
Isolation and Characterization of Colistin-Resistant Enterobacteriaceae from Foods in Two Italian Regions in the South of Italy

 , , , , , ,
, , , , , ,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Bacterial Strains
2.2. Colistin-Resistant Screening
2.3. Whole Genome Sequencing (WGS)
2.4. Antimicrobial Susceptibility Test
2.5. Nucleotide Sequence Accession Numbers
2.6. Statistical Analysis
3. Results
3.1. Isolation of Colistin Resistance Bacteria
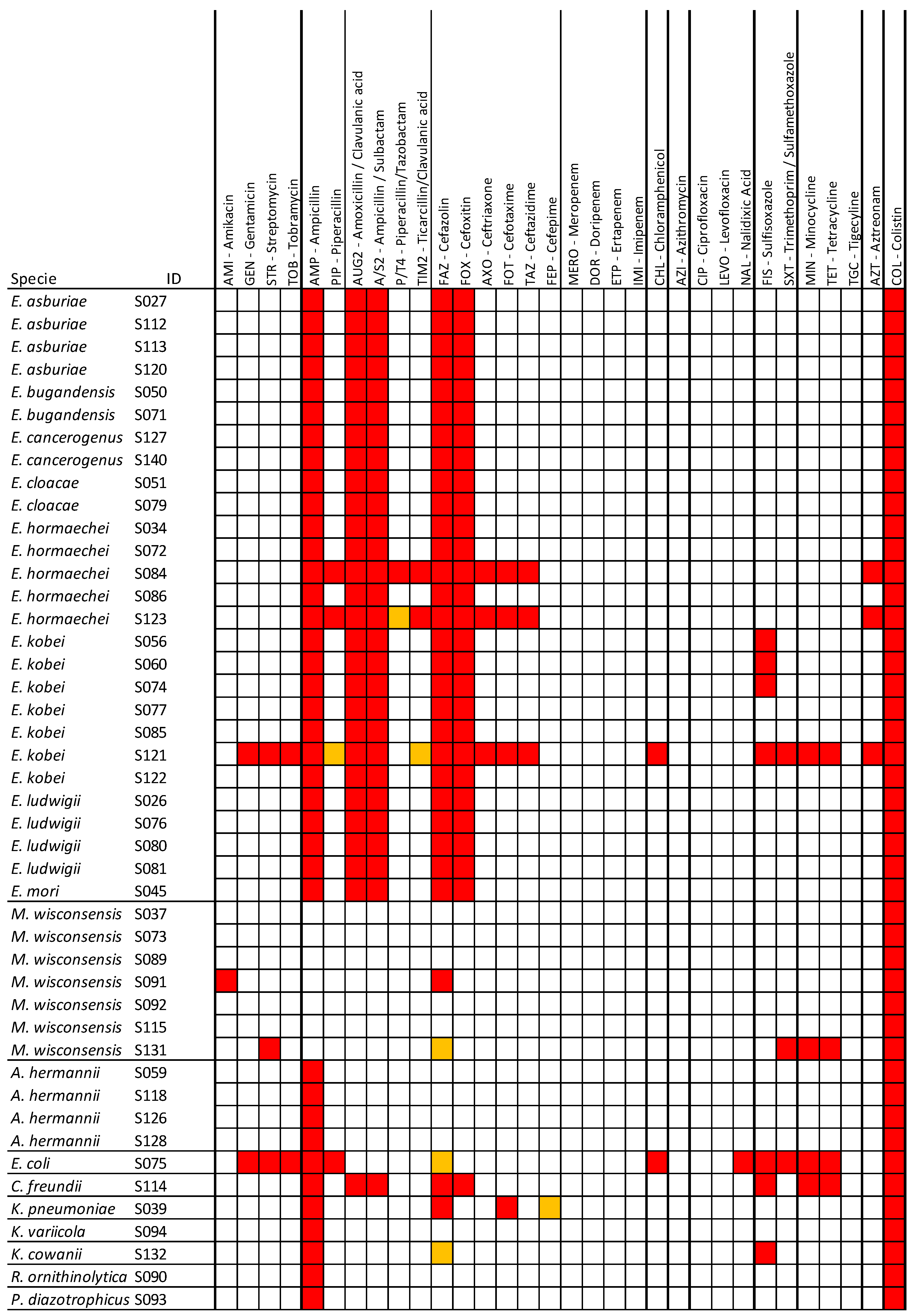
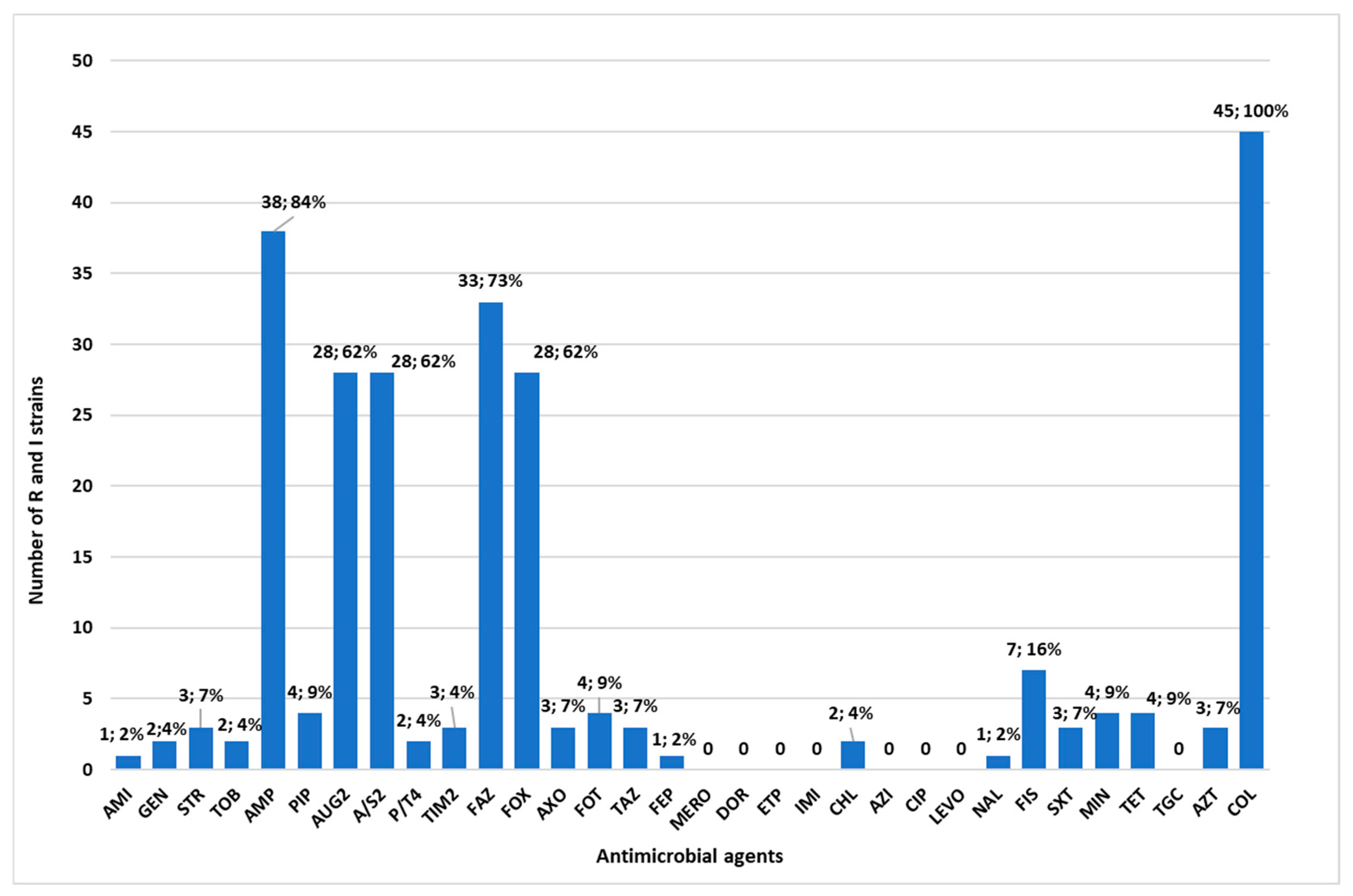
3.2. Antimicrobial Resistance Profiles of the Isolated Bacteria
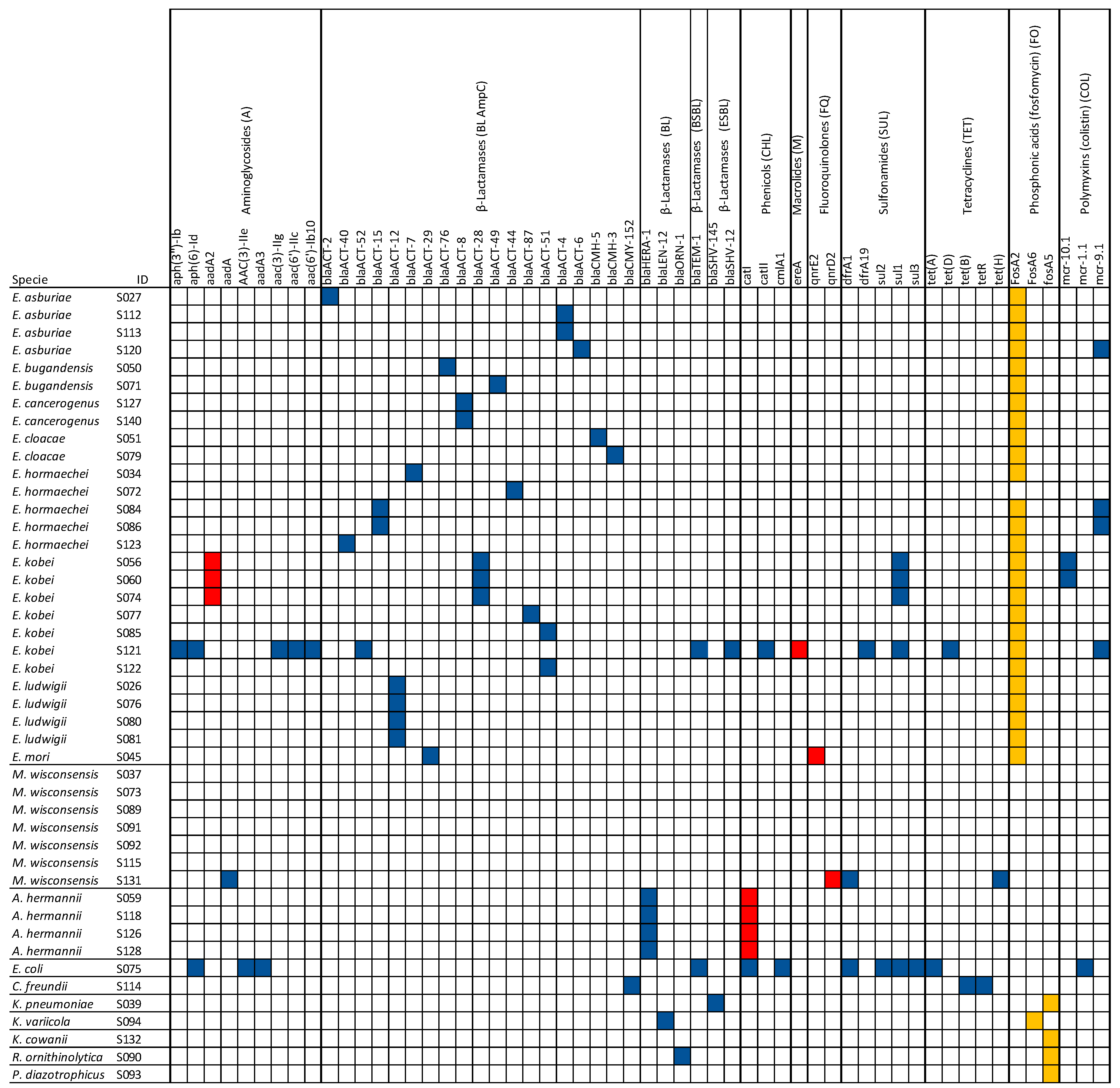
3.3. Detection of AMR, Virulence, and Plasmid Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Echols, R.M.; Tosiello, R.L.; Haverstock, D.C.; Tice, A.D. Demographic, Clinical, and Treatment Parameters Influencing the Outcome of Acute Cystitis. Clin. Infect. Dis. 1999, 29, 113–119. [Google Scholar] [CrossRef]
- Czaja, C.A.; Scholes, D.; Hooton, T.M.; Stamm, W.E. Population-Based Epidemiologic Analysis of Acute Pyelonephritis. Clin. Infect. Dis. 2007, 45, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasan, M.N.; Eckel-Passow, J.E.; Baddour, L.M. Impact of Healthcare-Associated Acquisition on Community-Onset Gram-Negative Bloodstream Infection: A Population-Based Study: Healthcare-Associated Gram-Negative BSI. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Sharma, L.; Dela Cruz, C.S.; Zhang, D. Clinical Epidemiology, Risk Factors, and Control Strategies of Klebsiella Pneumoniae Infection. Front. Microbiol. 2021, 12, 750662. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Kim, J.-H.; Lee, Y. Virulence Factors Associated With Escherichia Coli Bacteremia and Urinary Tract Infection. Ann. Lab. Med. 2022, 42, 203–212. [Google Scholar] [CrossRef] [PubMed]
- van Duijn, P.J.; Dautzenberg, M.J.D.; Oostdijk, E.A.N. Recent Trends in Antibiotic Resistance in European ICUs. Curr. Opin. Crit. Care 2011, 17, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Bouchillon, S.K.; Badal, R.E.; Hoban, D.J.; Hawser, S.P. Antimicrobial Susceptibility of Inpatient Urinary Tract Isolates of Gram-Negative Bacilli in the United States: Results from the Study for Monitoring Antimicrobial Resistance Trends (SMART) Program: 2009−2011. Clin. Ther. 2013, 35, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Thaden, J.T.; Fowler, V.G.; Sexton, D.J.; Anderson, D.J. Increasing Incidence of Extended-Spectrum β-Lactamase-Producing Escherichia Coli in Community Hospitals throughout the Southeastern United States. Infect. Control. Hosp. Epidemiol. 2016, 37, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Ojer-Usoz, E.; González, D.; Vitas, A. Clonal Diversity of ESBL-Producing Escherichia Coli Isolated from Environmental, Human and Food Samples. Int. J. Environ. Res. Public Health 2017, 14, 676. [Google Scholar] [CrossRef] [PubMed]
- Bourdichon, F.; Betts, R.; Dufour, C.; Fanning, S.; Farber, J.; McClure, P.; Stavropoulou, D.A.; Wemmenhove, E.; Zwietering, M.H.; Winkler, A. Processing Environment Monitoring in Low Moisture Food Production Facilities: Are We Looking for the Right Microorganisms? Int. J. Food Microbiol. 2021, 356, 109351. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2019–2020. EFSA J. 2022, 20, e07209. [Google Scholar] [CrossRef]
- Barlaam, A.; Parisi, A.; Spinelli, E.; Caruso, M.; Taranto, P.D.; Normanno, G. Global Emergence of Colistin-Resistant Escherichia Coli in Food Chains and Associated Food Safety Implications: A Review. J. Food Prot. 2019, 82, 1440–1448. [Google Scholar] [CrossRef]
- World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151552-8. [Google Scholar]
- Lay, K.K.; Jeamsripong, S.; Sunn, K.P.; Angkititrakul, S.; Prathan, R.; Srisanga, S.; Chuanchuen, R. Colistin Resistance and ESBL Production in Salmonella and Escherichia Coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area. Antibiotics 2021, 10, 657. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a Novel Plasmid-Mediated Colistin-Resistance Gene, Mcr-2, in Escherichia Coli, Belgium, June 2016. Eurosurveillance 2016, 21, 30280. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel Plasmid-Mediated Colistin Resistance Gene Mcr-3 in Escherichia Coli. mBio 2017, 8, e00543-17. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel Plasmid-Mediated Colistin Resistance Mcr-4 Gene in Salmonella and Escherichia Coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a Novel Transposon-Associated Phosphoethanolamine Transferase Gene, Mcr-5, Conferring Colistin Resistance in d-Tartrate Fermenting Salmonella Enterica Subsp. Enterica Serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [PubMed]
- AbuOun, M.; Stubberfield, E.J.; Duggett, N.A.; Kirchner, M.; Dormer, L.; Nunez-Garcia, J.; Randall, L.P.; Lemma, F.; Crook, D.W.; Teale, C.; et al. Mcr-1 and Mcr-2 (Mcr-6.1) Variant Genes Identified in Moraxella Species Isolated from Pigs in Great Britain from 2014 to 2015. J. Antimicrob. Chemother. 2018, 73, 2904. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-Q.; Li, Y.-X.; Lei, C.-W.; Zhang, A.-Y.; Wang, H.-N. Novel Plasmid-Mediated Colistin Resistance Gene Mcr-7.1 in Klebsiella Pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a Novel Mobile Colistin Resistance Gene, Mcr-8, in NDM-Producing Klebsiella Pneumoniae. Emerg. Microbes Infect. 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene Mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella Enterica Serotype Typhimurium Isolate. mBio 2019, 10, e00853-19. [Google Scholar] [CrossRef]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of Novel Mobile Colistin Resistance Gene Mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Johnson, A.P. Transferable Resistance to Colistin: A New but Old Threat: Table 1. J. Antimicrob. Chemother. 2016, 71, 2066–2070. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P. Emerging Plasmid-Encoded Colistin Resistance: The Animal World as the Culprit? J. Antimicrob. Chemother. 2016, 71, 2326–2327. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P. (Ed.) Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; M100; Clinical and Laboratory Standards Institute (USA): Berwyn, PA, USA, 2021; ISBN 978-1-68440-104-8. [Google Scholar]
- Bianco, A.; Capozzi, L.; Monno, M.R.; Del Sambro, L.; Manzulli, V.; Pesole, G.; Loconsole, D.; Parisi, A. Characterization of Bacillus Cereus Group Isolates From Human Bacteremia by Whole-Genome Sequencing. Front. Microbiol. 2021, 11, 599524. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Teixeira, M.; Pillay, S.; Urhan, A.; Abeel, T. SHIP: Identifying Antimicrobial Resistance Gene Transfer between Plasmids. Bioinformatics 2023, 39, btad612. [Google Scholar] [CrossRef] [PubMed]
- Sheu, C.-C.; Chang, Y.-T.; Lin, S.-Y.; Chen, Y.-H.; Hsueh, P.-R. Infections Caused by Carbapenem-Resistant Enterobacteriaceae: An Update on Therapeutic Options. Front. Microbiol. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Kujat Choy, S.; Neumann, E.-M.; Romero-Barrios, P.; Tamber, S. Contribution of Food to the Human Health Burden of Antimicrobial Resistance. Foodborne Pathog. Dis. 2024, 21, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Sparo, M.; Delpech, G.; García Allende, N. Impact on Public Health of the Spread of High-Level Resistance to Gentamicin and Vancomycin in Enterococci. Front. Microbiol. 2018, 9, 3073. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; McENTIRE, J.C.; Zhang, L.; Li, X.; Doyle, M.P. The Transfer of Antibiotic Resistance from Food to Humans: Facts, Implications and Future Directions: -EN- -FR- Le Transfert de l’antibiorésistance à l’homme Par l’intermédiaire Des Aliments: Les Faits, Les Conséquences et Les Orientations Futures -ES- Transferencia de La Resistencia a Los Antibióticos de Los Alimentos al Hombre: Datos, Consecuencias y Orientaciones Futuras. Rev. Sci. Tech. OIE 2012, 31, 249–260. [Google Scholar] [CrossRef]
- Lencina, F.A.; Bertona, M.; Stegmayer, M.A.; Olivero, C.R.; Frizzo, L.S.; Zimmermann, J.A.; Signorini, M.L.; Soto, L.P.; Zbrun, M.V. Prevalence of Colistin-Resistant Escherichia Coli in Foods and Food-Producing Animals through the Food Chain: A Worldwide Systematic Review and Meta-Analysis. Heliyon 2024, 10, e26579. [Google Scholar] [CrossRef]
- Vincenti, S.; Raponi, M.; Sezzatini, R.; Giubbini, G.; Laurenti, P. Enterobacteriaceae Antibiotic Resistance in Ready-to-Eat Foods Collected from Hospital and Community Canteens: Analysis of Prevalence. J. Food Prot. 2018, 81, 424–429. [Google Scholar] [CrossRef]
- Mushtaq, S.; Reynolds, R.; Gilmore, M.C.; Esho, O.; Adkin, R.; García-Romero, I.; Chaudhry, A.; Horner, C.; Bartholomew, T.L.; Valvano, M.A.; et al. Inherent Colistin Resistance in Genogroups of the Enterobacter Cloacae Complex: Epidemiological, Genetic and Biochemical Analysis from the BSAC Resistance Surveillance Programme. J. Antimicrob. Chemother. 2020, 75, 2452–2461. [Google Scholar] [CrossRef]
- Germanou, D.; Spernovasilis, N.; Papadopoulos, A.; Christodoulou, S.; Agouridis, A.P. Infections Caused by Moellerella Wisconsensis: A Case Report and a Systematic Review of the Literature. Microorganisms 2022, 10, 892. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martínez, L.; Silva, J.; Martínez-Romero, E. Klebsiella Variicola, A Novel Species with Clinical and Plant-Associated Isolates. Syst. Appl. Microbiol. 2004, 27, 27–35. [Google Scholar] [CrossRef]
- Leroy, A.-G.; Malandain, D.; Duchalais, É.; Meurette, G.; Corvec, S. Accurate MALDI-TOF Mass Spectrometry Identification of a Colistin-Resistant Moellerella Wisconsensis Strain. Médecine Et Mal. Infect. 2016, 46, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Parisi, S.G.; Bartolini, A.; Santacatterina, E.; Castellani, E.; Ghirardo, R.; Berto, A.; Franchin, E.; Menegotto, N.; De Canale, E.; Tommasini, T.; et al. Prevalence of Klebsiella Pneumoniae Strains Producing Carbapenemases and Increase of Resistance to Colistin in an Italian Teaching Hospital from January 2012 To December 2014. BMC Infect Dis 2015, 15, 244. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Feng, Y.; McNally, A.; Zong, Z. Occurrence of Colistin-Resistant Hypervirulent Klebsiella Variicola. J. Antimicrob. Chemother. 2018, 73, 3001–3004. [Google Scholar] [CrossRef] [PubMed]
- Hristova, P.M.; Alexandrova, A.S.; Lukanov, M.; Hitkova, H.Y.; Borisov, B.K. Catheter-Related Bloodstream Infection in Hemodialysis Patient Due to Atlantibacter Hermannii. Case Rep. Nephrol. Dial. 2023, 13, 142–147. [Google Scholar] [CrossRef]
- Cakir, N.; Süer, K.; Duygu, H. A Very Rare Diabetic Foot Co-Infection Due to Escherichia Hermannii and Enterobacter Aerogenes in an Elderly Patient. J. Clin. Diagn. Res. 2019, 13, DD01–DD02. [Google Scholar] [CrossRef]
- Shin, J.H.; Shin, D.; Kwon, K.T.; Ko, K.S. Colistin Heteroresistance in Citrobacter Freundii Clinical Isolates from Republic of Korea. Diagn. Microbiol. Infect. Dis. 2024, 108, 116187. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia Coli. Microbiol Spectr 2018, 6, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Schrauwen, E.J.A.; Huizinga, P.; Van Spreuwel, N.; Verhulst, C.; Kluytmans-van Den Bergh, M.F.Q.; Kluytmans, J.A.J.W. High Prevalence of the Mcr-1 Gene in Retail Chicken Meat in the Netherlands in 2015. Antimicrob. Resist. Infect. Control 2017, 6, 83. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Alali, W.Q.; Mahmoud, S.A.; Ghazawi, A.; Bharathan, G.; Melegh, S.; Rizvi, T.A.; Pál, T. Molecular Characterization of MCR-1 Producing Enterobacterales Isolated in Poultry Farms in the United Arab Emirates. Antibiotics 2022, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Nakayama, T.; Ariyoshi, T.; Uehara, S.; Uchitani, Y.; Tsuchida, S.; Nishiyama, H.; Morioka, I.; Koshinaga, T.; Kusabuka, A.; et al. Emergence of Phytobacter Diazotrophicus Carrying an IncA/C 2 Plasmid Harboring Bla NDM-1 in Tokyo, Japan. mSphere 2023, 8, e00147-23. [Google Scholar] [CrossRef]
- Smits, T.H.M.; Arend, L.N.V.S.; Cardew, S.; Tång-Hallbäck, E.; Mira, M.T.; Moore, E.R.B.; Sampaio, J.L.M.; Rezzonico, F.; Pillonetto, M. Resolving Taxonomic Confusion: Establishing the Genus Phytobacter on the List of Clinically Relevant Enterobacteriaceae. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Al-Kharousi, Z.S.; Guizani, N.; Al-Sadi, A.M.; Al-Bulushi, I.M. Antibiotic Resistance of Enterobacteriaceae Isolated from Fresh Fruits and Vegetables and Characterization of Their AmpC β-Lactamases. J. Food Prot. 2019, 82, 1857–1863. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhou, H.; Qin, L.; Pang, Z.; Qin, T.; Ren, H.; Pan, Z.; Zhou, J. Frequency, Antimicrobial Resistance and Genetic Diversity of Klebsiella Pneumoniae in Food Samples. PLoS ONE 2016, 11, e0153561. [Google Scholar] [CrossRef] [PubMed]
- Falomir, M.P.; Rico, H.; Gozalbo, D. Enterobacter and Klebsiella Species Isolated from Fresh Vegetables Marketed in Valencia (Spain) and Their Clinically Relevant Resistances to Chemotherapeutic Agents. Foodborne Pathog. Dis. 2013, 10, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Cui, Y.; Quan, J.; Zhao, D.; Han, X.; Shi, Q.; Wang, Q.; Jiang, Y.; Du, X.; Li, X.; et al. High Prevalence of Colistin Resistance and Mcr-9/10 Genes in Enterobacter Spp. in a Tertiary Hospital over a Decade. Int. J. Antimicrob. Agents 2022, 59, 106573. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Li, X.-P.; Fang, L.-X.; Jiang, P.; Pan, D.; Xia, J.; Liao, X.-P.; Liu, Y.-H.; Sun, J. Emergence of the Colistin Resistance Gene Mcr-1 in Citrobacter Freundii. Int. J. Antimicrob. Agents 2017, 49, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Boolchandani, M.; D’Souza, A.W.; Dantas, G. Sequencing-Based Methods and Resources to Study Antimicrobial Resistance. Nat. Rev. Genet. 2019, 20, 356–370. [Google Scholar] [CrossRef]
- Tang, B.; Chang, J.; Chen, Y.; Lin, J.; Xiao, X.; Xia, X.; Lin, J.; Yang, H.; Zhao, G. Escherichia Fergusonii, an Underrated Repository for Antimicrobial Resistance in Food Animals. Microbiol. Spectr. 2022, 10, e01617-21. [Google Scholar] [CrossRef] [PubMed]
- Scheller, D.; Twittenhoff, C.; Becker, F.; Holler, M.; Narberhaus, F. OmpA, a Common Virulence Factor, Is Under RNA Thermometer Control in Yersinia Pseudotuberculosis. Front. Microbiol. 2021, 12, 687260. [Google Scholar] [CrossRef] [PubMed]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids Carrying Antimicrobial Resistance Genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | N. of Samples | Number of Positive Food Samples | Percentage of Positive Samples for Type of Food (%) | N. of Isolates | Organism (n., Name of Food) |
|---|---|---|---|---|---|
| Raw Milk | 52 | 2 | 4 | 2 | E. hormaechei (1, bulk cow milk) |
| E. mori (1, bulk cow milk) | |||||
| Raw Meat | 207 | 11 | 5 | 11 | E. ludwigii (1, bovine meat) E. bugandensis (1, minced beef/pork) |
| E. kobei (1, fresh pork sausage, 1, sausage dough) | |||||
| E. asburiae(1, pork meat) | |||||
| M. wisconsensis (1, beef meat; 1, minced beef; 1, pork fresh sausages; 1, beef and pork hamburgher; 1, poultry meat) | |||||
| E. coli (1, turkey meat) | |||||
| Raw Seafood products | 133 | 4 | 3 | 4 | A. hermannii (1, filleted sea bass) |
| E. hormaechei (1, sea bass) | |||||
| M. wisconsensis (1, filleted sea bass) | |||||
| R. ornithinolytica (1, clams) | |||||
| Raw Bakery and pastry products, fresh pasta | 28 | 2 | 7 | 3 | E. cancerogenus (1, raw shortcrust pastry; 1, fresh egg pasta *) |
| E. kobei (1, fresh egg pasta *) | |||||
| Vegetables | 80 | 6 | 8 | 6 | E. bugandensis (1, beets) |
| E. cloacae (2, cucumbers) | |||||
| E. ludwigii (1, celery) | |||||
| E. kobei (1, frozen celery) | |||||
| K. cowanii (1, wheat flour) | |||||
| TOT | 500 | 25 | 5 | 26 |
| Source | N. of Samples | Number of Positive Food Samples | Percentage of Positive Samples for Type of Food (%) | N. of Isolates | Organism (n., Name of Food) |
|---|---|---|---|---|---|
| Milk and Cheese | 240 | 8 | 3 | 8 | E. asburiae (1, burrata cheese) |
| E. kobei (1, mozzarella; 1, sheep and goat cheese; 1, giuncata cheese) | |||||
| E. hormaechei (1, fresh cheese; 1, giuncata cheese) | |||||
| A. hermannii (1, mozzarella; 1, fresh cheese) | |||||
| Dried or cooked Sausages | 35 | 1 | 3 | 1 | A. hermannii (1, cured sausage) |
| Ready meals | 100 | 3 | 3 | 3 | E. ludwigii (1, seasoned salad) |
| C. freundii (1, grilled swordfish with olives) | |||||
| M. wisconsensis (1, breaded salmon fillet) | |||||
| Bakery and pastry products, fresh pasta | 27 | 0 | 0 | 0 | 0 |
| Ice-cream | 66 | 5 | 8 | 5 | K. pneumoniae (1, homemade ice-cream) |
| E. asburiae (2, homemade ice-cream) | |||||
| E. hormaechei (1, homemade ice-cream) | |||||
| P. diazotrophicus (1, homemade ice-cream) | |||||
| Vegetables | 32 | 2 | 6 | 2 | K. variicola (1, IV range mixed salad) |
| E. ludwigii (1, IV range fruit salad) | |||||
| TOT | 500 | 19 | 4 | 19 |
| Raw Food Samples | Organism | ID Samples |
| Source | Species | |
| Raw turkey meat | E. coli | S075 |
| Raw sea bass fillet | A. hermannii | S128 |
| Raw clams | R. ornithinolytica | S090 |
| Raw sea bass fillet | M. wisconsensis | S131 |
| Raw bovine meat | M. wisconsensis | S037 |
| Raw poultry meat | M. wisconsensis | S073 |
| Fresh pork sausage | M. wisconsensis | S091 |
| Raw minced bee | M. wisconsensis | S092 |
| Raw hamburger | M. wisconsensis | S089 |
| Wheat flour | K. cowanii | S132 |
| Raw pork meat | E. asburiae | S027 |
| Raw minced beef and pork | E. bugandensis | S071 |
| Beets (no rte) | E. bugandensis | S050 |
| Cucumber (no rte) | E. cloacae | S051 |
| Cucumber (no rte) | E. cloacae | S079 |
| Raw shortcrust pastry | E. cancerogenus | S127 |
| Fresh egg pasta * | *E. cancerogenus | S140 |
| Bulk cow milk | E. hormaechei | S034 |
| Raw sea bass | E. hormaechei | S086 |
| Raw bovine meat | E. ludwigii | S026 |
| Celery (no rte) | E. ludwigii | S076 |
| Bulk cow milk | E. mori | S045 |
| Raw fresh pork sausage | E. kobei | S060 |
| Raw sausage dough | E. kobei | S085 |
| Celery (no rte) | E. kobei | S077 |
| Fresh egg pasta * | * E. kobei | S121 |
| Rte Samples | Organism | ID Samples |
| Source | Species | |
| Homemade ice cream | K. pneumoniae | S039 |
| Mixed salad (rte) | K. variicola | S094 |
| Mozzarella | A. hermannii | S118 |
| Fresh cheese | A. hermannii | S126 |
| Seasoned sausage (rte) | A. hermannii | S059 |
| Grilled swordfish (rte) | C. freundii | S114 |
| Breaded salmon fillet (rte) | M. wisconsensis | S115 |
| Homemade ice cream | P. diazotrophicus | S093 |
| Burrata | E. asburiae | S120 |
| Homemade ice cream | E. asburiae | S112 |
| Homemade ice cream | E. asburiae | S113 |
| Fesh cheese | E. hormaechei | S072 |
| Seasoned salad (rte) | E. ludwigii | S081 |
| Fruit salad (rte) | E. ludwigii | S080 |
| Mozzarella | E. kobei | S056 |
| Sheep and goat cheese | E. kobei | S074 |
| Giuncata | E. kobei | S122 |
| Giuncata | E. hormaechei | S123 |
| Homemade ice-cream | E. hormaechei | S084 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraccalvieri, R.; Bianco, A.; Difato, L.M.; Capozzi, L.; Del Sambro, L.; Castellana, S.; Donatiello, A.; Serrecchia, L.; Pace, L.; Farina, D.; et al. Isolation and Characterization of Colistin-Resistant Enterobacteriaceae from Foods in Two Italian Regions in the South of Italy. Microorganisms 2025, 13, 163. https://doi.org/10.3390/microorganisms13010163
Fraccalvieri R, Bianco A, Difato LM, Capozzi L, Del Sambro L, Castellana S, Donatiello A, Serrecchia L, Pace L, Farina D, et al. Isolation and Characterization of Colistin-Resistant Enterobacteriaceae from Foods in Two Italian Regions in the South of Italy. Microorganisms. 2025; 13(1):163. https://doi.org/10.3390/microorganisms13010163
Chicago/Turabian StyleFraccalvieri, Rosa, Angelica Bianco, Laura Maria Difato, Loredana Capozzi, Laura Del Sambro, Stefano Castellana, Adelia Donatiello, Luigina Serrecchia, Lorenzo Pace, Donatella Farina, and et al. 2025. "Isolation and Characterization of Colistin-Resistant Enterobacteriaceae from Foods in Two Italian Regions in the South of Italy" Microorganisms 13, no. 1: 163. https://doi.org/10.3390/microorganisms13010163
APA StyleFraccalvieri, R., Bianco, A., Difato, L. M., Capozzi, L., Del Sambro, L., Castellana, S., Donatiello, A., Serrecchia, L., Pace, L., Farina, D., Galante, D., Caruso, M., Tempesta, M., & Parisi, A. (2025). Isolation and Characterization of Colistin-Resistant Enterobacteriaceae from Foods in Two Italian Regions in the South of Italy. Microorganisms, 13(1), 163. https://doi.org/10.3390/microorganisms13010163