

Kupffer Cells and Hepatocytes: A Key Relation in the Context of Canine Leishmaniasis

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
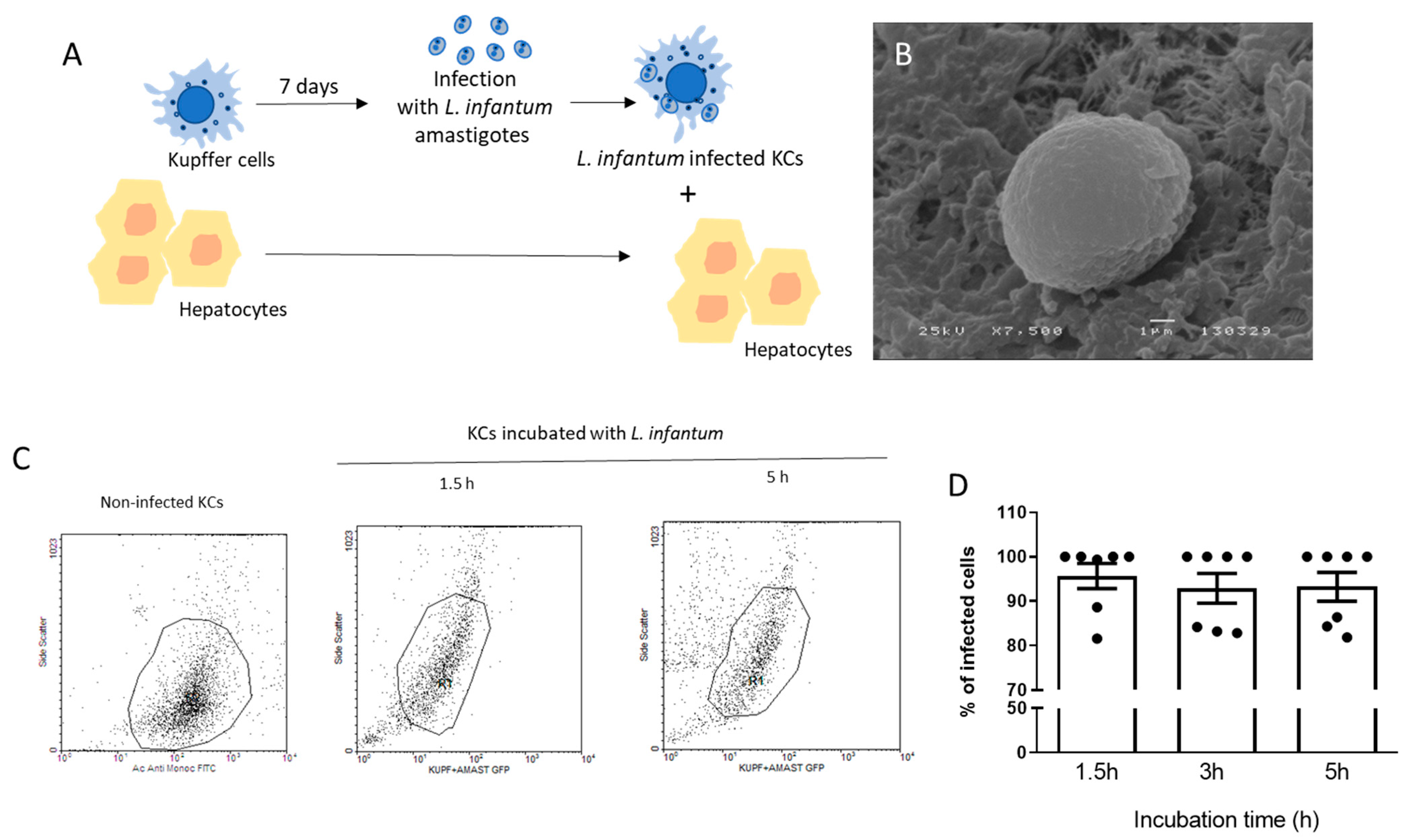
2.1. Isolation of Kupffer Cells and Hepatocytes
2.2. Parasites and Kupffer Cells Infection
2.3. Flow Cytometry
2.4. Scan Electron Microscopy (SEM)
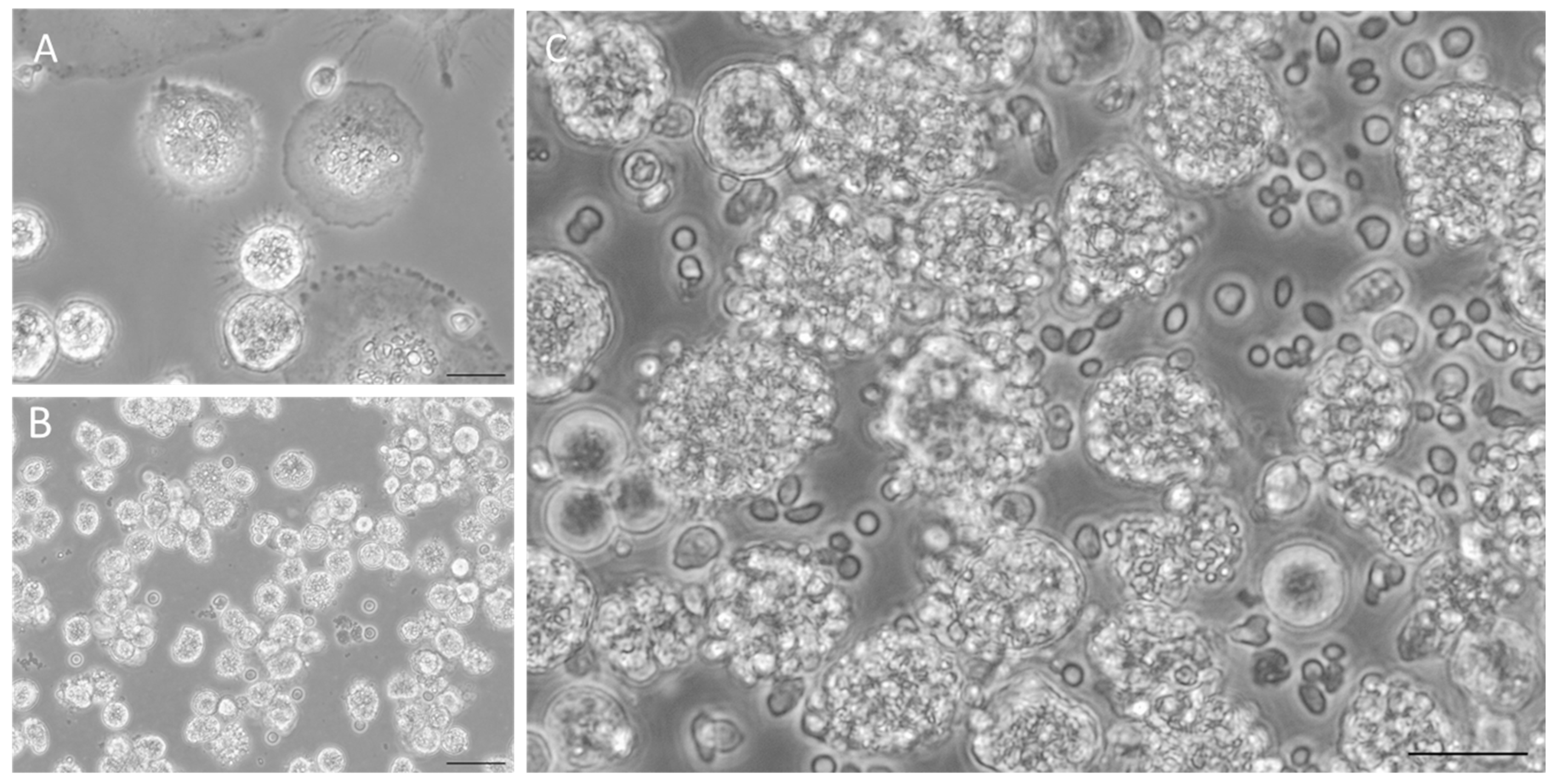
2.5. Co-Culture System
2.6. Real-Time PCR
2.7. Statistical Analysis
2.8. Ethical Considerations
3. Results and Discussion
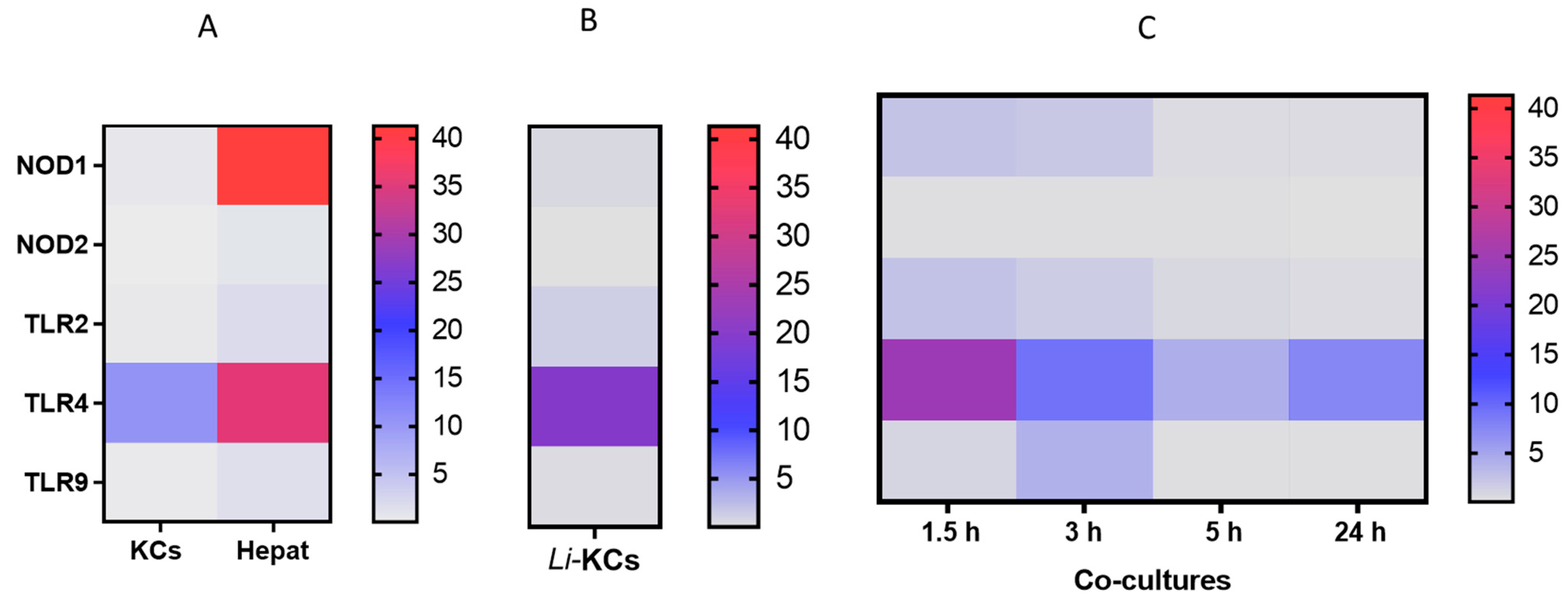
3.1. Co-Cultures Do Not Extensively Activate PRRs
3.2. Co-Cultures Can Generate Key Cytokines and Orchestrate an Immune Response
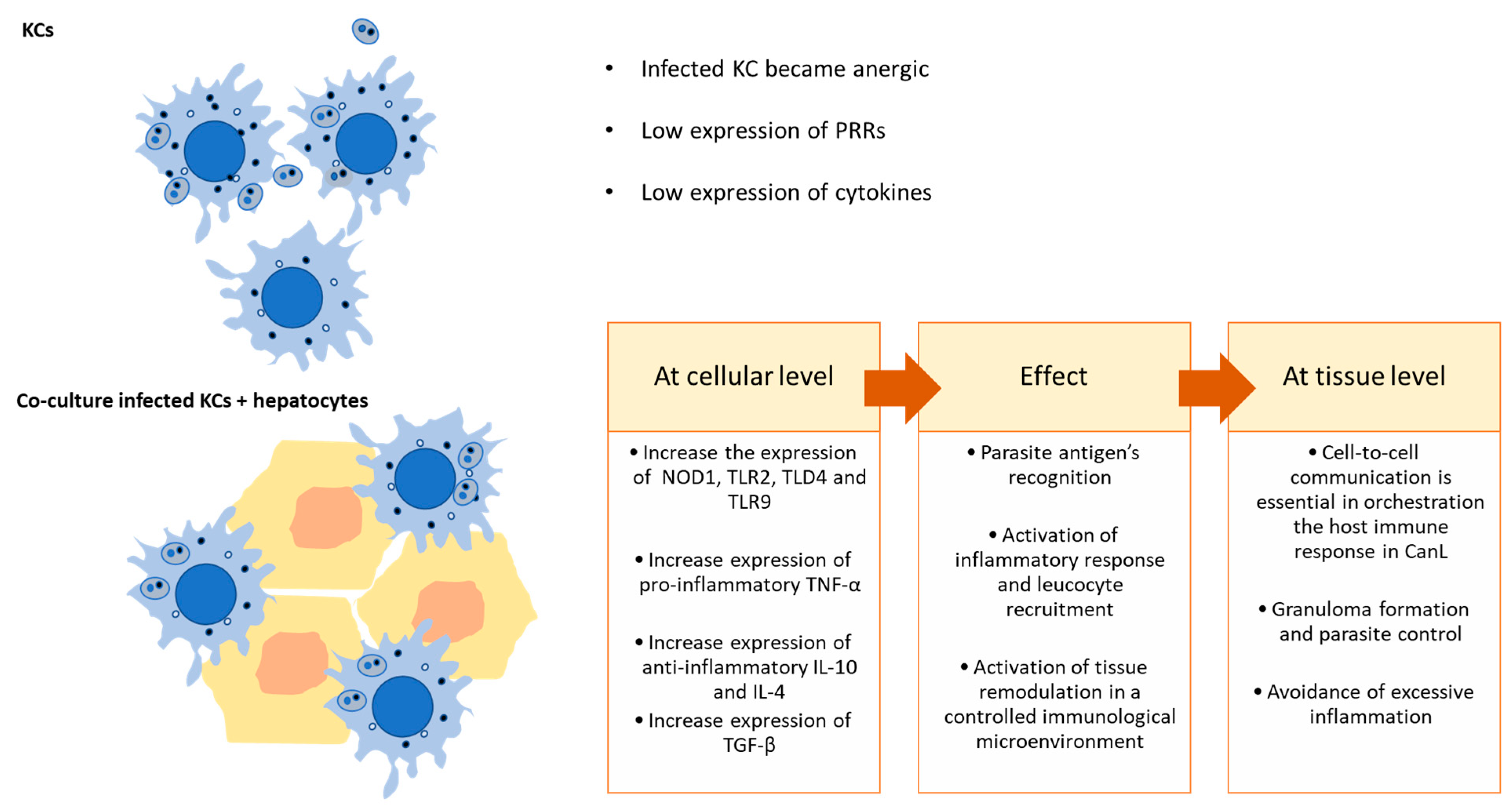
3.3. The Interaction of L. infantum-Infected KCs with Hepatocytes Is Key in Regulating Local Immune Response
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, B.M.; Maia, C.; Courtenay, O.; Llabrés-Brustenga, A.; Lotto Batista, M.; Moirano, G.; van Daalen, K.R.; Semenza, J.C.; Lowe, R. A climatic suitability indicator to support Leishmania infantum surveillance in Europe: A modelling study. Lancet Reg. Health-Eur. 2024, 43, 100971. [Google Scholar] [CrossRef] [PubMed]
- Palatnik-De-Sousa, C.B.; Day, M.J. One Health: The global challenge of epidemic and endemic leishmaniasis. Parasites Vectors 2011, 4, 197. [Google Scholar] [CrossRef] [PubMed]
- TDR|Neglected Tropical Disease (NTD) Research. Visceral Leishmaniasis Research. 2024. Available online: https://tdr.who.int/our-work/research-for-implementation/neglected-tropical-diseases-research/visceral-leishmaniasis-research (accessed on 6 September 2024).
- Vilas-Boas, D.F.; Nakasone, E.K.N.; Gonçalves, A.A.M.; Lair, D.F.; Oliveira, D.S.d.; Pereira, D.F.S.; Silva, G.G.; Conrado, I.d.S.S.; Resende, L.A.; Zaldívar, M.F.; et al. Global Distribution of Canine Visceral Leishmaniasis and the Role of the Dog in the Epidemiology of the Disease. Pathogens 2024, 13, 455. [Google Scholar] [CrossRef]
- Paz, S.; Majeed, A.; Christophides, G.K. Climate change impacts on infectious diseases in the Eastern Mediterranean and the Middle East (EMME)—Risks and recommendations. Clim. Chang. 2021, 169, 40. [Google Scholar] [CrossRef]
- Arikan, A.; Cakir, N. Climate change and future infectious diseases: A growing threat. New Microbes New Infect. 2023, 52, 101088. [Google Scholar] [CrossRef]
- Alexandre-Pires, G.; de Brito, M.T.; Algueró, C.; Martins, C.; Rodrigues, O.R.; da Fonseca, I.P.; Santos-Gomes, G. Canine leishmaniasis. Immunophenotypic profile of leukocytes in different compartments of symptomatic, asymptomatic and treated dogs. Vet. Immunol. Immunopathol. 2010, 137, 275–283. [Google Scholar] [CrossRef]
- Sánchez, M.A.; Diaz, N.L.; Zerpa, O.; Negron, E.; Convit, J.; Tapia, F.J. Organ-specific immunity in canine visceral leishmaniasis: Analysis of symptomatic and asymptomatic dogs naturally infected with Leishmania chagasi. Am. J. Trop. Med. Hyg. 2004, 70, 618–624. [Google Scholar] [CrossRef]
- Gao, B. Basic liver immunology. Cell. Mol. Immunol. 2016, 13, 265–266. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell. Mol. Immunol. 2016, 13, 301–315. [Google Scholar]
- Gong, J.; Tu, W.; Liu, J.; Tian, D. Hepatocytes: A key role in liver inflammation. Front. Immunol. 2022, 13, 1083780. [Google Scholar] [CrossRef] [PubMed]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Musrati, M.A.; De Baetselier, P.; Movahedi, K.; Van Ginderachter, J.A. Ontogeny, functions and reprogramming of Kupffer cells upon infectious disease. Front. Immunol. 2023, 14, 1238452. [Google Scholar] [CrossRef]
- Rodrigues, A.; Santos-Mateus, D.; Alexandre-Pires, G.; Valério-Bolas, A.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; de Jesus, J.; Alves-Azevedo, R.; Lopes-Ventura, S.; et al. Leishmania infantum exerts immunomodulation in canine Kupffer cells reverted by meglumine antimoniate. Comp. Immunol. Microbiol. Infect. Dis. 2017, 55, 42–52. [Google Scholar] [CrossRef]
- Rodrigues, A.; Alexandre-Pires, G.; Valério-Bolas, A.; Santos-Mateus, D.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; Nunes, T.; Alves-Azevedo, R.; Lopes-Ventura, S.; et al. Dog hepatocytes are key effector cells in the liver innate immune response to Leishmania infantum. Parasitology 2019, 146, 753–764. [Google Scholar] [CrossRef]
- Rodrigues, A.V.; Alexandre-Pires, G.; Valério-Bolas, A.; Santos-Mateus, D.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; Nunes, T.; Alves-Azevedo, R.; Santos, M.; et al. 3D-Hepatocyte Culture Applied to Parasitology: Immune Activation of Canine Hepatic Spheroids Exposed to Leishmania Infantum. Biomedicines 2020, 8, 628. [Google Scholar] [CrossRef]
- Rodrigues, A.V.; Valério-Bolas, A.; Alexandre-Pires, G.; Aires-Pereira, M.; Nunes, T.; Ligeiro, D.; Pereira da Fonseca, I.; Santos-Gomes, G. Zoonotic Visceral Leishmaniasis: New Insights on Innate Immune Response by Blood Macrophages and Liver Kupffer Cells to Leishmania infantum Parasites. Biology 2022, 11, 100. [Google Scholar] [CrossRef]
- Santos-Gomes, G.M.; Abranches, P. Comparative study of infectivity caused by promastigotes of Leishmania infantum MON-1, L. infantum MON-24 and L. donovani MON-18. Folia Parasitol. 1996, 43, 7–12. [Google Scholar]
- Rodrigues, O.R.; Moura, R.A.; Gomes-Pereira, S.; Santos-Gomes, G.M. H-2 complex influences cytokine gene expression in Leishmania infantum-infected macrophages. Cell. Immunol. 2006, 243, 118–126. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, R. Toll-like receptors in acute liver injury and regeneration. Int. Immunopharmacol. 2011, 11, 1433–1441. [Google Scholar] [CrossRef]
- Kesar, V.; Odin, J.A. Toll-like receptors and liver disease. Liver Int. 2014, 34, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Jeong, W.-I.; Tian, Z. Liver: An organ with predominant innate immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, N.; Kanai, T. Role of Toll-like receptors in immune activation and tolerance in the liver. Front. Immunol. 2014, 5, 221. [Google Scholar] [CrossRef]
- Kiziltas, S. Toll-like receptors in pathophysiology of liver diseases. World J. Hepatol. 2016, 8, 1354–1369. [Google Scholar] [CrossRef]
- Murray, H.W. Tissue granuloma structure-function in experimental visceral leishmaniasis. Int. J. Exp. Pathol. 2001, 82, 249–267. [Google Scholar] [CrossRef]
- Kaye, P.M.; Beattie, L. Lessons from other diseases: Granulomatous inflammation in leishmaniasis. Semin. Immunopathol. 2016, 38, 249–260. [Google Scholar] [CrossRef]
- Fischer, L.; Lucendo-villarin, B.; Hay, D.C.; O’farrelly, C. Human PSC-Derived Hepatocytes Express Low Levels of Viral Pathogen Recognition Receptors, but Are Capable of Mounting an Effective Innate Immune Response. Int. J. Mol. Sci. 2020, 21, 3831. [Google Scholar] [CrossRef]
- Anand, S.; Wang, P.; Yoshimura, K.; Choi, I.H.; Hilliard, A.; Chen, Y.H.; Wang, C.-R.; Schulick, R.; Flies, A.S.; Flies, D.B.; et al. Essential role of TNF family molecule LIGHT as a cytokine in the pathogenesis of hepatitis. J. Clin. Investig. 2006, 116, 1045–1051. [Google Scholar] [CrossRef]
- Osero, B.O.; Aruleba, R.T.; Brombacher, F.; Hurdayal, R. Unravelling the unsolved paradoxes of cytokine families in host resistance and susceptibility to Leishmania infection. Cytokine X 2020, 2, 100043. [Google Scholar] [CrossRef]
- O’Garra, A.; Vieira, P.L.; Vieira, P.; Goldfeld, A.E. IL-10–producing and naturally occurring CD4+ Tregs: Limiting collateral damage. J. Clin. Investig. 2004, 114, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Wang, X.Z. Interleukin-10 and chronic liver disease. World J. Gastroenterol. 2006, 12, 1681–1685. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, Y.; Huang, R.; Chen, Z.; Wang, X.; Chen, F.; Huang, Y. Interleukin-10 gene intervention ameliorates liver fibrosis by enhancing the immune function of natural killer cells in liver tissue. Int. Immunopharmacol. 2024, 127, 111341. [Google Scholar] [CrossRef] [PubMed]
- Poniatowski, L.A.; Wojdasiewicz, P.; Gasik, R.; Szukiewicz, D. Transforming Growth Factor Beta Family: Insight into the Role of Growth Factors in Regulation of Fracture Healing Biology and Potential Clinical Applications. Mediat. Inflamm. 2015, 2015, 137823. [Google Scholar] [CrossRef]
- Lehn, M.; Kandil, O.; Arena, C.; Rein, M.S.; Remold, H.G. Interleukin-4 fails to inhibit interferon-γ-induced activation of human colostral macrophages. Cell. Immunol. 1992, 141, 233–242. [Google Scholar] [CrossRef]
- Ho, J.L.; He, S.H.; Jose, M.C.R.; Wick, E.A. Interleukin-4 Inhibits Human Macrophage Activation by Tumor Necrosis Factor, Granulocyte-Monocyte Colony-Stimulating Factor, and Interleukin-3 for Antileishmanial Activity and Oxidative Burst Capacity. J. Infect. Dis. 1992, 165, 344–351. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, A.; Alexandre-Pires, G.; Valério-Bolas, A.; Nunes, T.; Pereira da Fonseca, I.; Santos-Gomes, G. Kupffer Cells and Hepatocytes: A Key Relation in the Context of Canine Leishmaniasis. Microorganisms 2024, 12, 1887. https://doi.org/10.3390/microorganisms12091887
Rodrigues A, Alexandre-Pires G, Valério-Bolas A, Nunes T, Pereira da Fonseca I, Santos-Gomes G. Kupffer Cells and Hepatocytes: A Key Relation in the Context of Canine Leishmaniasis. Microorganisms. 2024; 12(9):1887. https://doi.org/10.3390/microorganisms12091887
Chicago/Turabian StyleRodrigues, Armanda, Graça Alexandre-Pires, Ana Valério-Bolas, Telmo Nunes, Isabel Pereira da Fonseca, and Gabriela Santos-Gomes. 2024. "Kupffer Cells and Hepatocytes: A Key Relation in the Context of Canine Leishmaniasis" Microorganisms 12, no. 9: 1887. https://doi.org/10.3390/microorganisms12091887
APA StyleRodrigues, A., Alexandre-Pires, G., Valério-Bolas, A., Nunes, T., Pereira da Fonseca, I., & Santos-Gomes, G. (2024). Kupffer Cells and Hepatocytes: A Key Relation in the Context of Canine Leishmaniasis. Microorganisms, 12(9), 1887. https://doi.org/10.3390/microorganisms12091887