Mapping Epitopes of a Novel Peptidoglycan Cross-Linking Enzyme Cwp22 Recognized by Human Sera Obtained from Patients with Clostridioides difficile Infection and Cord Blood

 , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Blood Sera
2.1.1. Human Peripheral Blood Sera
2.1.2. Human Umbilical Cord Blood Sera
2.2. Bacterial Strains
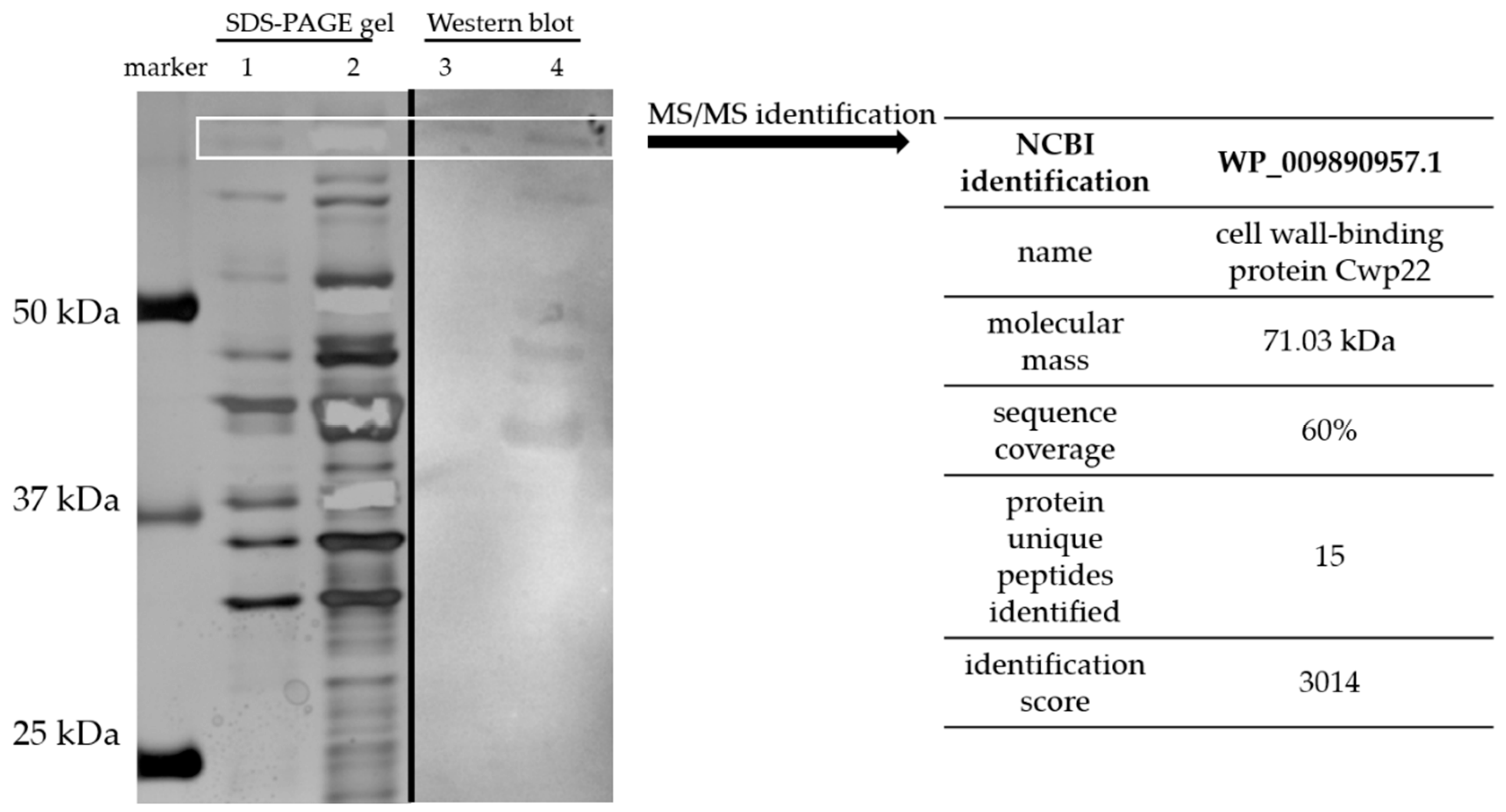
2.3. Protein Isolation
2.4. SDS-PAGE Electrophoresis
2.5. Silver Staining
2.6. Western Blotting
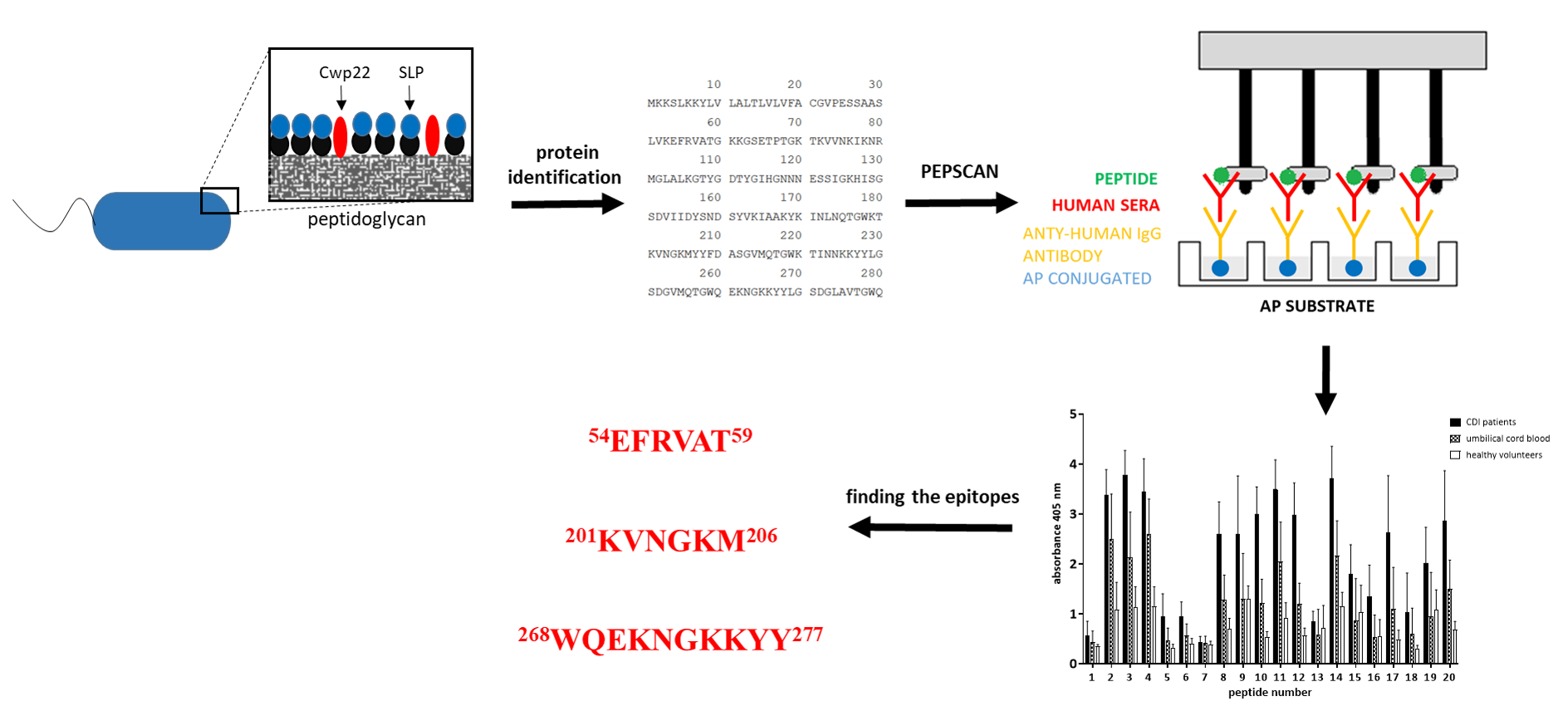
2.7. Protein Identification
2.8. Analysis of Sequence Homology
2.9. Protein Modeling
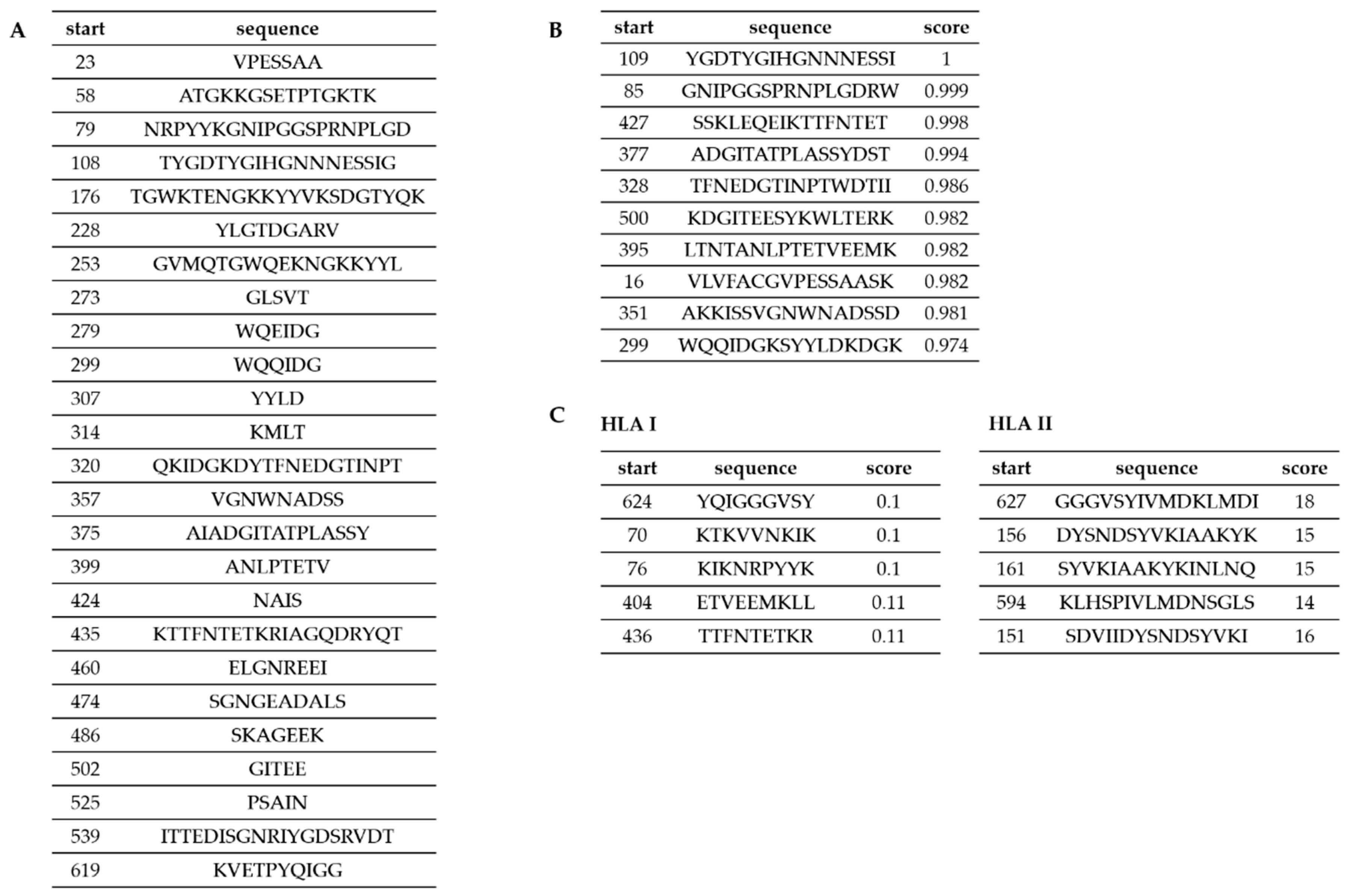
2.10. Predicting Epitopes
2.11. Chemical Synthesis of Peptides
2.12. Pin Enzyme-Linked Immunosorbent Assay
2.13. Epitope Identification
2.14. Epitope Analysis
3. Results
3.1. The Identification of Cwp22 as a New Immunoreactive CD Surface Protein
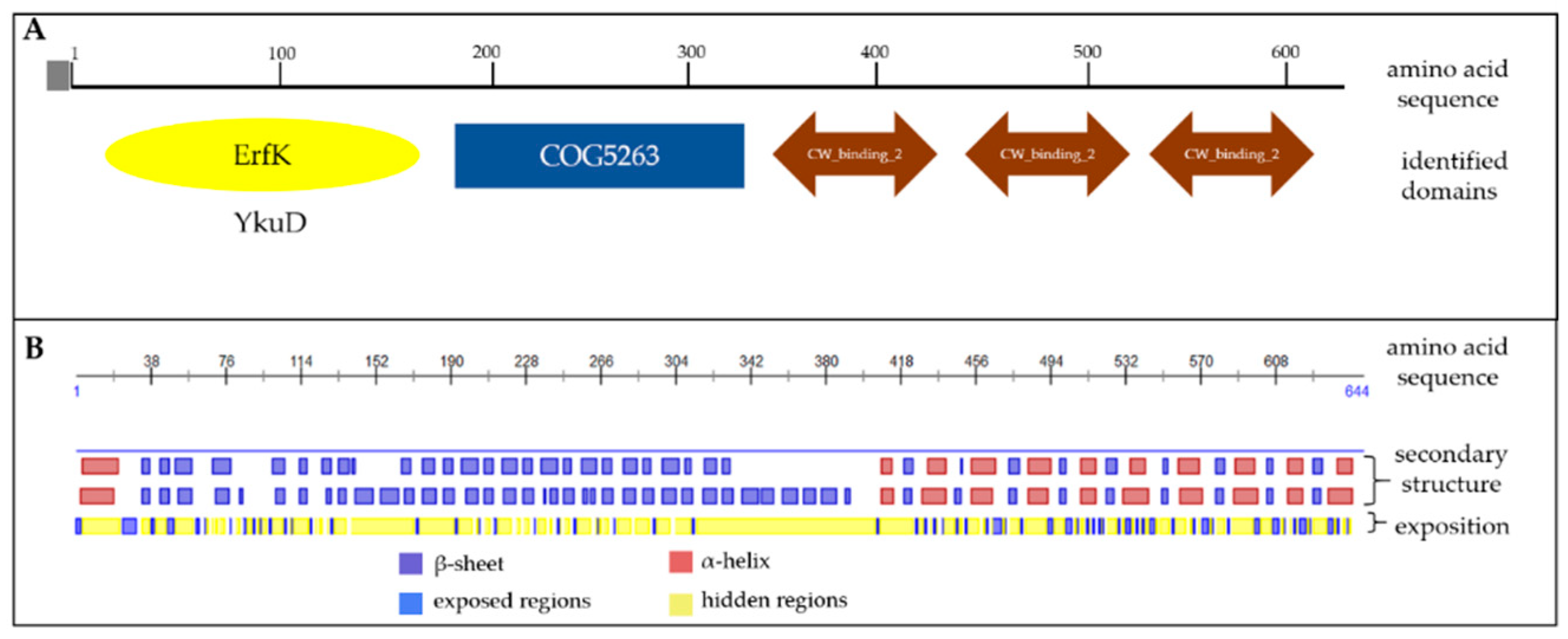
3.2. In Silico Analysis Shows that Cwp22 Protein is a Good Vaccine Antigen Candidate
3.3. Predicting the Epitopes of Cwp22 Protein
3.4. Epitope Mapping
3.5. Defining the Epitopes of Cwp22 Protein
3.6. Epitopes Characteristics
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lawson, P.A.; Citron, D.M.; Tyrrell, K.L.; Finegold, S.M. Reclassification of Clostridium difficile as Clostridioides difficile (Hall and O’Toole 1935) prévot 1938. Anaerobe 2016, 40, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Barbut, F.; Petit, J.C. Epidemiology of Clostridium difficile-associated infections. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2001, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Antonopoulos, D.A.; Kalra, A.; Tonelli, A.; Khalife, W.T.; Schmidt, T.M.; Young, V.B. Decreased diversity of the fecal microbiome in recurrent Clostridium Difficile—Associated diarrhea. J. Infect. Dis. 2008, 197, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P. Can we identify patients at high risk of recurrent Clostridium difficile infection? Predictors of C. difficile recurrence. Clin. Microbiol. Infect. 2012, 18, 21–27. [Google Scholar] [CrossRef]
- Thelestam, M.; Chaves-Olarte, E. Cytotoxic effects of the Clostridium difficile Toxins. In Clostridium Difficile; Aktories, K., Wilkins, T.D., Eds.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2000; pp. 85–96. ISBN 978-3-662-06272-2. [Google Scholar]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium difficile infection: new developments in epidemiology and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical practice guidelines for Clostridium difficile infection in adults and children: 2017 update by the Infectious diseases society of America (IDSA) and Society for healthcare epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C. Antibiotics and hospital-acquired Clostridium Difficile-Associated diarrhoea: A systematic review. J. Antimicrob. Chemother. 2003, 51, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, G.; Saleh, J.; Workman, D.; Pollak, R.; Elinoff, V.; Fraser, N.J.; Lefebvre, G.; Martens, M.; Mills, R.E.; Nathan, R.; et al. Defining the optimal formulation and schedule of a candidate toxoid vaccine against Clostridium difficile infection: A randomized Phase 2 clinical trial. Vaccine 2016, 34, 2170–2178. [Google Scholar] [CrossRef]
- Greenberg, R.N.; Marbury, T.C.; Foglia, G.; Warny, M. Phase I dose finding studies of an adjuvanted Clostridium difficile toxoid vaccine. Vaccine 2012, 30, 2245–2249. [Google Scholar] [CrossRef]
- Bruxelle, J.F.; Péchiné, S.; Collignon, A. Immunization strategies against Clostridium difficile. In Updates on Clostridium Difficile in Europe: Advances in Microbiology, Infectious Diseases and Public Health Volume 8; Mastrantonio, P., Rupnik, M., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2018; pp. 197–225. ISBN 978-3-319-72799-8. [Google Scholar]
- Péchiné, S.; Bruxelle, J.F.; Janoir, C.; Collignon, A. Targeting Clostridium difficile surface components to develop immunotherapeutic strategies against Clostridium difficile Infection. Front. Microbiol. 2018, 9, 1009. [Google Scholar] [CrossRef]
- Zhu, D.; Bullock, J.; He, Y.; Sun, X. Cwp22, A novel peptidoglycan Cross-Linking enzyme, plays pleiotropic roles in Clostridioides difficile. Environ. Microbiol. 2019, 28, 3076–3090. [Google Scholar] [CrossRef] [PubMed]
- Razim, A.; Pacyga, K.; Aptekorz, M.; Martirosian, G.; Szuba, A.; Pawlak-Adamska, E.; Brzychczy-Włoch, M.; Myc, A.; Gamian, A.; Górska, S. Epitopes identified in GAPDH from Clostridium difficile recognized as common antigens with potential autoimmunizing properties. Sci. Rep. 2018, 8, 13946. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Wait, R.; Begum, S.; Crossett, B.; Nagy, J.; Brown, K.; Fairweather, N. Proteomic analysis of cell surface proteins from Clostridium difficile. Proteomics 2005, 5, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Pantosti, A.; Cerquetti, M.; Viti, F.; Ortisi, G.; Mastrantonio, P. Immunoblot analysis of serum immunoglobulin G response to surface proteins of Clostridium difficile in patients with Antibiotic-Associated diarrhea. J. Clin. Microbiol. 1989, 27, 2594–2597. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Rost, B.; Yachdav, G.; Liu, J. The PredictProtein server. Nucleic Acids Res. 2004, 32, W321–W326. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving Sequence-Based B-Cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef]
- EL-Manzalawy, Y.; Dobbs, D.; Honavar, V. Predicting linear B-cell epitopes using string kernels. J. Mol. Recognit. JMR 2008, 21, 243–255. [Google Scholar] [CrossRef]
- Carter, J.M. Epitope Mapping of a protein using the Geysen (PEPSCAN) procedure. In Peptide Analysis Protocols; Humana Press: New Jersey, NJ, USA, 2014; Volume 36, pp. 207–224. ISBN 0-89603-274-4. [Google Scholar]
- Jarząb, A.; Witkowska, D.; Ziomek, E.; Dąbrowska, A.; Szewczuk, Z.; Gamian, A. Shigella flexneri 3a outer membrane protein C epitope is recognized by human umbilical cord sera and associated with Protective activity. PLoS ONE 2013, 8, e70539. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Overton, J.A.; Greenbaum, J.A.; Ponomarenko, J.; Clark, J.D.; Cantrell, J.R.; Wheeler, D.K.; Gabbard, J.L.; Hix, D.; Sette, A.; et al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 2015, 43, D405–D412. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, L.; Zhang, P.; LaBaer, J.; Hermjakob, H.; Li, D.; Yu, X. AAgAtlas 1.0: A human autoantigen database. Nucleic Acids Res. 2017, 45, D769–D776. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Drudy, D.; Kyne, L.; Brown, K.; Fairweather, N.F. Immunoreactive cell wall proteins of Clostridium difficile identified by human sera. J. Med. Microbiol. 2008, 57, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Biazzo, M.; Cioncada, R.; Fiaschi, L.; Tedde, V.; Spigaglia, P.; Mastrantonio, P.; Pizza, M.; Barocchi, M.A.; Scarselli, M.; Galeotti, C.L. Diversity of cwp loci in clinical isolates of Clostridium difficile. J. Med. Microbiol. 2013, 62, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Peltier, J.; Courtin, P.; El Meouche, I.; Lemée, L.; Chapot-Chartier, M.P.; Pons, J.L. Clostridium difficile has an original peptidoglycan structure with a high level of N-acetylglucosamine deacetylation and mainly 3-3 cross-links. J. Biol. Chem. 2011, 286, 29053–29062. [Google Scholar] [CrossRef]
- Bradshaw, W.J.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. The structure of the S-layer of Clostridium difficile. J. Cell Commun. Signal. 2018, 12, 319–331. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Warmerdam, P.A.M.; Plaisance, S.; Vanderlick, K.; Vandervoort, P.; Brepoels, K.; Collen, D.; Maeyer, M.D. Elimination of a Human T-cell Region in Staphylokinase by T-cell Screening and Computer Modeling. Thromb. Haemost. 2002, 87, 666–673. [Google Scholar]
- Palmeira, P.; Quinello, C.; Silveira-Lessa, A.L.; Zago, C.A.; Carneiro-Sampaio, M. IgG placental transfer in healthy and pathological pregnancies. Clin. Dev. Immunol. 2012, 2012, 985646. [Google Scholar] [CrossRef]
- Mäntyjärvi, R.; Hirvonen, T.; Toivanen, P. Maternal antibodies in human neonatal sera. Immunology 1970, 18, 449–451. [Google Scholar] [PubMed]
- Fagan, R.P.; Janoir, C.; Collignon, A.; Mastrantonio, P.; Poxton, I.R.; Fairweather, N.F. A proposed nomenclature for cell wall proteins of Clostridium difficile. J. Med. Microbiol. 2011, 60, 1225–1228. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ternan, N.G.; Jain, S.; Graham, R.L.J.; McMullan, G. Semiquantitative analysis of clinical heat stress in Clostridium difficile strain 630 using a GeLC/MS workflow with emPAI quantitation. PLOS ONE 2014, 9, e88960. [Google Scholar] [CrossRef] [PubMed]
- Pechine, S.; Janoir, C.; Collignon, A. Variability of Clostridium difficile surface proteins and specific serum antibody response in patients with Clostridium difficile-Associated Disease. J. Clin. Microbiol. 2005, 43, 5018–5025. [Google Scholar] [CrossRef]
- Pechine, S. Immunological properties of surface proteins of Clostridium difficile. J. Med. Microbiol. 2005, 54, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Lavollay, M.; Arthur, M.; Fourgeaud, M.; Dubost, L.; Marie, A.; Veziris, N.; Blanot, D.; Gutmann, L.; Mainardi, J.L. The peptidoglycan of stationary-Phase Mycobacterium tuberculosis predominantly contains Cross-Links generated by l,d-Transpeptidation. J. Bacteriol. 2008, 190, 4360–4366. [Google Scholar] [CrossRef]
- Sütterlin, L.; Edoo, Z.; Hugonnet, J.E.; Mainardi, J.L.; Arthur, M. Peptidoglycan Cross-Linking activity of l,d-Transpeptidases from Clostridium difficile and inactivation of these enzymes by β-Lactams. Antimicrob. Agents Chemother. 2018, 62, e01607–e01617. [Google Scholar] [CrossRef]
- Triboulet, S.; Dubée, V.; Lecoq, L.; Bougault, C.; Mainardi, J.-L.; Rice, L.B.; Ethève-Quelquejeu, M.; Gutmann, L.; Marie, A.; Dubost, L.; et al. Kinetic features of L,D-Transpeptidase inactivation critical for β-Lactam antibacterial activity. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Kao, D.J.; Hodges, R.S. Advantages of a synthetic peptide immunogen over a protein immunogen in the development of an Anti-Pilus vaccine for Pseudomonas aeruginosa. Chem. Biol. Drug Des. 2009, 74, 33–42. [Google Scholar] [CrossRef]
- Andersen, P.H.; Nielsen, M.; Lund, O. Prediction of residues in discontinuous B-cell epitopes using protein 3D structures. Protein Sci. 2006, 15, 2558–2567. [Google Scholar] [CrossRef]
- Atassi, M.Z.; Pai, R.C. Immunochemistry of sperm-whale myoglobin—XXII: Accurate delineation of the single reactive region in sequence 103–120 by immunochemical studies of synthetic peptides: The complete antigenic structure of the protein. Immunochemistry 1975, 12, 735–740. [Google Scholar] [CrossRef]
- Pyclik, M.; Górska, S.; Brzozowska, E.; Dobrut, A.; Ciekot, J.; Gamian, A.; Brzychczy-Włoch, M. Epitope mapping of Streptococcus agalactiae elongation factor Tu protein recognized by human sera. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Hancock, D.C.; OReilly, N.J. Synthetic peptides as antigens for antibody production. In Immunochemical Protocols; Burns, R., Ed.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2005; pp. 13–25. ISBN 978-1-59259-873-1. [Google Scholar]
- Pruksakorn, S.; Currie, B.; Brandt, E.; Phornphutkul, C.; Hunsakunachal, S.; Manmontri, A.; Robinson, J.H.; Kehoe, M.A.; Galbraith, A.; Good, M.F. Identification of T cell autoepitopes that Cross-React with the C-Terminal segment of the M protein of group A streptococci. Int. Immunol. 1994, 6, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Williams, W.B.; Liao, H.X.; Moody, M.A.; Kepler, T.B.; Alam, S.M.; Gao, F.; Wiehe, K.; Trama, A.M.; Jones, K.; Zhang, R.; et al. Diversion of HIV-1 vaccine–induced immunity by Gp41-Microbiota Cross-Reactive antibodies. Science 2015, 349, aab1253. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NCBI Identification | CD Strain | Characteristics | Sequence Coverage | % of Identical Amino Acids |
|---|---|---|---|---|
| CBA64897.1 | CD196 | Isolated from an adult, ribotype 027, producing toxins | 100% | 99.84 |
| PBF93815.1 | 7468-NonSp/ST97 | Isolated from a child, producing toxins | 100% | 99.69 |
| EQG56151.1 | DA00145 | 100% | 99.69 | |
| EQH98359.1 | F314 | Acute CDI, producing toxins | 100% | 99.69 |
| OFU21376.1 | HMSC19C09 | 100% | 99.69 | |
| OFU05949.1 | HMSC19D07 | 100% | 99.53 | |
| EHJ35368.1 | 70-100-2010 | 100% | 99.38 | |
| PBG98735.1 | 7481-NonSp/ST59 | 100% | 99.22 | |
| EQF50178.1 | CD174 | Asymptomatic carriage, no toxins | 100% | 99.07 |
| EQE30253.1 | CD34 | Asymptomatic carriage, no toxins | 100% | 99.22 |
| AFV69537.1 | C192 | Ribotype 012, hipervirulent | 100% | 99.22 |
| EQJ05755.1 | P7 | Reference strain, producing toxins | 100% | 99.07 |
| PBE99335.1 | 5573-CF/ST37 | Producing TcdB | 100% | 98.76 |
| EQE84199.1 | CD69 | Acute CDI, producing toxins | 100% | 98.91 |
| OJT75205.1 | NT422 | Ribotype 231 | 98% | 99.37 |
| EFH06251.1 | NAP08 | Hipervirulent | 100% | 96.89 |
| PBI20744 | 6615-NonSp/ST11 | Isolated from a child, hipervirulent | 100% | 96.74 |
| KPI50551 | RA09-70 | Producing TcdA | 100% | 93.17 |
| PBE36357.1 | 5537-D/ST9 | Isolated from a child, producing toxins | 100% | 91.93 |
| ERM37167 | P64 | Asymptomatic carriage, producing toxins | 79% | 99.41 |
| EQE47488 | CD42 | Acute CDI, producing toxins | 73% | 99.30 |
| EQK74493 | CD113 | Relapse, producing toxins | 72% | 99.51 |
| NCBI Identification | Protein Name | Bacteria Strain | Sequence Coverage | % of Identical Aminoacids |
|---|---|---|---|---|
| WP_092726434.1 | l,d-transpeptidase | Romboutsia lituseburensis | 53% | 56.07 |
| WP_007287442.1 | l,d-transpeptidase | Clostridium bartletti | 53% | 51.59 |
| WP_084157842.1 | Hipothetical protein | Bacillus manliponensis | 58% | 47.59 |
| WP_082007829.1 | Hipothetical protein | Terrisporobacter othiniensis | 54% | 52.10 |
| WP_052404759.1 | l,d-transpeptidase | Bacillus rubiinfantis | 55% | 48.66 |
| WP_066389794.1 | l,d-transpeptidase | Bacillus mesonae | 52% | 41.90 |
| KLR55379.1 | Surface protein | Clostridium sordelli | 53% | 45.98 |
| 54EFRVAT59 | 201KVNGKM207 | 268WQEKNGKKYY277 | |
|---|---|---|---|
| Molecular mass (g/mol) | 721.8 | 675.84 | 1343.49 |
| Water solubility | good | good | good |
| pI | 6.86 | 10.69 | 9.9 |
| Charge in physiological pH | 0 | +2 | +2 |
| Peptide structure |  | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razim, A.; Pacyga, K.; Martirosian, G.; Szuba, A.; Gamian, A.; Myc, A.; Górska, S. Mapping Epitopes of a Novel Peptidoglycan Cross-Linking Enzyme Cwp22 Recognized by Human Sera Obtained from Patients with Clostridioides difficile Infection and Cord Blood. Microorganisms 2019, 7, 565. https://doi.org/10.3390/microorganisms7110565
Razim A, Pacyga K, Martirosian G, Szuba A, Gamian A, Myc A, Górska S. Mapping Epitopes of a Novel Peptidoglycan Cross-Linking Enzyme Cwp22 Recognized by Human Sera Obtained from Patients with Clostridioides difficile Infection and Cord Blood. Microorganisms. 2019; 7(11):565. https://doi.org/10.3390/microorganisms7110565
Chicago/Turabian StyleRazim, Agnieszka, Katarzyna Pacyga, Gajane Martirosian, Andrzej Szuba, Andrzej Gamian, Andrzej Myc, and Sabina Górska. 2019. "Mapping Epitopes of a Novel Peptidoglycan Cross-Linking Enzyme Cwp22 Recognized by Human Sera Obtained from Patients with Clostridioides difficile Infection and Cord Blood" Microorganisms 7, no. 11: 565. https://doi.org/10.3390/microorganisms7110565
APA StyleRazim, A., Pacyga, K., Martirosian, G., Szuba, A., Gamian, A., Myc, A., & Górska, S. (2019). Mapping Epitopes of a Novel Peptidoglycan Cross-Linking Enzyme Cwp22 Recognized by Human Sera Obtained from Patients with Clostridioides difficile Infection and Cord Blood. Microorganisms, 7(11), 565. https://doi.org/10.3390/microorganisms7110565