
Microbial Diversity in Extreme Marine Habitats and Their Biomolecules



Abstract
:1. Introduction
2. Habitat: Hydrothermal Vents
2.1. Microbial Diversity
2.1.1. Hyper-Thermophilic and Thermophilic Microorganisms
2.1.2. Thermopiezophilic Microorganisms
2.1.3. Some Examples of Hyperthermophiles, Thermophiles and Mesophiles from Different Marine Areas
2.2. Metagenomic Analyses
2.3. Applications
2.3.1. Thermophilic Microorganisms and Their Metabolic Peculiarity
2.3.2. Thermozymes and Biopolymers
2.3.3. Other Compounds from Marine Thermophiles
3. Habitat: Cold Marine Environments
3.1. Microbial Diversity
3.1.1. Psychrophilic Microorganisms
3.1.2. Adaptation Strategies of Psycrophilic Microorganism and Their Metabolic Studies
3.2. Metagenomic Analysis
3.3. Applications
3.3.1. Enzyme Kinetics
3.3.2. Exopolysaccharide Producing Psycrophilic Microorganisms
4. Habitat: Hypersaline Environments
4.1. Microbial Diversity
4.1.1. Halophilic Microorganisms
4.1.2. Adaptive Strategies
4.2. Metagenomic Analysis
4.3. Applications of Halophiles
4.3.1. Enzymes
4.3.2. Biopolymers
4.3.3. Osmolytes
5. Poly-Extremophiles
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-mining the microbial treasures of the ocean: New natural products. Biotech. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhang, L.; Bazylinski, D.A. Deep-sea piezosphere and piezophiles: Geomicrobiology and biogeochemistry. Trends Microbiol. 2010, 18, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Kato, C.; Nogi, Y.; Arakawa, S. Isolation, Cultivation, and Diversity of Deep-Sea Piezophiles. In High-Pressure Microbiology; Michiels, C., Bartlett, D., Aersten, A., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 203–217. ISBN 978-1-55581-423-6. [Google Scholar]
- Oren, A. Taxonomy of halophilic Archaea: Current status and future challenges. Extremophiles 2014, 18, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.; Muyzer, G. Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Fernandez, A.B.; León, M.J.; Sanchez-Porro, C.; Rodriguez-Valera, F. The Santa Pola saltern as a model for studying the microbiota of hypersaline environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.S.S.; Salamah, A.A.; Badawi, Y.B.E.; Tayeb Mohamed, A.E.; Antranikian, G. Detergent, solvent and salt compatible thermoactive alkaline serine protease from halotolerant alkaliphilic Bacillus sp. NPST.AK15: Purification and characterization. Extremophiles 2015, 19, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.; Jorge, C.D.; Gonçalves, L.G.; Gonçalves, S.; Matias, P.M.; Santos, H. Mannosyl-glycerate: Structural analysis of biosynthesis and evolutionary history. Extremophiles 2014, 18, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Forterre, P.; Prangishvili, D.; Krupovic, M.; Koonin, E.V. Dark matter in archaeal genomes: A rich source of novel mobile elements, defence systems and secretory complexes. Extremophiles 2014, 18, 877–893. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Lentini, V.; Bunk, B.; Overmann, J.; Italiano, F.; Maugeri, T.L. Changes in prokaryotic community composition accompanying a pronounced temperature shift of a shallow marine thermal brine pool (Panarea Island, Italy). Extremophiles 2015, 19, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Klippel, B.; Sahm, K.; Basner, A.; Wiebusch, S.; John, P.; Lorenz, U.; Peters, A.; Abe, F.; Takahashi, K.; Kaiser, O.; Goesmann, A.; Jaenicke, S.; Grote, R.; Horikoshi, K.; Antranikian, G. Carbohydrate active enzymes identified by metagenomic analysis of deep-sea sediment bacteria. Extremophiles 2014, 18, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Gimmler, A.; Korn, R.; de Vargas, C.; Audic, S.; Stoeck, T. The Tara Oceans voyage reveals global diversity and distribution patterns of marine planktonic ciliates. Sci. Rep. 2016, 6, 33555. [Google Scholar] [CrossRef] [PubMed]
- Yooseph, S.; Sutton, G.; Rusch, D.B.; Halpern, A.L.; Williamson, S.J.; Remington, K.; Eisen, J.A.; Heidelberg, K.B.; Manning, G.; Li, W.; et al. The Sorcerer II Global Ocean Sampling expedition: Expanding the universe of protein families. PLoS Biol. 2007, 5, e16. [Google Scholar] [CrossRef] [PubMed]
- Kopf, A.; Bicak, M.; Kottmann, R.; Schnetzer, J.; Kostadinov, I.; Lehmann, K.; Fernandez-Guerra, A.; Jeanthon, C.; Rahav, E.; Ullrich, M.; et al. The ocean sampling day consortium. GigaScience 2015, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Orikasa, Y.; Hayashi, H.; Watanabe, K.; Yoshida, K.; Okuyama, H. Occurrence of trans monounsaturated and polyunsaturated fatty acids in Colwellia psychrerythraea strain 34H. J. Basic Microbiol. 2015, 55, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Industrial and environmental applications of halophilic microorganisms. Environ. Techn. 2010, 31, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, B.P.; Dodsworth, J.A.; Murugapiran, S.K.; Rinke, C.; Woyke, T. Impact of single-cell genomics and metagenomics on the emerging view of extremophile ‘‘microbial dark matter”. Extremophiles 2014, 18, 865–887. [Google Scholar] [CrossRef] [PubMed]
- Finore, I.; Di Donato, P.; Mastascusa, V.; Nicolaus, B.; Poli, A. Fermentation Technologies for the Optimization of Marine Microbial Exopolysaccharide Production. Mar. Drugs 2014, 12, 3005–3024. [Google Scholar] [CrossRef] [PubMed]
- León, M.J.; Fernández, A.B.; Ghai, R.; Sánchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. From metagenomics to pure culture: Isolation and characterization of the moderately halophilic bacterium Spiribacter. salinus gen. nov., sp. nov. Appl. Environ. Microbiol. 2014, 80, 3850–3857. [Google Scholar] [CrossRef] [PubMed]
- Arena, A.; Maugeri, T.L.; Pavone, B.; Iannello, D.; Gugliandolo, C.; Bisignano, G. Antiviral and immunoregulatory effect of a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis. Int. Immunopharmacol. 2006, 6, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Hong, Y.; Shao, Z.; Zhao, Y.; Liu, Z. A novel cold-active and alkali-stable beta-glucosidase gene isolated from the marine bacterium Martelella mediterranea. Appl. Biochem. Biotechnol. 2010, 162, 2136–2148. [Google Scholar] [CrossRef] [PubMed]
- Elleuche, S.; Schafers, C.; Blank, S.; Schroder, C.; Antranikian, G. Exploration of extremophiles for high temperature biotechnological processes. Curr. Opin. Microbiol. 2015, 25, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Canganella, F. Hydrothermal vent ecosystems and representative hyperthermophilic microorganisms. Ann. Microbiol. 2001, 51, 11–27. [Google Scholar]
- Cambon-Bonavita, M.A.; Raguénès, G.; Jean, J.; Vincent, P.; Guezennec, J. A novel polymer produced by a bacterium isolated from a deep-sea hydrothermal vent polychaete annelid. J. Appl. Microbiol. 2002, 93, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microb. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea., Bacteria, and Eukarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed]
- Kashefi, K.; Lovley, D.R. Extending the upper temperature limit for life. Science 2003, 301, 934. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [PubMed]
- Stetter, K.O.; König, H.; Stackebrandt, E. Pyrodictium. gen. nov., a new genus of submarine disc-shaped sulphur reducing archaebacteria growing optimally at 105 °C. Syst. Appl. Microbiol. 1983, 4, 535–551. [Google Scholar] [CrossRef]
- Pley, U.; Schipka, J.; Gambacorta, A.; Jannasch, H.W.; Fricke, H.; Rachel, R.; Stetter, K.O. Pyrodictium abyssi sp. nov. represents a novel heterotrophic marine archaeal hyperthermophile growing at 110 °C. Syst. Appl. Microbiol. 1991, 14, 245–253. [Google Scholar] [CrossRef]
- Zillig, W.; Holz, I.; Wunderl, S. Hyperthermus. butylicus gen. nov., sp. nov., a hyperthermophilic, anaerobic, peptide-fermenting, facultatively H2S-generating archaebacterium. Int. J. Syst. Bacteriol. 1991, 41, 169–170. [Google Scholar] [CrossRef]
- Hensley, S.A.; Jung, J.H.; Park, C.S.; Holden, J.F. Thermococcus. paralvinellae sp. nov. and Thermococcus. cleftensis sp. nov. of hyperthermophilic heterotrophs from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2014, 64, 3655–3659. [Google Scholar] [CrossRef] [PubMed]
- Alain, K.; Marteinsson, V.T.; Miroshnichenko, M.L.; Bonch-Osmolovskaya, E.A.; Prieur, D.; Birrien, J.L. Marinitoga. piezophila sp. nov., a rod-shaped, thermo-piezophilic bacterium isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2002, 52, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Miyazaki, M.; Hirayama, H.; Nakagawa, S.; Querellou, J.; Godfroy, A. Isolation and physiological characterization of two novel, piezophilic, thermophilic chemolithoautotrophs from a deep-sea hydrothermal vent chimney. Environ. Microbiol. 2009, 11, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, M.O.; Kelley, D.S.; Bolton, S.A.; Baross, J.A. Low archaeal diversity linked to subsea floor geochemical processes at the Lost City Hydrothermal Field, Mid-Atlantic Ridge. Environ. Microbiol. 2004, 6, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, T.L.; Bianconi, G.; Canganella, F.; Danovaro, R.; Gugliandolo, C.; Italiano, F.; Lentini, V.; Manini, E.; Nicolaus, B. Shallow hydrothermal vents in the southern Tyrrhenian Sea. Chem. Ecol. 2010, 26, 285–298. [Google Scholar] [CrossRef]
- Neuner, A.; Jannasch, H.W.; Belkin, S.; Stetter, K.O. Thermococcus. litoralis sp. nov.: A new species of extremely thermophilic marine archaeabacteria. Arch. Microbiol. 1990, 153, 205–207. [Google Scholar] [CrossRef]
- Nicolaus, B.; Lama, L.; Panico, A.; Schiano Moriello, V.; Romano, I.; Gambacorta, A. Production and characterization of exopolysaccharides excreted by thermophilic bacteria from shallow, marine hydrothermal vents of Flegrean Areas (Italy). Syst. Appl. Microbiol. 2002, 25, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Völkl, P.; Markiewicz, P.; Baikalov, C.; Fitz-Gibbon, S.; Stetter, K.O.; Miller, J.H. Genomic and cDNA sequence tags of the hyperthermophilic archaeon Pyrobaculum. aerophilum. Nucleic Acids Res. 1996, 24, 4373–4378. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D.; Freeze, H. Thermus aquaticus gen. n. and sp. n., a Nonsporulating Extreme Thermophile. J. Bacteriol. 1969, 98, 289–297. [Google Scholar] [PubMed]
- Barbier, G.; Godfroy, A.; Meunier, J.R.; Quérellou, J.; Cambon, M.A.; Lesongeur, F.; Grimont, P.A.; Raguénèset, G. Pyrococcus glycovorans sp. nov., a hyperthermophilic archaeon isolated from the East Pacific Rise. Int. J. Syst. Bacteriol. 1999, 49, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Birrien, J.L.; Zeng, X.; Jebbar, M.; Cambon-Bonavita, M.A.; Quérellou, J.; Oger, P.; Bienvenu, N.; Xiao, X.; Prieur, D. Pyrococcus. yayanosii sp. nov., an obligate piezophilic hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2011, 61, 2827–2831. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.; Völker, H.; Thomm, M. Thermococcus. aegaeicus sp. nov. and Staphylothermus. hellenicus sp. nov., two novel hyperthermophilic archaea isolated from geothermally heated vents off Palaeochori Bay, Milos, Greece. Int. J. Syst. Evol. Microbiol. 2000, 50, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.H.; Kelly, R.M. Characterization of amylolytic enzymes, having both alpha-1,4 and alpha-1,6 hydrolytic activity, from the thermophilic archaea Pyrococcus. furiosus and Thermococcus. litoralis. Appl. Environ. Microbiol. 1993, 59, 2614–2621. [Google Scholar] [PubMed]
- Miroshnichenko, M.L.; Bonch-Osmolovskaya, E.A.; Neuner, A.; Kostrikina, N.A.; Chernych, N.A.; Alekseev, V.A. Thermococcus. stetteri sp. nov., a new extremely thermophilic marine sulfur-metabolizing Archaebacterium. Syst. Appl. Microbiol. 1989, 12, 57–262. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kwak, Y.S.; Akiba, T.; Kudo, T.; Horikoshi, K. Thermococcus. profundus sp. nov., a new hyperthermophilic archaeon isolated from deep-sea hydrothermal vent. Syst. Appl. Microbiol. 1994, 17, 232–236. [Google Scholar] [CrossRef]
- Cambon-Bonavita, M.A.; Lesongeur, F.; Pignet, P.; Wery, N.; Lambert, C.; Godfroy, A.; Querellou, J.; Georges Barbier, G. Thermococcus. atlanticus sp. nov., a hyperthermophilic Archaeon isolated from a deep-sea hydrothermal vent in the Mid-Atlantic Ridge. Extremophiles 2003, 7, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Gorlas, A.; Croce, O.; Oberto, J.; Gauliard, E.; Forterre, P.; Marguet, E. Thermococcus. nautili sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal deep-sea vent. Int. J. Syst. Evol. Microbiol. 2014, 64, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- Gorlas, A.; Croce, O.; Oberto, J.; Gauliard, E.; Forterre, P.; Marguet, E. Thermococcus. prieurii sp. nov., a hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2013, 63, 2920–2926. [Google Scholar] [CrossRef] [PubMed]
- Pikuta, E.V.; Marsic, D.; Itoh, T.; Bej, A.K.; Tang, J.; Whitman, W.B.; Ng, J.D.; Garriot, O.K.; Hoover, R.B. Thermococcus. thioreducens sp. nov., a novel hyperthermophilic, obligately sulfur-reducing archaeon from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2007, 57, 1612–1618. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, M.O.; Kelley, D.S.; Delaney, J.R.; Baross, J.A. Incidence and diversity of microorganisms within the walls of an active deep-sea sulfide chimney. Appl. Environ. Microbiol. 2003, 69, 3580–3592. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.B.; Antranikian, G. Keratin degradation by Fervidobacterium. pennivorans, a novel thermophilic anaerobic species of the order thermotogales. Appl. Environ. Microbiol. 1996, 62, 2875–2882. [Google Scholar] [PubMed]
- Duffaud, G.D.; McCutchen, C.M.; Leduc, P.; Parker, K.N.; Kelly, R.M. Purification and characterization of extremely thermostable beta-mannanase, beta-mannosidase, and alpha-galactosidase from the hyperthermophilic eubacterium Thermotoga. neapolitana 5068. Appl. Environ. Microbiol. 1997, 63, 169–177. [Google Scholar] [PubMed]
- Atomi, H. Recent progress towards the application of hyperthermophiles and their enzymes. Curr. Opin. Chem. Biol. 2005, 9, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Klingeberg, M.; Hashwa, F.; Antranikian, G. Properties of extremely thermostable proteases from anaerobic hyperthermophilic bacteria. Appl. Microbiol. Biotechnol. 1991, 34, 715–719. [Google Scholar] [CrossRef]
- Jang, H.J.; Kim, B.C.; Pyun, Y.R.; Kim, Y.S. A novel subtilisin-like serine protease from Thermoanaerobacter. yonseiensis KB-1: Its cloning, expression, and biochemical properties. Extremophiles 2002, 6, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, K.G.; Schreiber, L.; Petersen, D.G.; Kjeldsen, K.U.; Lever, M.A.; Steen, A.D.; Stepanauskas, R.; Richter, M.; Kleindienst, S.; Lenk, S.; et al. Predominant archaea in marine sediments degrade detrital proteins. Nature 2013, 496, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Pietrow, O.; Panek, A.; Synowiecki, J. Extracellular proteolytic activity of Deinococcus. geothermalis. Afr. J. Biotechnol. 2013, 12, 4020–4027. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Salehghamari, E.; Khajeh, K.; Kabiri, M.; Naddaf, S. Production of an extracellular thermohalophilic lipase from a moderately halophilic bacterium, Salinivibrio. sp. strain SA-2. J. Basic Microbiol. 2008, 48, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Spanò, A.; Gugliandolo, C.; Lentinia, V.; Maugeri, T.L.; Anzelmo, G.; Poli, A.; Nicolaus, B. A novel EPS-producing strain of Bacillus licheniformis isolated from a shallow vent off Panarea Island (Italy). Curr. Microbiol. 2013, 67, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Rougeaux, H.; Kervarec, N.; Pichon, R.; Guerzennec, J. Structure of the exopolysaccharide of Vibrio diabolicus isolated from a deep–sea hydrothermal vent. Carbohydr. Res. 1999, 322, 40–45. [Google Scholar] [CrossRef]
- Harmsen, H.J.M.; Prieur, D.; Jeanthon, C. Distribution of microorganisms in deep-sea hydrothermalvent chimneys investigated by whole-cell hybridization and enrichment culture of thermophilic subpopulations. Appl. Environ. Microbiol. 1997, 63, 2876–2883. [Google Scholar] [PubMed]
- Harmsen, H.J.M.; Prieur, D.; Jeanthon, C. Group-specific 16S-rRNA-targeted oligonucleotide probes to identify thermophilic bacteria in marine hydrothermal vents. Appl. Environ. Microbiol. 1997b, 63, 4061–4068. [Google Scholar]
- Huber, H.; Stetter, K.O. Archaeoglobales ord. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 349–353. ISBN 978-0-387-21609-6. [Google Scholar]
- Huber, H.; Stetter, K.O. Desulfurococcales. ord. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 180–196. ISBN 978-0-387-21609-6. [Google Scholar]
- Huber, H.; Stetter, K.O. Thermoproteales. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 170–179. ISBN 978-0-387-21609-6. [Google Scholar]
- Takai, K.; Inagaki, F.; Nakagawa, S.; Hirayama, H.; Nunoura, T.; Sako, Y.; Nealson, K.A.; Horikoshi, K. Isolation and phylogenetic diversity of members of previously uncultivated ε-Proteobacteria in deep-sea hydrothermal vents. FEMS Microbiol. Lett. 2003, 218, 167–174. [Google Scholar] [PubMed]
- Kormas, K.A.; Tivey, M.; Von Damm, K.; Teske, A. Bacterial and archaeal phylotypes associated with distinct mineralogical layers of a white smoker spire from a deep-sea hydrothermal vent site (9° N, East Pacific Rise). Environ. Microbiol. 2006, 8, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Wang, F.; Guo, L.; Chen, Z.; Sievert, S.M.; Meng, J.; Huang, G.; Li, Y.; Yan, Q.; Wu, S.; et al. Comparative metagenomics of microbial communities inhabiting deep-sea hydrothermal vent chimneys with contrasting chemistries. Int. J. Syst. Evol. Microbiol. 2011, 5, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Guazzaroni, M.E.; Morgante, V.; Mirete, S.; Gonzalez-Pastor, J.E. Novel acid resistance genes from the metagenome of the Tinto River, an extremely acidic environment. Environ. Microbiol. 2013, 15, 1088–1102. [Google Scholar] [CrossRef] [PubMed]
- Urich, T.; Lanez, N.A.; Stokke, R.; Pedersen, R.B.; Bayer, C.; Thorseth, I.H.; Schleper, C.; Steen, I.H.; Øvreas, L. Microbial community structure and functioning in marine sediments associated with diffuse hydrothermal venting assessed by integrated metaomics. Environ. Microbiol. 2014, 16, 2699–2710. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, B.; Trincone, A.; Esposito, E.; Vaccaro, M.R.; Gambacorta, A.; De Rosa, M. Calditol tetraether lipids of the archaebacterium Sulfolobus solfataricus. Biosynth. Stud. Biochem. J. 1990, 266, 785–791. [Google Scholar]
- Koga, Y. Thermal Adaptation of the Archaeal and Bacterial Lipid Membranes. Archaea 2012, 2012, 789652. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.J.; Lewis, H.; Trejo, E.; Winston, V.; Evilia, C. Protein Adaptations in Archaeal Extremophiles. Archaea 2013, 373275. [Google Scholar] [CrossRef] [PubMed]
- Dipasquale, L.; Romano, I.; Picariello, G.; Calandrelli, V.; Lama, L. Characterization of a native cellulose activity from an anaerobic thermophilic hydrogen-producing bacterium Thermosipho. sp. strain 3. Ann. Microbiol. 2014, 64, 1493–1503. [Google Scholar] [CrossRef]
- Poli, A.; Di Donato, P.; Abbamondi, G.R.; Nicolaus, B. Synthesis, Production, and Biotechnological Applications of Exopolysaccharides and Polyhydroxyalkanoates by Archaea. Archaea 2011, 2011, 693253. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Spanò, A.; Maugeri, T.L.; Poli, A.; Arena, A.; Nicolaus, B. Role of Bacterial Exopolysaccharides as Agents in Counteracting Immune Disorders Induced by Herpes Virus. Microorganisms 2015, 3, 464–483. [Google Scholar] [CrossRef] [PubMed]
- Lama, L.; Tramice, A.; Finore, I.; Anzelmo, G.; Calandrelli, V.; Pagnotta, E.; Tommonaro, G.; Poli, A.; Di Donato, P.; Nicolaus, B.; et al. Degradative actions of microbial xylanolytic activities on hemicelluloses from rhizome of Arundo donax. AMB Express 2014, 4, 55. [Google Scholar] [CrossRef] [PubMed]
- Finore, I.; Poli, A.; Di Donato, P.; Lama, L.; Trincone, A.; Fagnano, M.; Mori, M.; Nicolaus, B.; Tramice, A. The hemicellulose extract from Cynara. cardunculus: A source of value-added biomolecules produced by xylanolytic thermozymes. Green Chem. 2016, 18, 2460–2472. [Google Scholar] [CrossRef]
- Courtois, A.; Berthou, C.; Guezennec, J.; Boisset, C.; Bordron, A. Exopolysaccharides Isolated from Hydrothermal Vent Bacteria Can Modulate the Complement System. PLoS ONE 2014, 9, e94965. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.G.; Borges, N.; Serra, F.; Fernandes, P.L.; Dopazo, H.; Santos, H. Evolution of the biosynthesis of di-myo-inositol phosphate, a marker of adaptation to hot marine environments. Environ. Microbiol. 2012, 14, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Lamosa, P.; Gonçalves, L.G.; Rodrigues, M.V.; Martins, L.O.; Raven, N.D.; Santos, H. Occurrence of 1-glyceryl-1-myo-inosityl phosphate in hyperthermophiles. Appl. Environ. Microbiol. 2006, 72, 6169–6173. [Google Scholar] [CrossRef] [PubMed]
- Wilson, Z.E.; Brimble, M.A. Molecules derived from the extremes of life. Nat. Prod. Rep. 2009, 26, 1–140. [Google Scholar] [CrossRef]
- Deming, J.W.; Eicken, H. Life in ice. In Planets and Life: The Emerging Science of Astrobiology; Sullivan, W.T., Baross, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 292–312, ISBN-13: 9780521824217. [Google Scholar]
- Deming, J.W. Psychrophiles and polar regions. Curr. Opin. Microbiol. 2002, 5, 301–309. [Google Scholar] [CrossRef]
- Staley, J.T.; Junge, K.; Deming, J.W. And some like it cold: Sea ice Microbiology. In Biodiversity of Microbial Life: Foundation of Earth’s Biosphere; Staley, J.T., Reysenbach, A.L., Eds.; Wiley-Liss, Inc.: New York, NY, USA, 2001; pp. 423–438. [Google Scholar]
- Chen, M.; Xiao, X.; Wang, P.; Zeng, X.; Wang, F. Arthrobacter. ardleyensis sp. nov., isolated from Antarctic lake sediment and deep-sea sediment. Arch. Microbiol. 2005, 183, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Tang, S.K.; Zhang, Y.Q.; Li, Z.Y.; Yi, L.B.; Wang, Y.X.; Li, W.J.; Cui, X.L. Arthrobacter. halodurans sp. nov., a new halotolerant bacterium isolated from sea water. Antonie Leeuwenhoek 2009, 96, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Reddy, G.S.N.; Delille, D.; Shivaji, S. Arthrobacter. gangotriensis sp. nov. and Arthrobacter. kerguelensis sp. nov. from Antarctica. Int. J. Syst. Evol. Microbiol. 2004, 54, 2375–2378. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Bae, J.W.; Nam, Y.D.; Kwon, H.Y.; Park, J.R.; Shin, K.S.; Kim, K.H.; Quan, Z.X.; Rhee, S.K.; An, K.G.; et al. Arthrobacter. subterraneus sp. nov., isolated from deep subsurface water of the South Coast of Korea. J. Microbiol. Biotechnol. 2007, 17, 1875–1879. [Google Scholar] [PubMed]
- Bowman, J.P.; Nichols, D.S. Novel members of the family Flavobacteriaceae from Antarctic maritime habitats including Subsaximicrobium wynnwilliamsii gen. nov., sp. nov., Subsaximicrobium saxinquilinus sp. nov., Subsaxibacter broadyi gen. nov., sp. nov., Lacinutrix copepodicola gen. nov., sp. nov., and novel species of the genera Bizionia, Gelidibacter and Gillisia. Int. J. Syst .Evol. Microbiol. 2005, 55, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, I.; Kawasaki, K.; Iwata, H.; Matsuyama, H.; Okuyama, H. Assignment of Vibrio sp. strain ABE-1 to Colwelliamaris. sp. nov., a new psychrophilic bacterium. Int. J. Syst. Bacteriol. 1998, 48, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Hosoya, S.; Kato, C.; Horikoshi, K. Colwellia. piezophila sp. nov., a novel piezophilic species from deep-sea sediments of the Japan Trench. Int. J. Syst. Evol. Microbiol. 2004, 54, 1627–1631. [Google Scholar] [CrossRef] [PubMed]
- Marx, J.G.; Shelly, D.; Carpenter, J.W.; Deming, A. Production of cryoprotectant extracellular polysaccharide substances (EPS) by the marine psychrophilic bacterium Colwellia. psychrerythraea strain 34H under extreme conditions. Can. J. Microbiol. 2009, 55, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Carillo, S.; Casillo, A.; Pieretti, G.; Parrilli, E.; Sannino, F.; Bayer-Giraldi, M.; Cosconati, S.; Novellino, E.; Ewert, M.; Deming, J.W.; et al. A Unique Capsular Polysaccharide Structure from the Psychrophilic Marine Bacterium Colwellia. psychrerythraea 34H that mimics antifreeze (Glyco)proteins. J. Am. Chem. Soc. 2015, 137, 179–189. [Google Scholar] [CrossRef] [PubMed]
- De Pascale, D.; Cusano, A.M.; Autore, F.; Parrilli, E.; Di Prisco, G.; Marino, G.; Tutino, M.L. The cold-active Lip1 lipase from the Antarctic bacterium Pseudoalteromonas. haloplanktis TAC125 is a member of a new bacterial lipolytic enzyme family. Extremophiles 2008, 12, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L.A.; Lysenko, A.M.; Rohde, M.; Mikhailov, V.V.; Stackebrandt, E. Psychrobacter. maritimus sp. nov. and Psychrobacter. arenosus sp. nov., isolated from coastal sea ice and sediments of the Sea of Japan. Int. J. Syst. Evol. Microbiol. 2004, 54, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.; Lee, Y.M.; Hwang, C.Y.; Park, H.; Jung, Y.J.; Kim, M.K.; Hong, S.G.; Kim, J.H.; Lee, H.K. Psychroserpens. jangbogonensis sp. nov., a psychrophilic bacterium isolated from Antarctic marine sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Minami, H.; Sakaki, T.; Kasahara, H.; Watanabe, A.; Onoda, T.; Hirota, K.; Yumoto, I. Psychrobacter. oceani sp. nov., isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, I.; Hirota, K.; Sogabe, Y.; Nodasaka, Y.; Yokota, Y.; Hoshino, T. Psychrobacter. okhotskensis sp. nov., a lipase-producing facultative psychrophile isolated from the coast of the Okhotsk Sea. Int. J. Syst. Evol. Microbiol. 2003, 53, 1985–1989. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, A.; Honda, D.; Yamamoto, H.; Kitamura, K.; Higashihara, T. Phylogenetic analysis of psychrophilic bacteria isolated from the Japan Trench, including a description of the deep-sea species Psychrobacter. pacificensis sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Auman, A.J.; Breezee, J.L.; Gosink, J.J.; Kämpfer, P.; Staley, J.T. Psychromonas ingrahamii sp. nov., a novel gas vacuolate, psychrophilic bacterium isolated from Arctic polar sea ice. Int. J. Syst. Evol. Microbiol. 2006, 56, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; McCammon, S.A.; Nichols, D.S.; Skerratt, J.H.; Rea, S.M.; Nichols, P.D.; McMeekin, T.A. Shewanella gelidimarina sp. nov. and Shewanella frigidimarina sp. nov., novel Antarctic species with the ability to produce eicosapentaenoic acid (20:5 omega 3) and grow anaerobically by dissimilatory Fe(III) reduction. Int. J. Syst. Bacteriol. 1997, 47, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Srimathi, S.; Jayaraman, G.; Feller, G.; Danielsson, B.; Narayanan, P.R. Intrinsic halotolerance of the psychrophilic alpha-amylase from Pseudoalteromonas haloplanktis. Extremophiles 2007, 11, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Huang, Z.; Liu, Z. A novel cold-active and salt-tolerant alpha-amylase from marine bacterium Zunongwangia. profunda: Molecular cloning, heterologous expression and biochemical characterization. Extremophiles 2014, 18, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Rabus, R.; Ruepp, A.; Frickey, T.; Rattei, T.; Fartmann, B.; Stark, M.; Bauer, M.; Zibat, A.; Lombardot, T.; Becker, I.; et al. The genome of Desulfotalea. psychrophila, a sulfate-reducing bacterium from permanently cold Arctic sediments. Environ. Microbiol. 2004, 6, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xiao, X.; Wang, P.; Wang, F. Screening and characterization of psychrotrophic lipolytic bacteria from deep-sea sediments. J. Microbiol. Biotechnol. 2004, 14, 952–958. [Google Scholar]
- Carrión, O.; Delgado, L.; Mercade, E. New emulsifying and cryoprotective exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Ivanova, N.; He, Z.; Huebner, M.; Zhou, J.; Tiedje, J. Architecture of thermal adaptation in an Exiguobacterium. sibiricum strain isolated from 3 million year old permafrost: A genome and transcriptome approach. BMC Genom. 2008, 9, 547. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; Mc Cammon, S.A.; Brown, J.L.; McMeekin, T.A. Glaciecola punicea gen. nov., sp. nov. and Glaciecola pallidula gen. nov., sp. nov.: Psychrophilic bacteria from Antarctic sea-ice habitats. Int. J. Syst. Bacteriol. 1998, 48, 1213–1222. [Google Scholar] [CrossRef]
- Reddy, G.S.N.; Raghavan, P.U.M.; Sarita, N.B.; Prakash, J.S.S.; Nagesh, N.; Delille, D.; Shivaji, S. Halomonas glaciei sp. nov. isolated from fast ice of Adelie Land, Antarctica. Extremophiles 2003, 7, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jeong, K.W.; Jin, B.; Ryu, K.S.; Kim, E.H.; Ahn, J.H.; Kim, Y. Structural and dynamic features of cold-shock proteins of Listeria monocytogenes, a psychrophilic bacterium. Biochemistry 2013, 52, 2492–2504. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.C.; Li, H.R.; Xin, Y.H.; Chi, Z.M.; Zhou, P.J.; Yu, Y. Marinobacter psychrophilus sp. nov., a psychrophilic bacterium isolated from the Arctic. Int. J. Syst. Evol. Microbiol. 2008, 58, 1463–1466. [Google Scholar] [CrossRef] [PubMed]
- Franzmann, P.D.; Liu, Y.; Balkwill, D.L. Methanogenium. frigidum sp. nov., a psychrophilic, H2-using methanogen from Ace Lake, Antarctica. Int. J. Syst. Bacteriol. 1997, 47, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, S.; Lee, J.M.; Park, S.; Jung, W.; Kang, J.S.; Joo, H.M.; Seo, K.W.; Kang, S.H. Moritella. dasanensis sp. nov., a psychrophilic bacterium isolated from the Arctic ocean. Int. J. Syst. Evol. Microbiol. 2008, 584, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Mykytczuk, N.C.S.; Foote, S.J.; Omelon, C.R.; Southam, G.; Greer, C.W.; Whyte, L.G. Bacterial growth at 15 °C, molecular insights from the permafrost bacterium Planococcus. halocryophilus Or1. Int. J. Syst. Evol. Microbiol. 2013, 7, 1211–1226. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.S.N.; Matsumoto, G.I.; Schumann, P.; Stackebrandt, E.; Shivaji, S. Psychrophilic pseudomonas from Antarctica: Pseudomonas antarctica sp. nov., Pseudomonas meridiana sp. nov. and Pseudomonas proteolytica sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; Mc Cammon, S.A.; Lewis, T.; Skerratt, J.H.; Brown, J.L.; Nichols, D.S.; McMeekin, T.A. Psychroflexus torquis gen. nov., sp. nov.a psychrophilic species from Antarctic sea ice, and reclassification of Flavobacterium gondwanense (Dobson et al. 1993) as Psychroflexus gondwanense gen. nov., comb. nov. Microbiology 1998, 144, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Jung-Hyun, L.; Ji-Sun, R.; Chiaki, K.; Sang-Jin, K. Shewanella donghaensis sp. nov., a psychrophilic, piezosensitive bacterium producing high levels of polyunsaturated fatty acid, isolated from deep-sea sediments. Int. J. Syst. Evol. Microbiol. 2007, 57, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Morita, N.; Tanaka, M.; Okuyama, H. Biosynthesis of fatty acids in the docosahexaenoic acid-producing bacterium Moritella. marina strain MP-I. Biochem. Soc. Trans. 2000, 28, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Timothy, J.W.; Wilkins, D.; Sheree, Y.; Allen, M.A.; Brown, M.V.; Lauro, F.M.; Cavicchioli, R. Psychrophiles. Annu. Rev. Earth Planet. Sci. 2013, 41, 87–115. [Google Scholar] [CrossRef]
- Jones, P.G.; Inouye, M. The cold-shock response: A hot topic. Mol. Microbiol. 1994, 11, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.; Staley, J.T.; Danchin, A.; Ting, Z.; Brettin, T.S.; Hauser, L.J.; Land, M.L.; Thompson, LS. Genomics of an extreme psychrophile, Psychromonas ingrahamii. BMC Genom. 2008, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, H.; Li, J.; Griffith, M.; Glick, B.R. Relationship between antifreeze protein and freezing resistance in Pseudomonas putida GR12-2. Curr. Microbiol. 2001, 43, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S. Defying the activity-stability trade-off in enzymes: Taking advantage of entropy to enhance activity and thermostability. Crit. Rev. Biotechnol. 2017, 37, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Siddiqui, K.S. David Andrews and Kevin R Sowers. Low-temperature extremophiles and their application. Curr. Opin. Biotechnol. 2002, 13, 253–261. [Google Scholar] [CrossRef]
- Grzymski, J.J.; Carter, B.J.; DeLong, E.F.; Feldman, R.A.; Ghadiri, A.; Murray, A.E. Comparative Genomics of DNA Fragments from Six Antarctic Marine Planktonic Bacteria. Appl. Environ. Microbiol. 2006, 72, 1532–1541. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Wiezer, A.; Strittmatter, A.W.; Daniel, R. Phylogenetic diversity and metabolic potential revealed in a glacier ice metagenome. Appl. Environ. Microbiol. 2009, 75, 7519–7526. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Dornez, E.; Verjans, P.; Arnaut, F.; Delcour, J.A.; Courtin, C.M. Use of psychrophilic xylanases provides insight into the xylanase functionality in bread making. J. Agric. Food Chem. 2011, 59, 9553–9562. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S. Some like it hot, some like it cold: Temperature dependent biotechnological applications and improvements in extremophilic enzymes. Biotechnol. Adv. 2015, 33, 1912–1922. [Google Scholar] [CrossRef] [PubMed]
- Hoyoux, A.; Jennes, I.; Dubois, P.; Genicot, S.; Dubail, F.; Francois, J.M.; Baise, E.; Feller, G.; Gerday, C. Cold-adapted β-galactosidase from the Antarctic psychrophile Pseudoalteromonas. haloplanktis. Appl. Environ. Microbiol. 2001, 67, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, S.; Qiao, L.; Chen, X.; Pang, X.; Shi, M.; Zhang, X.Y.; Qin, Q.L.; Zhou, B.C.; Zhang, Y.Z.; et al. A novel exopolysaccharide from deep-sea bacterium Zunongwangia. profunda SM-A87: Low-cost fermentation, moisture retention, and antioxidant activities. Appl. Microbiol. Biot. 2014, 98, 7437–7445. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Life at high salt conditions. In The Prokaryotes. A Handbook on the Biology of Bacteria: Ecophysiology and Biochemistry; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 2, pp. 263–282. ISBN 978-0-387-30742-8. [Google Scholar]
- DasSarma, S.; DasSarma, P. Halophiles. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar]
- Caumette, P. Ecology and physiology o fphototrophic bacteria and sulphate reducing bacteria in marine salterns. Experientia 1993, 49, 473–481. [Google Scholar] [CrossRef]
- Ma, Y.; Galinski, E.A.; Grant, W.D.; Oren, A.; Ventosa, A. Meeting review. Halophiles Life in Saline Environments. Appl. Environ. Microbiol. 2010, 76, 6971–6981. [Google Scholar] [CrossRef] [PubMed]
- Arahal, D.R.; Ludwig, W.; Schleifer, K.H.; Ventosa, A. Phylogeny of the family Halomonadaceae. based on 23S and 16S rDNA sequence analyses. Int. J. Syst. Evol. Microbiol. 2002, 52, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Shokes, R.F.; Trabant, P.K.; Presley, B.J.; Reid, D.F. Anoxic, hypersaline basin in the Northern Gulf of Mexico. Science 1977, 196, 1443–1446. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Eder, W.; Fareleira, P.; Santos, H.; Huber, R. Salinisphaera. shabanensis gen. nov., sp. nov., a novel, moderately halophilic bacterium from the brine-seawater interface of the Shaban Deep, Red Sea. Extremophiles 2003, 7, 29–34. [Google Scholar] [PubMed]
- Antunes, A.; Taborda, M.; Huber, R.; Moissl, C.; Nobre, M.F.; Da Costa, M.S. Halorhabdus. tiamatea sp. nov., a non-pigmented, extremely halophilic archaeon from a deep-sea, hypersaline anoxic basin of the Red Sea, and emended description of the genus. Halorhabdus. Int. J. Syst. Evol. Microbiol. 2008, 58, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Arahal, D.R.; García, M.T.; Vargas, C.; Cánovas, D.; Nieto, J.J.; Ventosa, A. Chromohalobacter salexigens sp. nov., a moderately halophilic species that includes Halomonas elongata DSM 3043 and ATCC 33174. Int. J. Syst. Evol. Microbiol. 2001, 51, 1457–1462. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, M.; Cabrera, A.; Incerti, C.; Fuentes, S.; Russell, N.J.; Ramos-Cormenzana, A.; Monteoliva-Sánchez, M. Chromohalobacter salarius sp. nov., a moderately halophilic bacterium isolated from a solar saltern in Cabo de Gata, Almeria, southern Spain. Int. J. Syst. Evol. Microbiol. 2007, 57, 1238–1242. [Google Scholar] [CrossRef] [PubMed]
- Mormile, M.R.; Biesen, M.A.; Gutierrez, M.C.; Ventosa, A.; Pavlovich, J.B.; Onstott, T.C.; Fredrickson, J.K. Isolation of Halobacterium. salinarum retrieved directly from halite brine inclusions. Envir. Microbiol. 2003, 5, 1094–1102. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Haloferax. alexandrinus sp. nov., an extremely halophilic canthaxanthin-producing archaeon from a solar saltern in Alexandria (Egypt). Int. J. Syst. Evol. Microbiol. 2002, 52, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.W.; Wu, Y.H.; Wang, C.S.; Oren, A.; Zhou, P.J.; Wu, M. Haloferax. larsenii sp. nov., an extremely halophilic archaeon from a solar saltern. Int. J. Syst. Evol. Microbiol. 2007, 57, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Cui, H.L. Halomicroarcula. salina sp. nov., isolated from a marine solar saltern. Int. J. Syst. Evol. Microbiol. 2015, 65, 1628–1633. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Esposito, E.; Orlando, P.; Lama, L.; Giordano, A.; De Appolonia, F.; Nicolaus, B.; Gambacorta, A. Halomonas. alkaliantarctica sp. nov., isolated from saline lake Cape Russell in Antarctica, an alkalophilic moderately halophilic, exopolysaccharide-producing bacterium. Syst. Appl. Microbiol. 2007, 30, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Nicolaus, B.; Denizci, A.A.; Yavuzturk, B.; Kazan, D. Halomonas. smyrnensis sp. nov., a moderately halophilic, exopolysaccharide-producing bacterium. Int. J. Syst. Evol. Microbiol. 2013, 63, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Arahal, D.R.; Castillo, A.M.; Ludwig, W.; Schleifer, K.H.; Ventosa, A. Proposal of Cobetia. marina gen. nov., comb. nov., within the family Halomonadaceae., to include the species Halomonas. marina. Syst. Appl. Microbiol. 2002, 25, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Nicolaus, B.; Lama, L.; Manca, M.C.; Gambacorta, A. Characterization of a haloalkalophilic strictly aerobic bacterium, isolated from Pantelleria island. Syst. Appl. Microbiol. 1996, 19, 326–333. [Google Scholar] [CrossRef]
- Kaye, J.Z.; Márquez, M.C.; Ventosa, A.; Baross, J.A. Halomonas. neptunia sp. nov., Halomonas. sulfidaeris sp. nov., Halomonas. axialensis sp. nov. and Halomonas. hydrothermalis sp. nov.: Halophilic bacteria isolated from deep-sea hydrothermal-vent environments. Int. J. Syst. Evol. Microbiol. 2004, 54, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Jin, L.; Yang, H.C.; Lee, S.T. Halomonas. gomseomensis sp. nov., Halomonas. janggokensis sp. nov., Halomonas. salaria sp. nov. and Halomonas. denitrificans sp. nov., moderately halophilic bacteria isolated from saline water. Int. J. Syst. Evol. Microbiol. 2007, 57, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Ginzburg, M.; Ginzburg, B.Z.; Hochstein, L.I.; Volcani, B.E. Haloarcula marismortui (Volcani) sp. nov., nom. rev., an extremely halophilic bacterium from the Dead Sea. Int. J. Syst. Bacteriol. 1990, 40, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Arahal, D.R.; Dewhirst, F.E.; Paster, B.J.; Volcani, B.E.; Ventosa, A. Phylogenetic analyses of some extremely halophilic archaea isolated from Dead Sea water, determined on the basis of their 16S rRNA sequences. Appl. Environ. Microbiol. 1996, 62, 3779–3786. [Google Scholar] [PubMed]
- Yoon, J.H.; Kang, K.H.; Oh, T.K.; Park, Y.H. Halobacillus. locisalis sp. nov., a halophilic bacterium isolated from a marine solar saltern of the Yellow Sea in Korea. Extremophiles 2004, 8, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium. halobium isolated from a Tunisian solar saltern. BMC Complement. Altern. Med. 2013, 13, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Weisburg, W.G.; Kessel, M.; Woese, C.R. Halobacteroides. halobius gen. nov., sp. nov., a moderately halophilic anaerobic bacterium from the bottom sediments of the Dead Sea. Syst. Appl. Microbiol. 1984, 5, 58–70. [Google Scholar] [CrossRef]
- Hua, N.P.; Kanekiyo, A.; Fujikura, K.; Yasuda, H.; Naganuma, T. Halobacillus. profundi sp. nov. and Halobacillus. kuroshimensis sp. nov., moderately halophilic bacteria isolated from a deep-sea methane cold seep. Int. J. Syst. Evol. Microbiol. 2007, 57, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Liu, Z.X.; Zhang, Y.Q.; Zhang, Y.X.; Tang, S.K.; Borrathybay, E.; Li, W.J.; Cui, X.L. Halobacillus. naozhouensis sp. nov., a moderately halophilic bacterium isolated from a sea anemone. Antonie Leeuwenhoek 2009, 96, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kang, S.J.; Lee, C.H.; Oh, H.W.; Oh, T.K. Halobacillus. yeomjeoni sp. nov., isolated from a marine solar saltern in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 2413–2417. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kang, S.J.; Oh, T.K. Halobacillus. seohaensis sp. nov., isolated from a marine solar saltern in Korea. Int. J. Syst. Evol. Microbiol. 2008, 58, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Shimane, Y.; Hatada, Y.; Minegishi, H.; Mizuki, T.; Echigo, A.; Miyazaki, M.; Ohta, Y.; Usami, R.; Grant, W.D.; Horikoshi, K. Natronoarchaeum. mannanilyticum gen. nov., sp. nov., an aerobic, extremely halophilic archaeon isolated from commercial salt. Int. J. Syst. Evol. Microbiol. 2010, 60, 2529–2534. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Pletschke, B.I. Review of the enzymatic machinery of Halothermothrix. orenii with special reference to industrial applications. Enzyme Microb. Technol. 2014, 55, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Uzyol, K.S.; Sarıyar-Akbulut, B.; Denizci, A.A.; Kazan, D. Thermostable α-amylase from moderately halophilic Halomonas. sp. AAD21. Turk. J. Biol. 2012, 36, 327–338. [Google Scholar] [CrossRef]
- Corral, P.; Gutierrez, M.C.; Castillo, A.M.; Dominguez, M.; Lopalco, P.; Corcelli, A.; Ventosa, A. Natronococcus. roseus sp. nov., a haloalkaliphilic archaeon from a hypersaline lake. Int. J. Syst. Evol. Microbiol. 2013, 63, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Jiang, X.; Xu, Z.; Fang, Y.; Li, L. Characterization of a novel metallo-β-lactamases fold hydrolase from Pelagibacterium. halotolerans, a marine halotolerant bacterium isolated from East China Sea. Extremophiles 2016, 20, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Seth-Pasricha, M.; Bidle, K.A.; Bidle, K.D. Specificity of archaeal caspase activity in the extreme halophile Haloferax volcanii. Environ. Microbiol. Rep. 2013, 5, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pomares, F.; Bautista, V.; Ferrer, J.; Pire, C.; Marhuenda-Egea, F.C.; Bonete, M.J. Alpha-amylase activity from the halophilic archaeon Haloferax. mediterranei. Extremophiles 2003, 7, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Mesbah, N.M.; Wiegel, J. Halophilic alkali- and thermostable amylase from a novel polyextremophilic Amphibacillus. sp. NM- Ra2. Int. J. Biol. Macromol. 2014, 70, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Parolis, H.; Parolis, L.A.S.; Boán, I.F.; Rodríguez-Valera, F.; Widmalm, G.; Manca, M.C.; Jansson, P.E.; Sutherland, I.W. The structure of the exopolysaccharide produced by the halophilic Archaeon Haloferax. mediterranei strain R4 (ATCC 33500). Carbohydr. Res. 1996, 295, 147–156. [Google Scholar] [CrossRef]
- Lee, H.K.; Chun, J.; Moon, E.J.; Ko, S.H.; Lee, D.S.; Lee, H.S.; Bae, K.S. Hahella. chejuensis gen. nov., sp. nov., an extracellular-polysaccharide-producing marine bacterium. Int. J. Sist. Evol. Microbiol. 2001, 51, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Poli, A.; Finore, I.; Huertas, F.J.; Gambacorta, A.; Pelliccione, S.; Nicolaus, G.; Lama, L.; Nicolaus, B. Haloterrigena hispanica sp. nov., an extremely halophilic archaeon from Fuente de Piedra, southern Spain. Int. J. Syst. Evol. Microbiol. 2007, 57, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Finore, I.; Nicolaus, G.; Huertas, F.J.; Lama, L.; Nicolaus, B.; Poli, A. Halobacillus. alkaliphilus sp. nov., a halophilic bacterium isolated from a salt lake in Fuente de Piedra, southern Spain. Int. J. Syst. Evol. Microbiol. 2008, 58, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Carillo, S.; Pieretti, G.; Lindner, B.; Romano, I.; Nicolaus, B.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. The Lipid A from the Haloalkaliphilic Bacterium Salinivibrio. sharmensis Strain BAGT. Mar. Drugs 2013, 11, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Özturk, H.U.; Sariyar Akbulut, B.; Ayan, B.; Poli, A.; Denizci, A.A.; Utkan, G.; Nicolaus, B.; Kazan, D. Moderately Halophilic Bacterium Halomonas. sp. AAD12: A promising candidate as a Hydroxyectoine Producer. J. Microbiol. Biochem. Technol. 2015, 7, 262–268. [Google Scholar] [CrossRef]
- Siglioccolo, A.; Paiardini, A.; Piscitelli, M.; Pascarella, S. Structural adaptation of extreme halophilic proteins through decrease of conserved hydrophobic contact surface. BMC Struct. Biol. 2011, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Eichler, J. Facing extremes: Archaeal surface-layer (glyco)proteins. Microbiology 2003, 149, 3347–3351. [Google Scholar] [CrossRef] [PubMed]
- Ghai, R.; Pašić, L.; Fernández, A.B.; Martin-Cuadrado, A.B.; Megumi Mizuno, C.; McMahon, K.D.; Papke, R.T.; Stepanauskas, R.; Rodriguez-Brito, B.; Rohwer, F.; Sánchez-Porro, C.; Ventosa, A.; Rodríguez-Valera, F. New Abundant Microbial Groups in Aquatic Hypersaline Environments. Sci. Rep. 2011, 1, 135. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sánchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. Prokaryotic taxonomic and metabolic diversity of an intermediate salinity hipersaline habitat assessed by metagenomics. FEMS Microbiol. Ecol. 2014, 88, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, S.; Yilan, G.; Akbulut, B.S.; Poli, A.; Kazan, D. Interplay of adaptive capabilities of Halomonas. sp. AAD12 under salt stress. J. Biosci. Bioeng. 2012, 114, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Narasingarao, P.; Podell, S.; Ugalde, J.A.; Brochier-Armanet, C.; Emerson, J.B.; Brocks, J.J.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. De novo metagenomic assembly reveals abundant novel major lineage of archaea in hypersaline microbial communities. Int. J. Syst. Evol. Microbiol. 2011, 6, 81–93. [Google Scholar] [CrossRef]
- Coronado, M.J.; Vargas, C.; Hofemeister, J.; Ventosa, A.; Nieto, J.J. Production and biochemical characterization of an α-amylase from the moderate halophile Halomonas. meridiana. FEMS Microbiol. Lett. 2000, 183, 67–71. [Google Scholar] [CrossRef]
- Lama, L.; Romano, I.; Calandrelli, V.; Nicolaus, B.; Gambacorta, A. Purification and characterization of a protease produced by an aerobic haloalkaliphilic species belonging to the Salinivibrio. genus. Res. Microbiol. 2005, 156, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Purohit, M.K.; Raval, V.H.; Pandey, S.; Akbari, V.G.; Rawal, C.M. Capturing the potential of Haloalkaliphilic bacteria from the saline habitats through culture dependent and metagenomic approaches. Current Research, Technology and Education Topics. In Applied Microbiology and Microbial Biotechnology; Méndez-Vilas, A., Ed.; Format Research Centre: Badajoz, Spain, 2010; pp. 81–87, ISBN-13: 978-84-614-6195-0. [Google Scholar]
- Moreno, M. de L.; Màrquez, M.C.; Garcìa, M.T.; Mellado, E. Halophilic bacteria and archaea as producers of lipolytic enzymes. In Biotechnology of Extremophiles. Advances and challenges; Rampelotto, P.H., Ed.; Springer International publishing: Switzerland, 2016; pp. 375–397. ISBN 978-3-319-13520-5. [Google Scholar]
- Margesin, R.; Schinner, F. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles 2001, 5, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Bestvater, T.; Louis, P.; Galinski, E.A. Heterologous ectoine production in Escherichia coli: By-passing the metabolic bottle-neck. Saline Syst. 2008, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Ollivier, B.; Caumette, P.; Garcia, J.L.; Mah, R.A. Anaerobic bacteria from hypersaline environments. Microbiol. Rev. 1994, 58, 27–38. [Google Scholar] [PubMed]
- Xu, Y.; Wang, Z.; Xue, Y.; Zhou, P.; Ma, Y.; Ventosa, A.; Grant, W.D. Natrialba. hulunbeirensis sp. nov. and Natrialba. chahannaoensis sp. nov., novel haloalkaliphilic archaea from soda lakes in Inner Mongolia Autonomous Region, China. Int. J. Syst. Evol. Microbiol. 2001, 51, 1693–1698. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.J.; Sarmiento, B.F.; Mesbah, N.M.; Wiegel, J. Natronolimnobius ‘aegyptiacus’, a novel poly-extremophilic archaeon isolated from athalassohaline lakes in the Wadi An Natrun, Egypt. Proceedings of Abs SE Branch Annual Meeting Am Soc Microbiol, Savannah, GA, USA, 12–15 September 2009. [Google Scholar]
- Xu, Y.; Zhou, P.; Tian, X. Characterization of two novel haloalkaliphilic archaea, Natronorubrum. bangense gen. nov., sp. nov. and Natronorubrum. tibetense gen. nov., sp. nov. Int. J. Syst. Bacteriol. 1999, 49, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Mesbah, N.M.; Hedrick, D.B.; Peacock, A.D.; Rohde, M.; Wiegel, J. Natranaerobius. thermophilus gen. nov., sp. nov., a halophilic, alkalithermophilic bacterium from soda lakes of the Wadi An Natrun, Egypt, and proposal of Natranaerobiaceae. fam. Nov. and Natranaerobiales. ord. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2507–2512. [Google Scholar] [CrossRef] [PubMed]
- Mesbah, N.M.; Wiegel, J. Natronovirga. wadinatrunensis gen. nov., sp. nov. and Natranaerobius. trueperi sp. nov., two halophilic, alkalithermophilic microorganisms from soda lakes of the Wadi An Natrun, Egypt. Int. J. Syst. Evol. Microbiol. 2009, 59, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F.; Trüper, H.G. Ectothiorhodospira. halochloris sp. nov., a new extremely halophilic phototrophic bacterium containing bacteriochlorophyll b. Arch. Microbiol. 1977, 114, 114–121. [Google Scholar] [CrossRef]
- Bowers, K.J.; Mesbah, N.M.; Wiegel, J. Natranaerobius. 'grantii' and Natranaerobius. 'jonesii', spp. nov., two anaerobic halophilic alkaliphiles isolated from the Kenyan-Tanzanian Rift [abstract]. Proceedings of Abst. Gen. Meet Am. Soc. Microbiol., Boston, MA, USA, 25–28 October 2008. I-007. [Google Scholar]
- Kumar, V.; Satyanarayana, T. Thermo-alkali-stable xylanase of a novel polyextremophilic Bacillus halodurans TSEV1 and its application in biobleaching. Int. Biodeter. Biodegr. 2012, 75, 138–145. [Google Scholar] [CrossRef]
- Kumar, V.; Satyanarayana, T. Generation of xylooligosaccharides from microwave irradiated agroresidues using recombinant thermo-alkali-stable endoxylanase of the polyextremophilic bacterium Bacillus halodurans expressed in Pichia. pastoris. Bioresour. Technol. 2015, 179, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Vijayalaxmi, S.; Prakash, P.; Jayalakshmi, S.K.; Mulimani, V.H.; Sreeramulu, K. Production of extremely alkaliphilic, halotolerent, detergent, and thermostable mannanase by the free and immobilized cells of Bacillus halodurans PPKS-2. Purification and characterization. Appl. Biochem. Biotechnol. 2013, 171, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Karan, R.; Capes, M.D.; DasSarma, P.; DasSarma, S. Cloning, overexpression, purification, and characterization of a polyextremophilic beta-galactosidase from the Antarctic haloarchaeon Halorubrum. lacusprofundi. BMC Biotechnol. 2013, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Mol. Biol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Cowan, D.A.; Ramond, J.B.; Makhalanyane, T.P.; De Maayer, P. Metagenomics of extreme environments. Curr. Opin. Microbiol. 2015, 25, 97–102. [Google Scholar] [CrossRef] [PubMed]

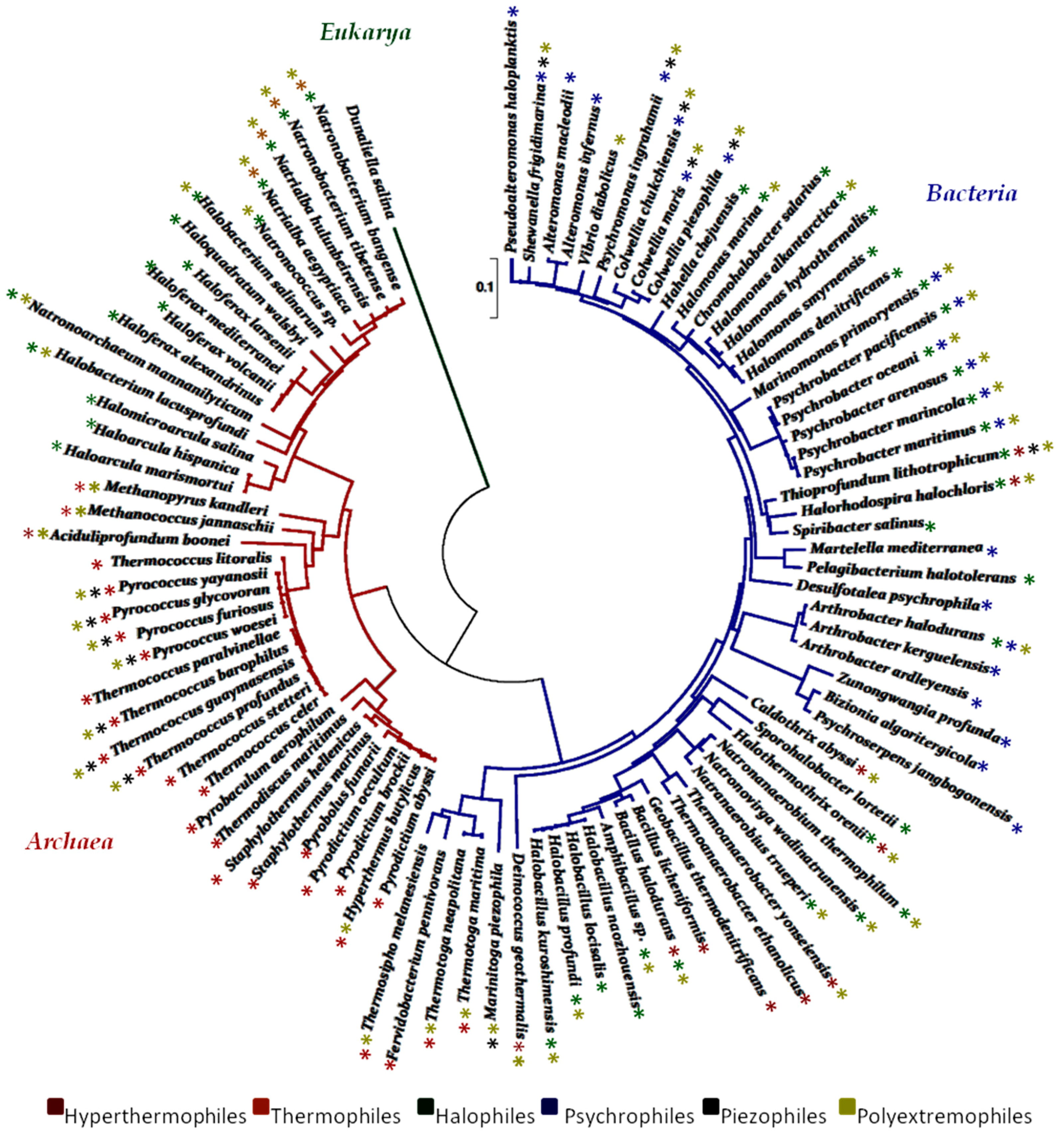

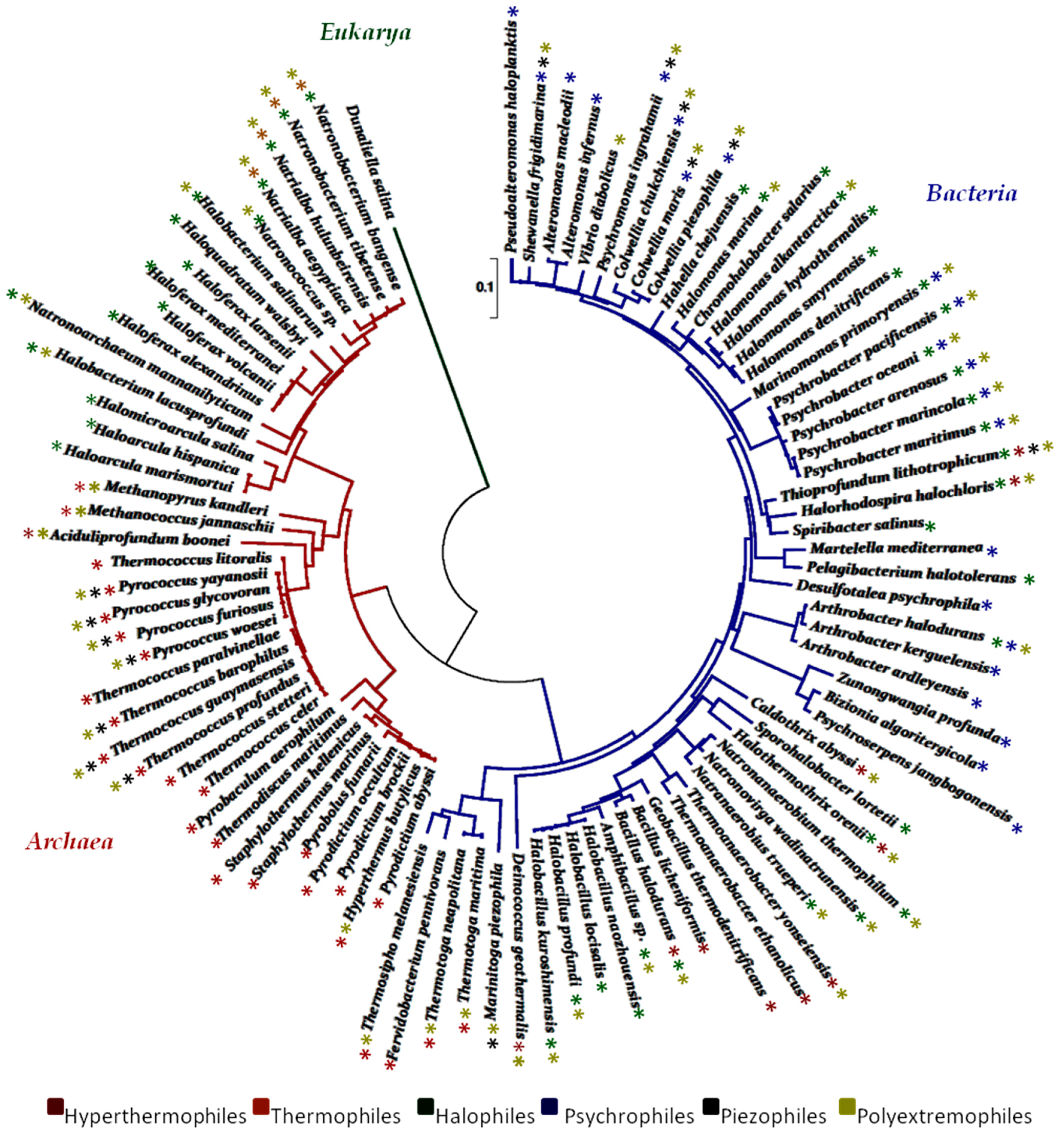
{kind=link}
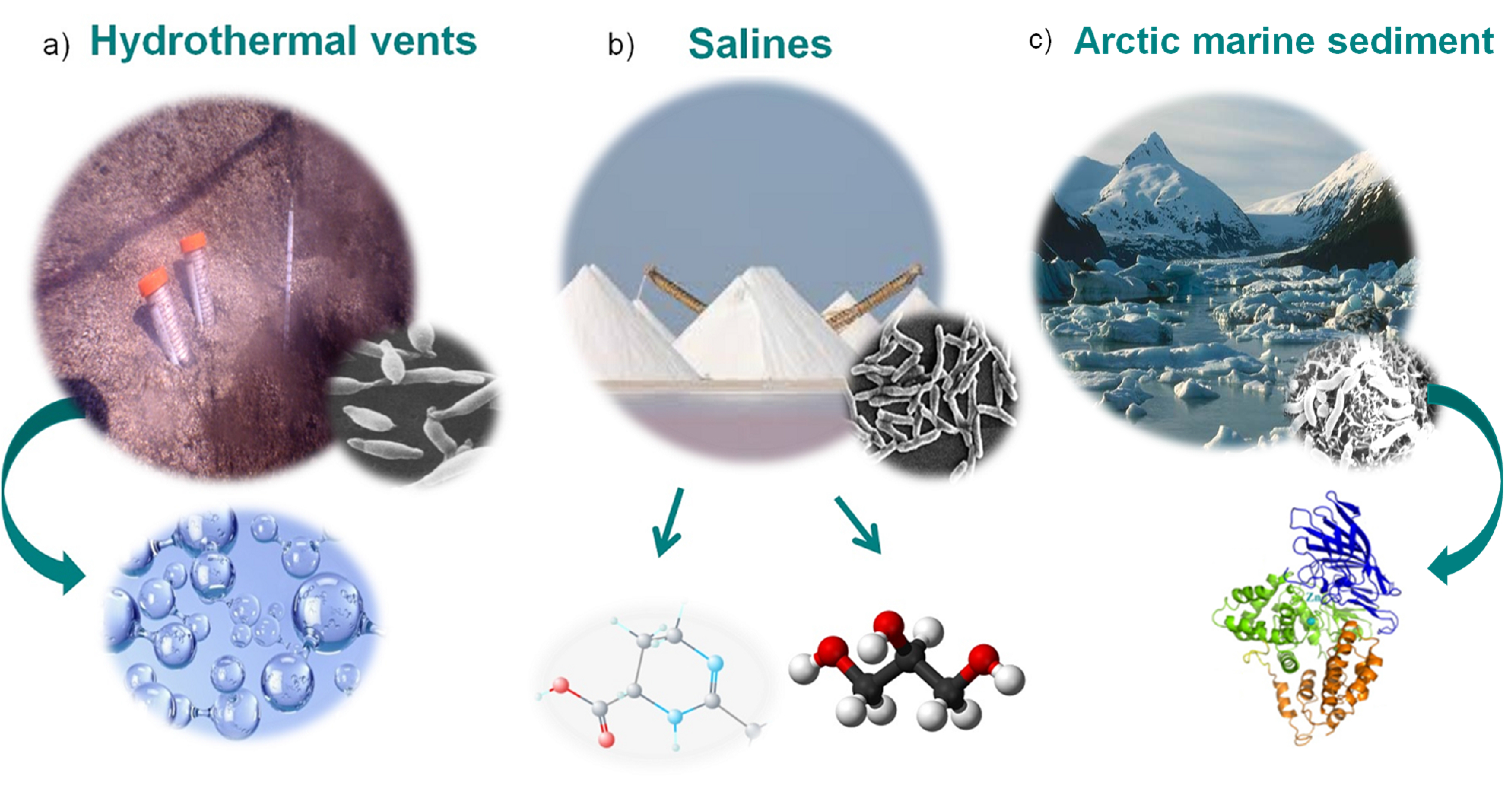
{kind=link}
| Microorganisms | Growth Metabolism | Site and Temperature of Isolation | Reference |
|---|---|---|---|
| Pyrodictium occultum | H₂+ S° ⇒ H₂S | Submarine solfataric field off Vulcano, ItalySSubmarine field off Vulcano, Italy; 105 °C ubmarine solfataric field off Vulcano, Italy; | [30] |
| Pyrodictium abyssi | Organic [H] + S° ⇒ H₂S + CO₂ +organic acids | Marine hot abyssal sites black smokers; 97 °C | [31] |
| Hyperthermus butylicus | Organic [H] + S° ⇒ H₂S + butanol + propionic acid | Hydrothermally heated flat-sea sediments off the coast of Sari Miguel, Azores; 100 °C | [32] |
| Pyrococcus woesei | Organic [H] + S° ⇒ H₂S | Vulcano Island Beach, Italy; 104 °C | [32] |
| Methanopyrus kandleri | H₂ + CO₂ ⇒ CH4 | “Black smoker” chimney, sea floor of the Gulf of California; 98 °C | [24] |
| Pyrolobus fumarii | H₂ + NH3 (or S2O3)−2 ⇒ NH4+1 (or H2S) | “Black smoker”; Mid Atlantic Ridge (depth 3650 m); 106 °C | [26] |
| Staphylothermus marinus | Yeast extract ⇒ H2S + CO2 + organic acids | Beach of Vulcano, Italy, 92 °C | [26] |
| Thermococcus guaymasensis | Organic [H] + S° ⇒ H2S | Guaymas Basin hydrothermal vent, California, 88 °C | [24] |
| Thermococcus barophilus | Organic [H] + S° ⇒ H2S | Hydrothermal vent site on the Mid-Atlantic Ridge; 85 °C | [24] |
| Thermodiscus maritimus | Organic [H] + S° ⇒ H2S | Hot marine sediment at the beach of Vulcano Island; 90 °C | [26] |
| Methanococcus jannaschii | H2 + CO2 ⇒ CH4 | “White smoker” chimney on the East Pacific Rise at 20°50′ N latitude and 109°06′ W longitude at a depth of 2600 m; 88 °C | [26] |
| Thermococcus paralvinellae | Organic [H] + S° ⇒ H₂S | Hydrothermal vent chimneys in the north-eastern Pacific Ocean; 28–88 °C | [33] |
| Pyrococcus furiosus | Organic + S° ⇒ H₂CO₂: + H₂S | Geothermally heated marine sediments at the beach of Porto di Levante, Vulcano, Italy; 100 °C | [26] |
| Microorganisms | Site and Temperature of Isolation | References |
|---|---|---|
| Bacillus thermodenitrificans strain B3-72, Bacillus licheniformis strain B3-15 | Water of a shallow hydrothermal vent, Vulcano Island, Italy | [21] |
| Geobacillus sp. strain 4004 | Sediment in marine hot spring near the seashore of Maronti, Ischia Island, Italy | [39] |
| Hyperthermus butylicus | Hydrothermally heated flat-sea sediments off the coast of Sari Miguel, Azores; 107 °C | [32] |
| Pyrobaculum aerophilum | Maronti Beach, Ischia, Italy; 100 °C | [40] |
| Thermus aquaticus | Thermal springs in Yellowstone National Park and thermal spring in California; 70 °C | [41] |
| Pyrolobus fumarii | “Black smoker”; Mid Atlantic Ridge (depth 3650 m); 106 °C | [26] |
| Pyrococcus glycovorans | Deep-sea hydrothermal vent located on the East Pacific Rise; 95 °C | [42] |
| Pyrococcus yayanosii | Deep-sea hydrothermal vent, Mid-Atlantic Ridge; 98 °C | [43] |
| Pyrodictium abyssi | Marine hot abyssal sites black smokers; 97 °C | [31] |
| Pyrodictium brockii | Submarine solfataric field off Vulcano, Italy; Submarine solfataric field of Vulcano Italy, 105 °C | [30] |
| Staphylothermus hellenicus | Geothermally heated vents off Palaeochori Bay, Milos, Greece; 85 °C | [44] |
| Thermococcus celer | Solfataric marine water hole on a beach of Vulcano, Italy; marine solfataric fields of Kraternaya cove (Ushishir archipelago, Northern Kurils 88 °C | [45] |
| Thermococcus litoralis | Shallow submarine thermal spring from Lucrino and Vulcano in Italy; 88 °C | [38] |
| Thermococcus stetteri | Marine Ushishir archipelago, Northern Kurils solfataricus fields of Kraternaya 75 °C | [46] |
| Thermococcus profundus | Deep-sea hydrothermal vent, Middle Okinawa Trough; 80 °C | [47] |
| Thermococcus aegaeicus | Geothermally heated vents off Palaeochori Bay, Milos, Greece; 88–90 °C | [44] |
| Thermococcus atlanticus | Deep-sea hydrothermal vent in the Mid-Atlantic Ridge; 85 °C | [48] |
| Thermococcus cleftensis | Hydrothermal vent chimneys in the north-eastern Pacific Ocean; 88 °C | [33] |
| Thermococcus nautili | Hydrothermal deep-sea vent, East Pacific Rise; 80 °C | [49] |
| Thermococcus prieurii | Deep-sea hydrothermal vent, East Pacific Rise; 85 °C | [50] |
| Thermococcus thioreducens | Deep-sea hydrothermal vent site on the Mid-Atlantic Ridge; 83–85 °C | [51] |
| Thermotoga maritima | Geothermally heated locales on the sea floor; 80 °C | [52] |
| Microorganisms | Isolation Sites | Applications | Enzyme | Reference |
|---|---|---|---|---|
| Fervidobacterium pennivorans V5 (recombinant) | Hot springs, Azores islands | Starch conversion; detergent | Amylase debranching | [53] |
| Fervidobacterium pennivorans | Hot spring, Azores island | Poultry industry; detergent; fish industry | Serine peptidase | [53] |
| Pyrococcus furiosus | Hydrothermal vent, Italy | Starch saccharification; detergent | Pullulanase | [45] |
| Thermococcus sp. | Deep-sea hydrothermal vent, USA | Starch conversion; detergent | Alpha-amylase | [45] |
| Thermotoga neapolitana 5068 | Hot spring, Italy | Dietary Supplements | Alpha-galactosidase | [54] |
| Thermococcus litoralis | Deep-sea hydrothermal vent, Italy | Detergent | Proline dipeptidase | [55] |
| Thermococcus litoralis | Deep-sea hydrothermal vent Shallow submarine thermal springs and oil wells | Poultry industry; detergent; fish industry | Serine peptidase | [56] |
| Thermoanaerobacter yonseiensis | Geothermal hot stream at Sileri, Indonesia | Poultry industry; detergent; fish industry | Serine protease | [57] |
| Aciduliprofundum boonei | Hydrothermal vent, Mid Atlantic Ridge | Detergent | Cysteine peptidase | [58] |
| Caldithrix abyssi | Deep-sea hydrothermal chimneys, Mid Atlantic Ridge | Detergent | Metallo carboxy-peptidase | [58] |
| Deinococcus geothermalis | Deep-ocean subsurfaces, Italy | Detergent | Thermo-alkali-stable peptidase | [59] |
| Salinivibrio sp. SA-2 | Hypersaline brackish water, Iran | Detergent | Lipase | [60] |
| Microorganisms | EPS-Structure and Chemical Compositions 1 | Applications and Activity | Reference | |
| Bacillus thermodenitrificans strain B3-72 | manno-pyranosidic trisacchacaride repeating unit; Man:Glc Molar ratio:1:0.2 | Immunomodulatory and antiviral activities | [21] | |
| Bacillus licheniformis strain T4 | beta-manno-pyranosidic trisaccharide repeating unit; Fruc/Fuc/Glc/GalNAc/Man Molar ratios: 1.0:0.75:0.28:tr:tr | Anti-citotoxicy activity | [61] | |
| Bacillus licheniformis strain B3-15 | Tetrasaccharide repeating unit; Man manno-pyranosidic configuration | Antiviral activity | [21] | |
| Geobacillus sp. strain 4004 | Gal:Man:GlcN:Ara Molar ratios:1.0:0.8:0.4:02 | Pharmaceutical application | [39] | |
| Thermococcus litoralis | Man | Biofilm formation | [26] | |
| Vibrio diabolicus | GlcNAc and GalNAc Molar ratios:1.0:1.0 | Regenerating activity on bone and skin | [62] | |
| Microorganisms | Site and Temperature of Isolation | References |
|---|---|---|
| Arthrobacter ardleyensi | Antarctic Ardley Island lake sediment; 25 °C | [89] |
| Arthrobacter halodurans | Sea water collected from the South China Sea; 28 °C | [90] |
| Arthrobacter kerguelensis | Sea water, Kerguelen Islands, Antarctica; 22 °C | [91] |
| Arthrobacter subterraneus | Deep subsurface water of the South Coast of Korea; 28 °C | [92] |
| Bizionia algoritergicola | Sea ice–sea water, East Antarctica; −2 °C | [93] |
| Colwellia chukchiensis | Chukchi Sea in the Arctic Ocean; 23–25 °C | [16] |
| Colwellia maris | Seawater, Abashiri coast off the Okhotsuku Sea, Hokkaido; 0–22 °C | [94] |
| Colwellia piezophila | Deep-sea sediments of the Japan Trench; 10 °C | [95] |
| Colwellia psychrerythraea | Sea ice and marine sediments; Arctic. 8 °C | [16,96,97] |
| Pseudoalteromonas haloplanktis | Antarctic coastal sea water; 12 °C | [98] |
| Psychrobacter arenosus, P. marincola, P. maritimus, P. submarinus, P. fulvigenes | Coastal sea ice and sediments of the Sea of Japan; 25–28 °C | [99] |
| Psychroserpens jangbogonensis | Ross Sea in the Southern Ocean, Antarctica; 15 °C | [100] |
| Psychrobacter oceani | Sediment from Pacific Ocean at depth of 7167 m (37°48′ N 143°52′ E); 10–15 °C | [101] |
| Psychrobacter okhotskensis | Monbetsu coast of the Okhotsk Sea in Hokkaido, Japan; 25 °C | [102] |
| Psychrobacter pacificensis | Deep seawater in the Japan Trench off Hachijo Island, Japan; 25 °C | [103] |
| Psychromonas ingrahamii | Sea ice core from Point Barrow, Alaska, USA; −12–10 °C | [104] |
| Shewanella frigidimarina and S. gelidimarina | Coastal areas of the Vestfold Hills in eastern Antarctica (68′S 78′′E) Antarctic sea ice; 15–17 °C | [105] |
| Microorganisms | Isolation Sites | Applications | Enzyme | Reference |
|---|---|---|---|---|
| Pseudoalteromonas haloplanktis | Antarctica | Starch conversion; detergent | Amylase Endo-amylase | [106] |
| Zunongwangia profunda | Deep-sea, China | Starch conversion; detergent | Alpha-amylase | [107] |
| Martelella mediterranea | Lake Martel, Spain | cellulose conversion; detergent | Beta-glucosidase | [22] |
| Desulfotalea psychrophila | Marine sediments, Antarctica | Agriculture; food; pharmaceutical industries | Esterase | [108] |
| Pseudoalteromonas haloplanktis | Marine Antarctic | Detergent | Lipase | [98] |
| Psychrobacter sp. wp37 | Deep-sea sediments, West Pacific | Detergent | Lipase | [109] |
| Microorganisms | EPS-Structure and Chemical Composition 1 | Applications and Activity | Reference | |
| Pseudomonas sp. ID1 | Carbohydrates 33.8% (Glc 17.0%, Gal 8.6%, Fuc 8.2%)/Uronic acids 2.4%/Proteins 2.8% | Cryo-protection and emulsifying activities | [110] | |
| Pseudomonas strain CAM025 | Glc/GalA/Rha/Gal Molar ratios: 1:0.5:0.1:0.08 | Cryo-protection and trace metal binding | [110] | |
| Microorganisms | Site and Temperature of Isolation | References |
|---|---|---|
| Chromohalobacter salexigens | Solar saltern, Bonaire, Netherlands Antilles; 37 °C | [144] |
| Chromohalobacter salarius | Solar saltern Cabo de Gata, Almería, southern Spain; 35 °C | [145] |
| Halobacterium salinarum | Badwater salt pan, Death Valley, California; 37 °C | [146] |
| Haloferax alexandrines | El-Mallahet, solar saltern near Alexandria City in Egypt; 37 °C | [147] |
| Haloferax larsenii | Solar saltern (122°17’ N 29°55’ E), Zhoushan archipelago, Zhe-Jiang, China; 42–45 °C | [148] |
| Halomicroarcula salina | Yinggehai marine solar saltern near Shanya city of Hainan Province, China; 37 °C | [149] |
| Halomonas alkaliantarctica | Saline lake in Cape Russell, Antarctica; 30 °C | [150] |
| Halomonas smyrnensis | Camaltı Saltern Area, Aegean Region of Turkey, 37 °C | [151] |
| Halomonas marina | Marine water (Hawaii, USA); 37 °C and 26 °C | [152] |
| Halomonas pantelleriensis | Hard sand, lake of Venere, Pantelleria Island, Sicily, Italy; 30 °C | [153] |
| Halomonas axialensis, H. hydrothermalis, H. sulfidaeris, H. neptunia | Hydrothermal fluids low-temperature, sulfide rock and hydrothermal plumes in North and South Pacific Ocean vent fields located at 1530–2580 m depth; 20 °C | [154] |
| Halomonas denitrificans, H. salaria, H. janggokensis, H. gomseomensis | Saline water of Gomseom, solar saltern, saline water of Janggok solar saltern, seawater in Anmyeond; Korea; 25–30 °C | [155] |
| Haloarcula marismourti, H. hispanica | Salt lake Dead sea, 31°30′ N, 35°30′ E; 37 °C | [156,157] |
| Halobacillus locisalis | Marine solar saltern, Baekryung Island of the Yellow Sea, Korea; 30 °C | [158] |
| Halobacterium halobium | Solar saltern ,Central-Eastern coast of Tunisia, 34°39′ N and 10°42′ E; 37 °C | [159] |
| Halobacteroides halobius | Salt lake Dead sea sediment ; 37 °C | [160] |
| Halobacillus profundi, H. kuroshimensis | Deep-sea carbonate rock at a methane cold seep in Kuroshima Knoll, Japan; 25 °C | [161] |
| Halobacillus naozhouensis | South China Sea, Naozhou Island on the Leizhou Bay; 30 °C | [162] |
| Halobacillus seohaensis, H. yeomjeoni | Sediment marine solar saltern, Byunsan, Korea; 37 °C | [163,164] |
| Natronoarchaeum mannanilyticum | Japanese seawater in Niigata prefecture; 37 °C | [165] |
| Sporohalobacter lortetii, S. marismortui | Salt lake Dead sea; 35–37 °C | [88] |
| Microorganisms | Isolation Sites | Applications | Enzyme | Reference |
|---|---|---|---|---|
| Halothermothrix orenii | Tunisian salt lake | Starch conversion; detergent | Amylase Endoamylase | [166] |
| Halomonas sp. AAD21 | Saltern area, Turkey | Starch conversion; detergent | Alpha-amylase | [167] |
| Natronococcus sp. | Soda Lake, China | Detergent | Lipase | [168] |
| Pelagibacterium halotolerans | East China Sea | Metallo beta-lactamase | [169] | |
| Haloferax volcanii | Dead Sea, the Great Salt Lake, and oceanic environments with high NaCl | Detergent | Cysteine peptidase | [170] |
| Haloferax mediterranei | Saltern, Spain | Starch conversion; detergent | Endoamylase | [171] |
| Amphibacillus sp. NM-Ra2 | Hypersaline alkaline lake Egypt | Starch conversion; detergent | Gluco-amylo pullulanase | [172] |
| Microorganisms | EPS-Structure and Chemical Composition 1 | Applications and Activity | Reference | |
| Haloferax mediterranei | → 4)-β-D-GlcNAcA-(1→6)-α-D-Man-(1→ 4)-β-D-GlcNAcA-3-O-SO3−-(1→ | Candidate in oil recovery overall in oil deposits with high salinity concentrations | [173] | |
| Halomonas alkaliantarctica strain CRSS | Glc:Fru:GlcNAc:GalNAc Molar ratio:1.0:0.7:0.3:tr | High viscosity | [11] | |
| Hahella chejuensis | Glc:Gal Molar ratio:0.68:1.0 | Biosurfactant and detoxification of polluted areas from petrochemical oils | [174] | |
| Alteromonas infernus | Repeating unit of uronic acids (GlcA and GalA) and neutral sugars (Gal and Glc) and substituted with one sulfate group | Cartilage tissue engineering applications due to “Heparin-like” behavior | [81] | |
| Alteromonas macleodii subsp. fijiensis | Glc/Gal/Man/Rha/Fuc/GlcA/GalA Molar ratio: 1/1.9/0.4/0.6/0.2/1.2/2.8 | Protection of sensitive skin against chemical, mechanical and UVB aggressions | [25] | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. https://doi.org/10.3390/microorganisms5020025
Poli A, Finore I, Romano I, Gioiello A, Lama L, Nicolaus B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms. 2017; 5(2):25. https://doi.org/10.3390/microorganisms5020025
Chicago/Turabian StylePoli, Annarita, Ilaria Finore, Ida Romano, Alessia Gioiello, Licia Lama, and Barbara Nicolaus. 2017. "Microbial Diversity in Extreme Marine Habitats and Their Biomolecules" Microorganisms 5, no. 2: 25. https://doi.org/10.3390/microorganisms5020025
APA StylePoli, A., Finore, I., Romano, I., Gioiello, A., Lama, L., & Nicolaus, B. (2017). Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms, 5(2), 25. https://doi.org/10.3390/microorganisms5020025