Herzog, C.; Maestre, F.T.; Pescador, D.S.; et al. Diversity of Archaea and Niche Preferences among Putative Ammonia-oxidizing Nitrososphaeria Dominating across European Arable Soils. Environ. Microbiol. 2022, 24, 341–356. [Google Scholar] [CrossRef]
- Norton, J.M.; Klotz, M.G.; Stein, L.Y.; Arp, D.J.; Bottomley, P.J.; Chain, P.S.; Hauser, L.J.; Land, M.L.; Larimer, F.W.; Shin, M.W.; et al. Complete genome sequence of Nitrosospira multiformis, an ammonia-oxidizing bacterium from the soil environment. Appl. Environ. Microbiol. 2008, 74, 3559–3572. [Google Scholar] [CrossRef]
- Castellano-Hinojosa, A.; González-López, J.; Bedmar, E.J. Distinct effect of nitrogen fertilisation and soil depth on nitrous oxide emissions and nitrifiers and denitrifiers abundance. Biol. Fertil. Soils 2018, 54, 829–840. [Google Scholar] [CrossRef]
- Mushinski, R.M.; Gentry, T.J.; Dorosky, R.J.; Boutton, T. Forest harvest intensity and soil depth alter inorganic nitrogen pool sizes and ammonia oxidizer community composition. Soil Boil. Biochem. 2017, 112, 216–227. [Google Scholar] [CrossRef]
- Koops, H.-P.; Purkhold, U.; Pommerening-Röser, A.; Timmermann, G.; Wagner, M. The lithoautotrophic ammonia-oxidizing bacteria. In The Prokaryotes; Springer: Berlin, Germany, 2006; pp. 778–811. [Google Scholar]
- Braker, G.; Zhou, J.; Wu, L.; Devol, A.H.; Tiedje, J.M. Nitrite reductase genes (nirK and nirS) as functional markers to investigate diversity of denitrifying bacteria in Pacific Northwest marine sediment communities. Appl. Environ. Microbiol. 2000, 66, 2096–2104. [Google Scholar] [CrossRef]
- Jones, C.M.; Hallin, S. Ecological and evolutionary factors underlying global and local assembly of denitrifier communities. ISME J. 2010, 4, 633–641. [Google Scholar] [CrossRef]
- Cavigelli, M.A.; Robertson, G.P. The Functional Significance of Denitrifier Community Composition in a Terrestrial Ecosystem. Ecology 2000, 81, 1402–1414. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C.; Wang, W.; Hughes, J.M.; Lewis, T.; Hou, E.; Shen, J. Vertical distribution of soil denitrifying communities in a wet sclerophyll forest under long-term repeated burning. Microb. Ecol. 2015, 70, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, X.; Li, X.; Li, X.; Wang, J.; Zhang, H. Changes of Microbial Population and N-Cycling Function Genes with Depth in Three Chinese Paddy Soils. PLoS ONE 2017, 12, e0189506. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Wang, S.; Zhang, J.; Cai, Z. Effects of soil moisture on gross N transformations and N2O emission in acid subtropical forest soils. Biol. Fertil. Soils 2014, 50, 1099–1108. [Google Scholar] [CrossRef]
- Szukics, U.; Abell, G.C.J.; Hödl, V.; Mitter, B.; Sessitsch, A.; Hackl, E.; Zechmeister-Boltenstern, S. Nitrifiers and denitrifiers respond rapidly to changed moisture and increasing temperature in a pristine forest soil. FEMS Microbiol. Ecol. 2010, 72, 395–406. [Google Scholar] [CrossRef]
- Ribbons, R.R.; Levy-Booth, D.J.; Masse, J.; Grayston, S.J.; McDonald, M.A.; Vesterdal, L.; Prescott, C.E. Linking microbial communities, functional genes and nitrogen-cycling processes in forest floors under four tree species. Soil Biol. Biochem. 2016, 103, 181–191. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.R.; Huang, Y.T.; Fu, S.L.; Wang, J.X.; Ming, A.G.; Li, X.Z.; Yao, M.J.; Li, H. Tree species mixture inhibits soil organic carbon mineralization accompanied by decreased r-selected bacteria. Plant Soil 2018, 431, 203–216. [Google Scholar] [CrossRef]
- Sun, X.; Han, X.; Ping, F.; Zhang, L.; Zhang, K.; Chen, M.; Wu, W. Effect of rice-straw biochar on nitrous oxide emissions from paddy soils under elevated CO2 and temperature. Sci. Total. Environ. 2018, 628, 1009–1016. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Primer Sequence (5′-3′) | Real-Time Fluorescent Quantitative PCR Reaction Conditions |
|---|---|---|---|
| AOB amoA | amoA-1F | GGGGTTTCTACTGGTGGT | 95 °C/30 s; 40 cycles of 95 °C/5 s, 58 °C/34 s |
| amoA-2R | CCCCTCKGSAAAGCCTTCTTC | ||
| nirS | Cd3AF | GTSAACGTSAAGGARACSGG | 95 °C/30 s; 45 cycles of 95 °C/5 s, 60 °C/40 s |
| R3cd | GASTTCGGRTGSGTCTTGA | ||
| nirK | 1F | GGMATGGTKCCSTGGCA | 95 °C/30 s; 40 cycles of 95 °C/5 s, 60 °C/40 s |
| 5R | GCCTCGATCAGRTTRTGGTT |
| Plantation | Soil Layer /(cm) | SMC /(%) | pH | SOM /(g/kg) | TN /(g/kg) | HN/ (mg/kg) | NH4+-N /(mg/kg) | NO3--N /(mg/kg) | AK /(mg/kg) | AP /(mg/kg) | MBC /(mg/kg) | MBN /(mg/kg) | PNR /(mg/kg/d) | PDR /(mg/kg/d) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R | 0–5 | 19.91 ± 3.65 Aa | 7.04 ± 0.21 Ba | 78.44 ± 17.54 Aab | 5.24 ± 0.97 Aa | 176.64 ± 17.16 Aa | 12.27 ± 0.81 Aa | 54.65 ± 15.44 Aa | 210.57 ± 19.60 Aa | 5.30 ± 0.92 Aa | 298.61 ± 36.85 Aa | 39.51 ± 9.28 Aa | 12.07 ± 6.99 Aa | 60.11 ± 9.58 Aa |
| 5–10 | 15.57 ± 1.75 Ba | 7.80 ± 0.10 Aa | 28.01 ± 9.10 Ba | 2.13 ± 0.55 Ba | 79.45 ± 13.94 Ba | 9.75 ± 0.53 Bab | 25.81 ± 5.19 Ba | 132.92 ± 8.89 Ba | 2.16 ± 0.52 Ba | 126.04 ± 37.75 Ba | 21.39 ± 2.99 Ba | 6.16 ± 2.65 Aa | 36.26 ± 8.46 Ba | |
| Q | 0–5 | 12.38 ± 1.03 Ab | 6.50 ± 0.40 Bb | 66.51 ± 15.66 Ab | 4.11 ± 0.55 Ab | 123.10 ± 17.70 Ab | 12.55 ± 0.85 Aa | 26.68 ± 3.74 Ab | 137.69 ± 11.22 Ab | 3.49 ± 0.17 Ab | 138.45 ± 35.80 Ab | 15.63 ± 1.51 Ab | 0.76 ± 0.86 Ab | 26.07 ± 7.13 Ab |
| 5–10 | 9.48 ± 0.81 Bb | 7.32 ± 0.30 Ab | 18.18 ± 3.73 Bb | 1.61 ± 0.25 Ba | 48.25 ± 3.41 Bb | 9.61 ± 0.35 Bb | 13.73 ± 1.04 Bb | 57.62 ± 4.63 Bb | 1.21 ± 0.16 Bb | 82.54 ± 15.82 Bb | 12.39 ± 3.95 Ab | 0.05 ± 0.03 Ab | 12.97 ± 2.44 Bb | |
| RQ | 0–5 | 12.43 ± 0.78 Ab | 6.19 ± 0.42 Bb | 91.58 ± 13.43 Aa | 5.61 ± 0.69 Aa | 140.10 ± 38.19 Ab | 11.79 ± 0.84 Aa | 36.19 ± 0.65 Ab | 130.37 ± 15.30 Ab | 4.91 ± 0.76 Aa | 45.55 ± 4.36 Bc | 9.30 ± 3.14 Ab | 0.25 ± 0.13 Ab | 27.30 ± 5.01 Ab |
| 5–10 | 10.16 ± 0.64 Bb | 6.88 ± 0.37 Ac | 26.37 ± 4.08 Bab | 2.07 ± 0.28 Ba | 44.70 ± 11.35 Bb | 10.18 ± 0.65 Ba | 15.39 ± 2.36 Bb | 72.03 ± 12.08 Bc | 2.25 ± 0.18 Ba | 56.26 ± 8.15 Ab | 9.86 ± 3.67 Ab | 0.09 ± 0.07 Bb | 15.27 ± 5.62 Bb |
| Sample | Shannon | Simpson | Chao1 | ACE |
|---|---|---|---|---|
| R5 | 9.91 ± 0.09 Aa | 0.997 ± 0.00 Ba | 3641.70 ± 232.99 Aa | 3753.28 ± 253.81 Aa |
| Q5 | 9.44 ± 0.47 Ab | 0.995 ± 0.00 Ab | 3079.28 ± 549.90 Aa | 3139.68 ± 549.43 Ab |
| RQ5 | 9.59 ± 0.30 Aab | 0.996 ± 0.00 Aab | 3300.94 ± 409.77 Aa | 3407.28 ± 424.25 Aab |
| R10 | 9.97 ± 0.23 Aa | 0.998 ± 0.00 Aa | 3585.26 ± 347.20 Aa | 3673.87 ± 321.55 Aa |
| Q10 | 9.70 ± 0.29 Aa | 0.996 ± 0.00 Ab | 3562.25 ± 481.29 Aa | 3591.92 ± 453.83 Aa |
| RQ10 | 9.23 ± 0.32 Ab | 0.995 ± 0.00 Ab | 2823.21 ± 507.57 Ab | 2819.65 ± 489.59 Ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Chen, J.; Zheng, Y.; Jiang, R.; Sang, Y.; Zhang, J. The Effects of Mixed Robinia pseudoacacia and Quercus variabilis Plantation on Soil Bacterial Community Structure and Nitrogen-Cycling Gene Abundance in the Southern Taihang Mountain Foothills. Microorganisms 2024, 12, 1773. https://doi.org/10.3390/microorganisms12091773
Yang Y, Chen J, Zheng Y, Jiang R, Sang Y, Zhang J. The Effects of Mixed Robinia pseudoacacia and Quercus variabilis Plantation on Soil Bacterial Community Structure and Nitrogen-Cycling Gene Abundance in the Southern Taihang Mountain Foothills. Microorganisms. 2024; 12(9):1773. https://doi.org/10.3390/microorganisms12091773
Chicago/Turabian StyleYang, Yi, Jing Chen, Yiwei Zheng, Rui Jiang, Yuqiang Sang, and Jinsong Zhang. 2024. "The Effects of Mixed Robinia pseudoacacia and Quercus variabilis Plantation on Soil Bacterial Community Structure and Nitrogen-Cycling Gene Abundance in the Southern Taihang Mountain Foothills" Microorganisms 12, no. 9: 1773. https://doi.org/10.3390/microorganisms12091773