Candida glabrata: A Lot More Than Meets the Eye


Abstract
:1. Introduction
2. Genomic Architecture
3. Salient Pathobiological Features
3.1. Adherence
3.2. Biofilm Formation
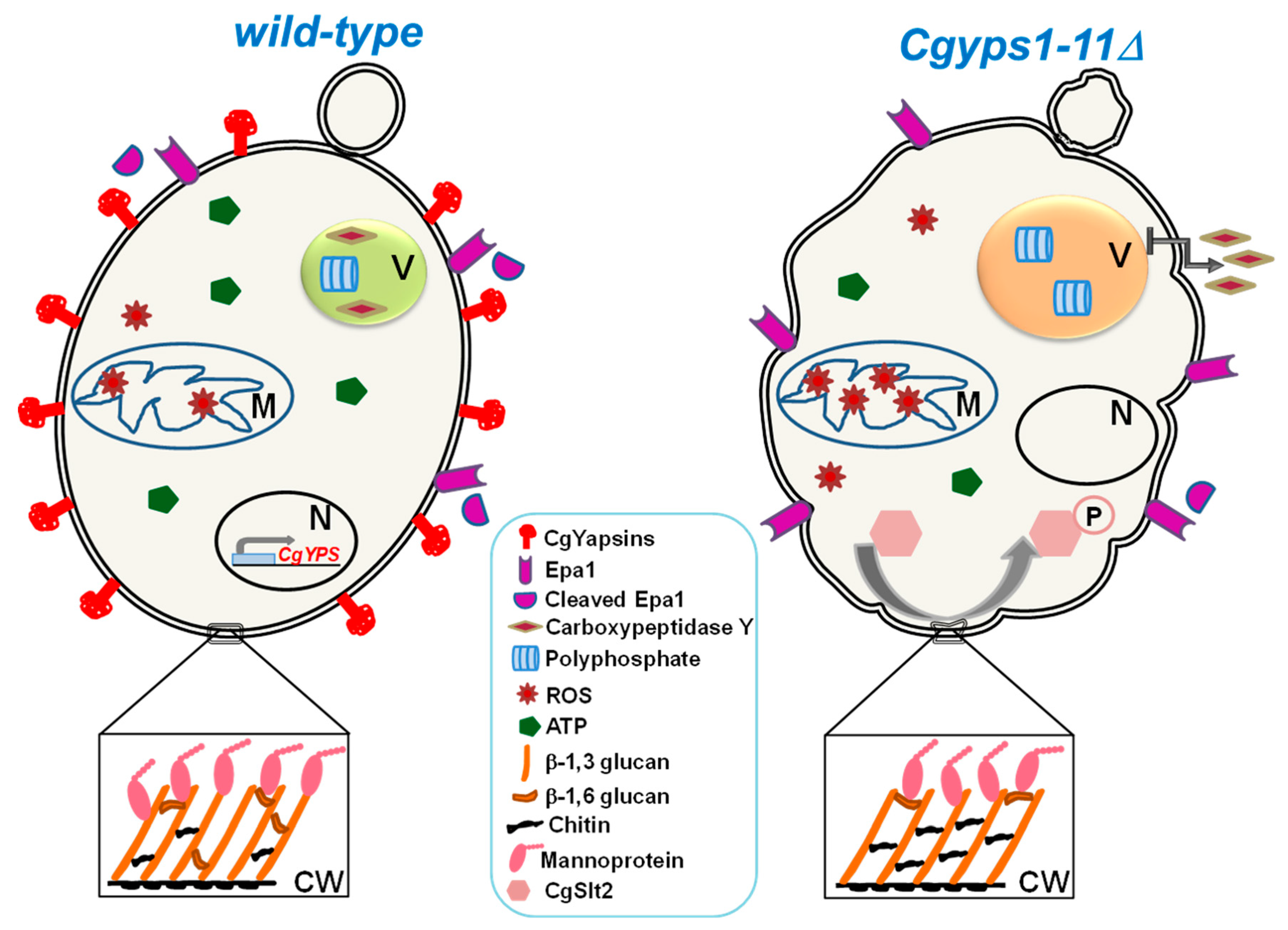
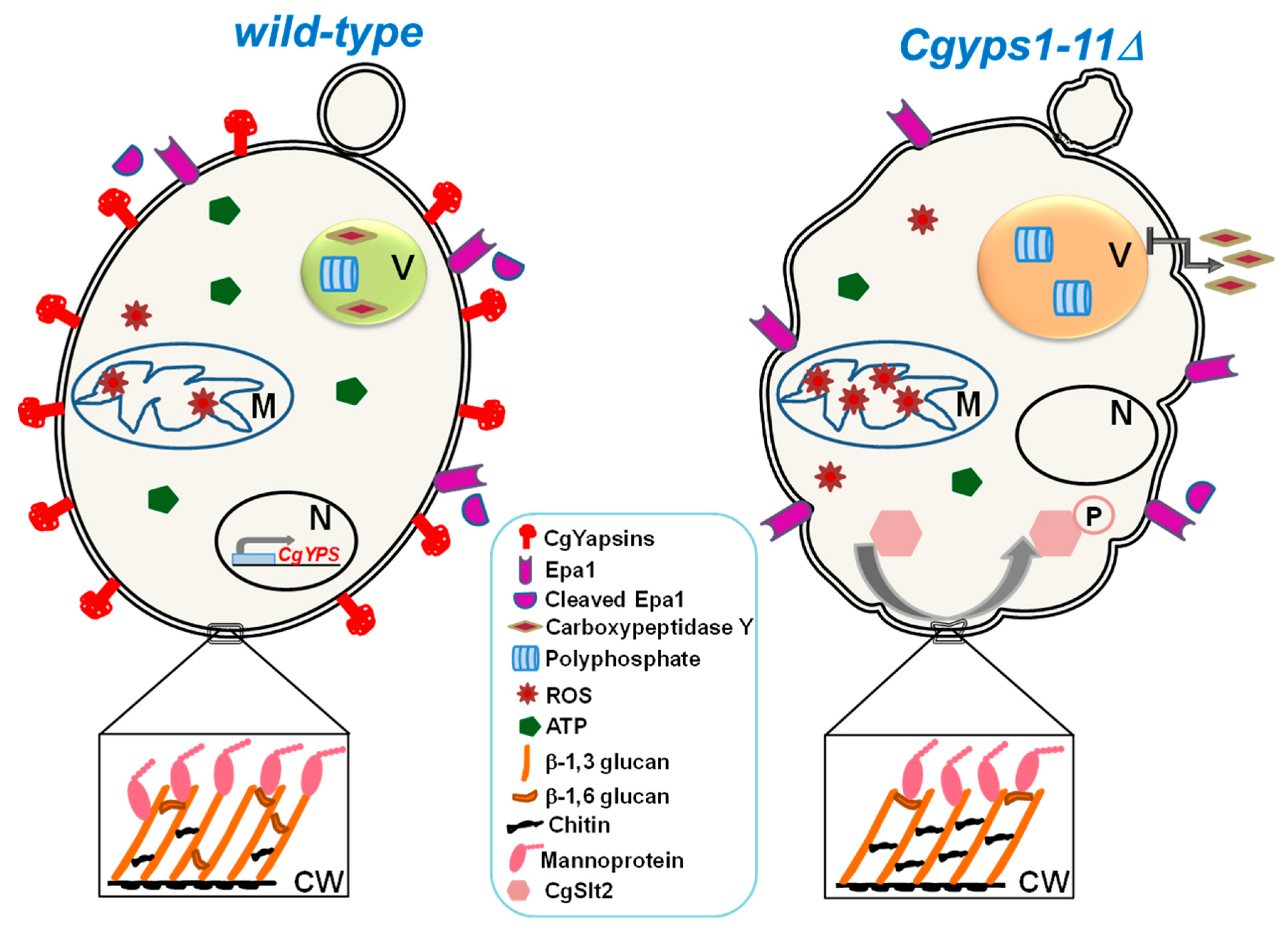
3.3. Aspartyl Proteases
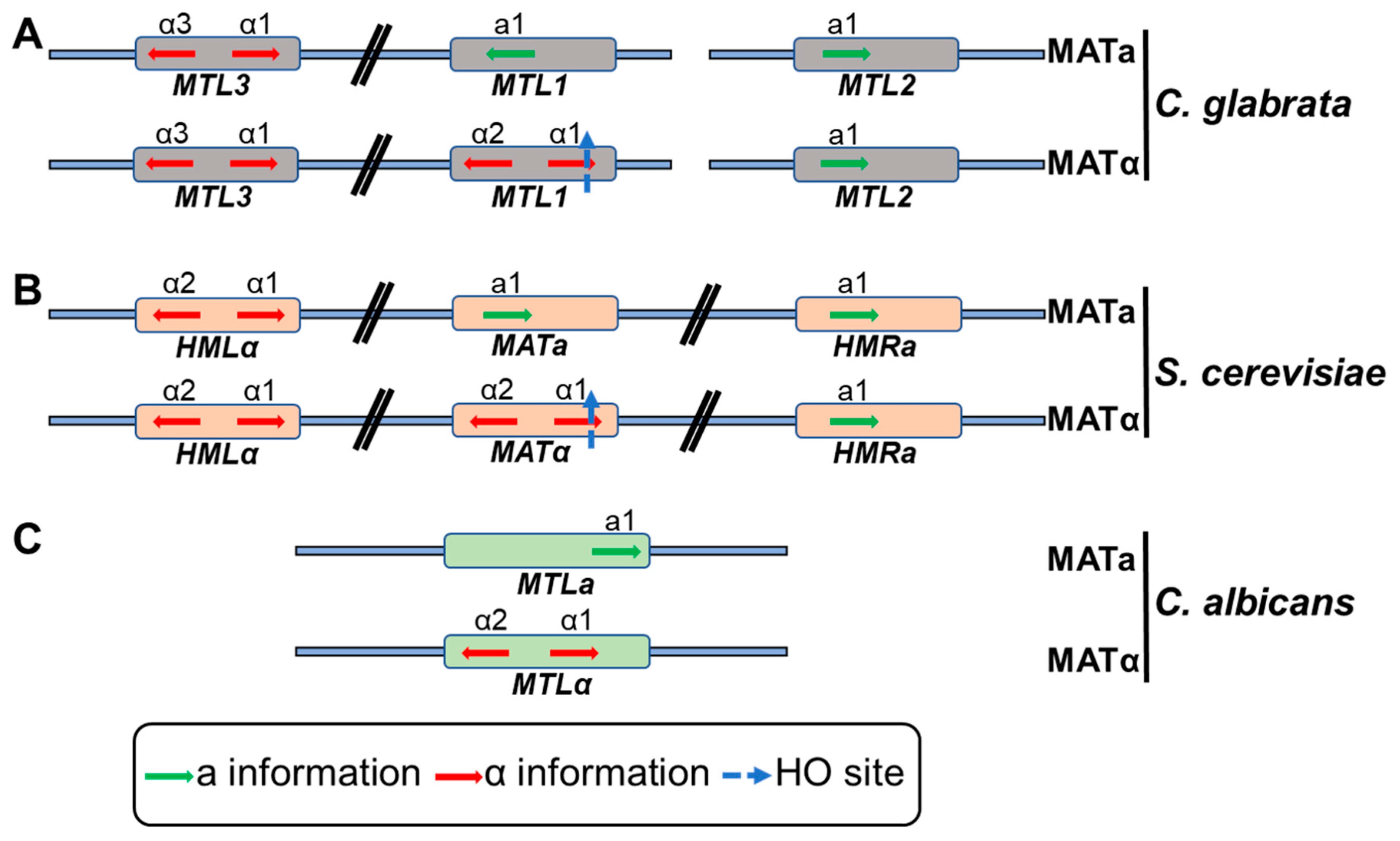
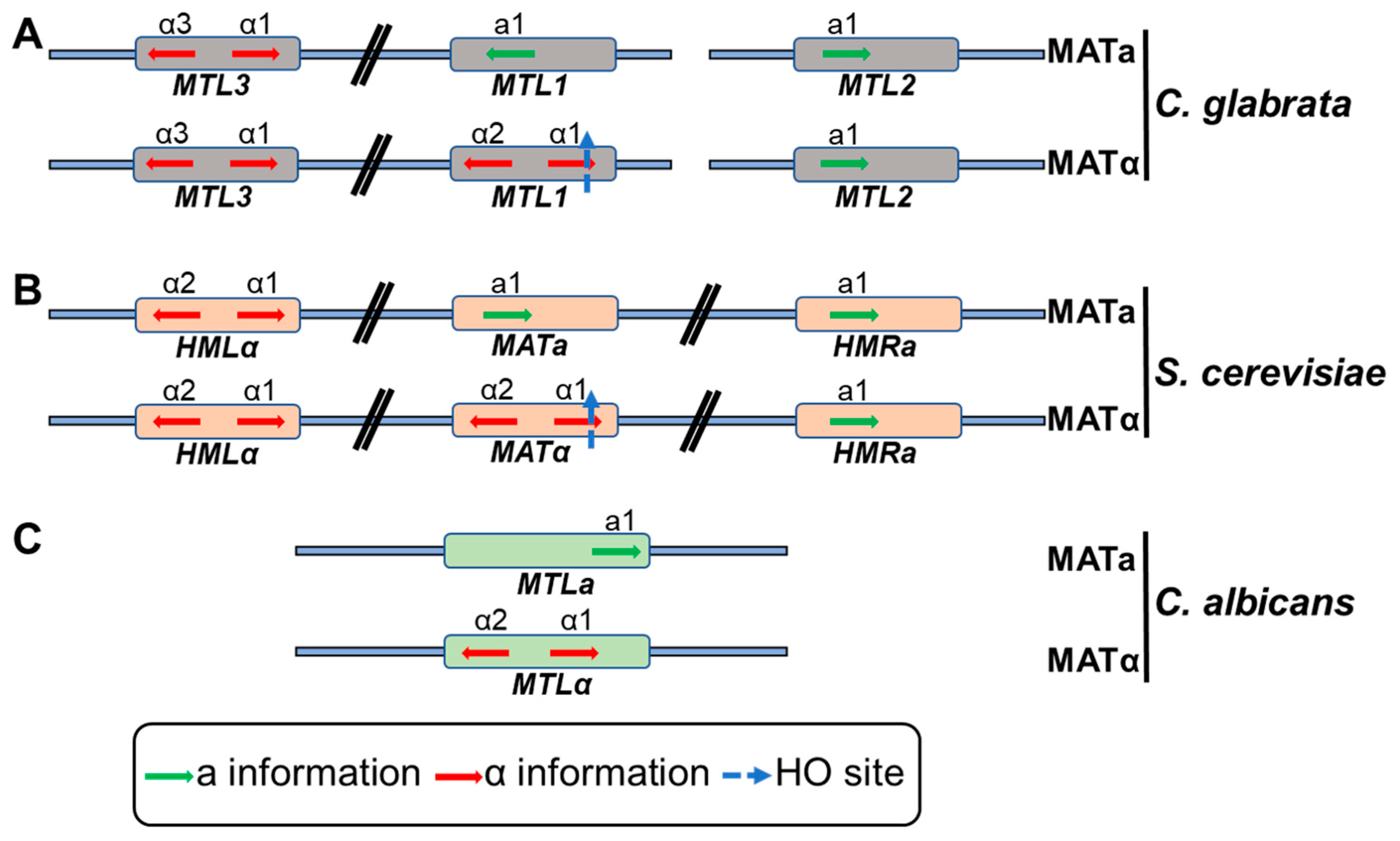
3.4. Colony and Mating-Type Switching
3.5. Stress Response Mechanisms
3.6. Nutrient Acquisition Pathways
3.6.1. Carbon Acquisition
3.6.2. Phosphate Acquisition
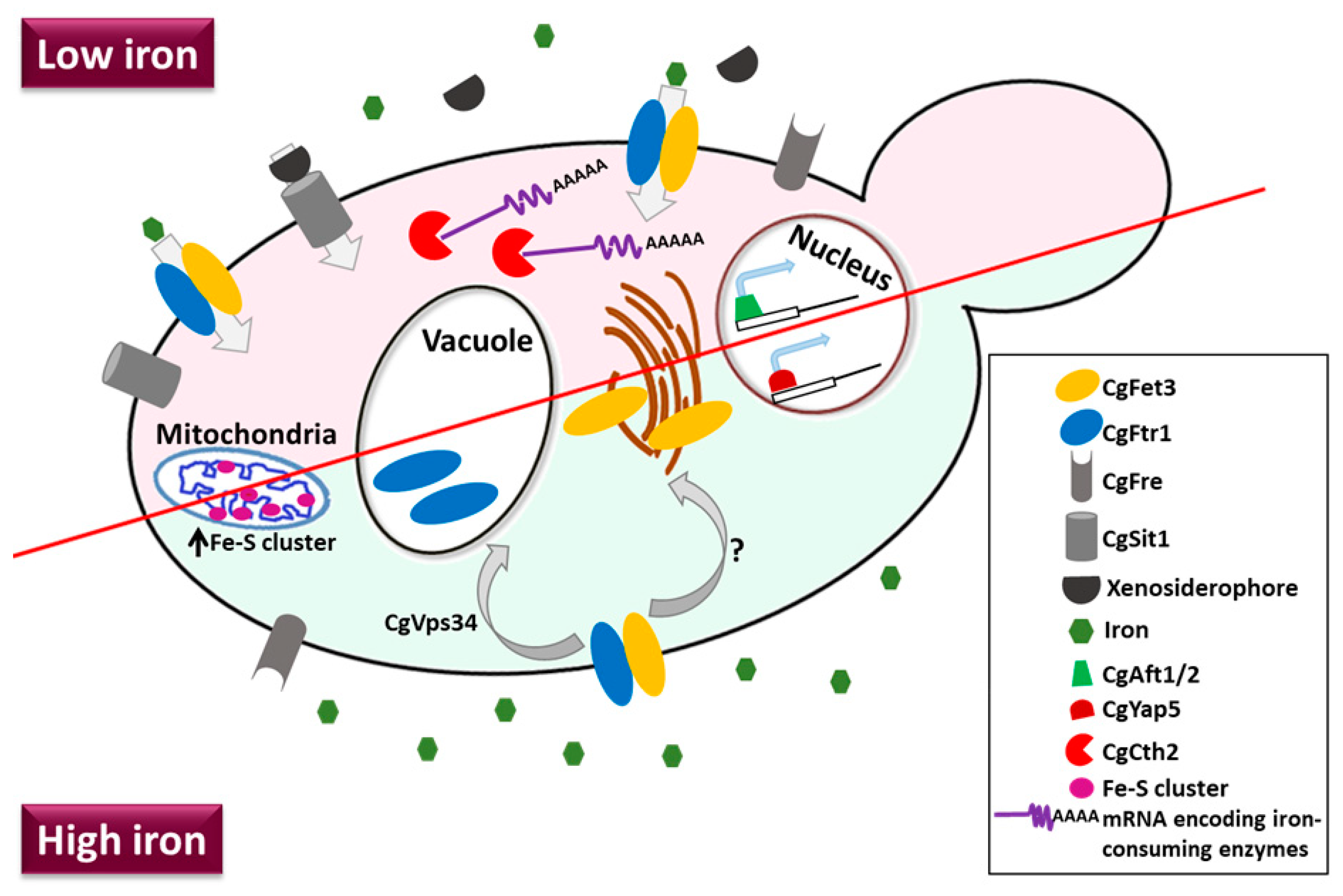
3.6.3. Iron Acquisition
4. C. glabrata-Host Interaction
4.1. C. glabrata-Epithelial Cell Interaction
4.2. C. glabrata-Endothelial Cell Interaction
4.3. C. glabrata-Neutrophil Interaction
4.4. C. glabrata-Natural Killer Cell Interaction
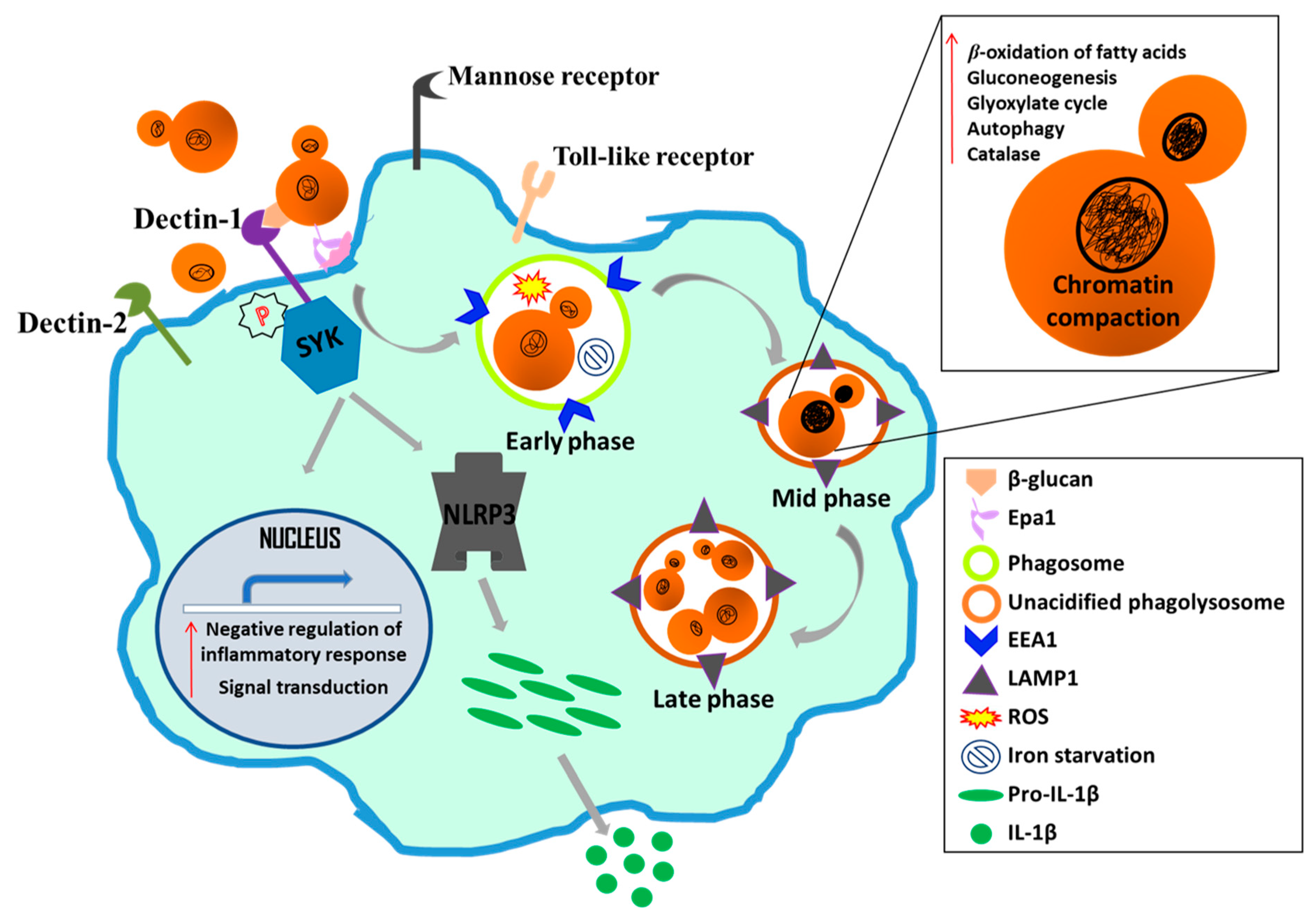
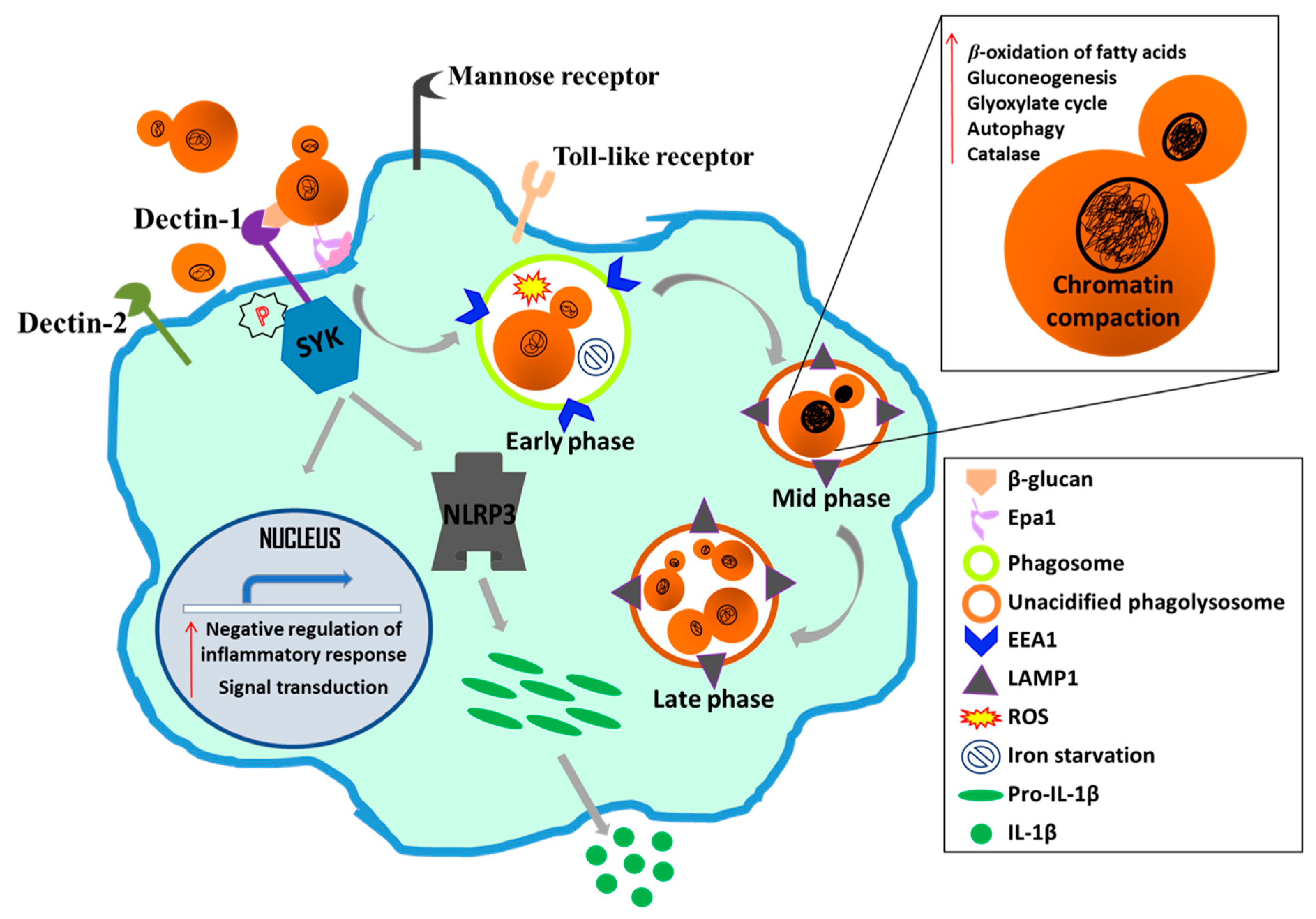
4.5. C. glabrata-Macrophage Interaction
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pfaller, M.A.; Messer, S.A.; Moet, G.J.; Jones, R.N.; Castanheira, M. Candida bloodstream infections: Comparison of species distribution and resistance to echinocandin and azole antifungal agents in Intensive Care Unit (ICU) and non-ICU settings in the SENTRY antimicrobial surveillance program (2008–2009). Int. J. Antimicrob. Agents 2011, 38, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Montagna, M.T.; Lovero, G.; Borghi, E.; Amato, G.; Andreoni, S.; Campion, L.; Lo Cascio, G.; Lombardi, G.; Luzzaro, F.; Manso, E.; et al. Candidemia in intensive care unit: A nationwide prospective observational survey (GISIA-3 study) and review of the European literature from 2000 through 2013. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 661–674. [Google Scholar] [PubMed]
- Yapar, N. Epidemiology and risk factors for invasive candidiasis. Ther. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Sood, P.; Rudramurthy, S.M.; Chen, S.; Kaur, H.; Capoor, M.; Chhina, D.; Rao, R.; Eshwara, V.K.; Xess, I.; et al. Incidence, characteristics and outcome of ICU-acquired candidemia in India. Intensive Care Med. 2014, 41, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.; Arbefeville, S.; Boyken, L.; Kroeger, J.; Pfaller, M. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn. Microbiol. Infect. Dis. 2012, 73, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.H.; Chakrabarti, A.; Li, R.Y.; Patel, A.K.; Watcharananan, S.P.; Liu, Z.; Chindamporn, A.; Tan, A.L.; Sun, P.L.; Wu, U.I.; et al. Incidence and species distribution of candidaemia in Asia: A laboratory-based surveillance study. Clin. Microbiol. Infect. 2015, 21, 946–953. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Andes, D.R.; Diekema, D.J.; Horn, D.L.; Reboli, A.C.; Rotstein, C.; Franks, B.; Azie, N.E. Epidemiology and outcomes of invasive candidiasis due to non-albicans species of Candida in 2,496 patients: Data from the Prospective Antifungal Therapy (PATH) registry 2004–2008. PLoS ONE 2014, 9, e101510. [Google Scholar] [CrossRef]
- Khatib, R.; Johnson, L.B.; Fakih, M.G.; Riederer, K.; Briski, L. Current trends in candidemia and species distribution among adults: Candida glabrata surpasses C. albicans in diabetic patients and abdominal sources. Mycoses 2016, 59, 781–786. [Google Scholar] [CrossRef]
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef]
- Achkar, J.M.; Fries, B.C. Candida infections of the genitourinary tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A. A history of research on yeasts 8: Taxonomy. Yeast 2004, 21, 1141–1193. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Phylogenetic circumscription of Saccharomyces, Kluyveromyces and other members of the Saccharomycetaceae, and the proposal of the new genera Lachancea, Nakaseomyces, Naumovia, Vanderwaltozyma and Zygotorulaspora. FEMS Yeast Res. 2003, 4, 233–245. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef]
- Zacharioudakis, I.; Zervou, F.; Mylonakis, E. T2 Magnetic Resonance Assay: Overview of available data and clinical implications. J. Fungi 2018, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Clancy, C.J.; Nguyen, M.H. Diagnosing invasive candidiasis. J. Clin. Microbiol. 2018, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef]
- Gabaldón, T.; Martin, T.; Marcet-Houben, M.; Durrens, P.; Bolotin-Fukuhara, M.; Lespinet, O.; Arnaise, S.; Boisnard, S.; Aguileta, G.; Atanasova, R.; et al. Comparative genomics of emerging pathogens in the Candida glabrata clade. BMC Genomics 2013, 14, 623. [Google Scholar] [CrossRef]
- Jong, H.S.; Myung, J.C.; Jeong, W.S.; Jung, S.I.; Cho, D.; Seung, J.K.; Soo, H.K.; Myung, G.S.; Soon, P.S.; Dong, W.R. Changes in karyotype and azole susceptibility of sequential bloodstream isolates from patients with Candida glabrata candidemia. J. Clin. Microbiol. 2007, 45, 2385–2391. [Google Scholar]
- Bader, O.; Schwarz, A.; Kraneveld, E.A.; Tangwattanchuleeporn, M.; Schmidt, P.; Jacobsen, M.D.; Gross, U.; de Groot, P.W.J.; Weig, M. Gross karyotypic and phenotypic alterations among different progenies of the Candida glabrata CBS138/ATCC2001 reference strain. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef]
- Ahmad, K.M.; Ishchuk, O.P.; Hellborg, L.; Jørgensen, G.; Skvarc, M.; Stenderup, J.; Jørck-Ramberg, D.; Polakova, S.; Piškur, J. Small chromosomes among Danish Candida glabrata isolates originated through different mechanisms. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2013, 104, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Rocha, E.P.C.; Brunet, F.; Vergassola, M.; Dujon, B. Highly variable rates of genome rearrangements between hemiascomycetous yeast lineages. PLoS Genet. 2006, 2, 0253–0261. [Google Scholar] [CrossRef] [PubMed]
- Desai, C.; Mavrianos, J.; Chauhan, N. Candida glabrata Pwp7p and Aed1p are required for adherence to human endothelial cells. FEMS Yeast Res. 2011, 11, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Marcet-Houben, M.; Gabaldón, T. Beyond the whole-genome duplication: Phylogenetic evidence for an ancient interspecies hybridization in the baker’s yeast lineage. PLoS Biol. 2015, 13, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Carreté, L.; Ksiezopolska, E.; Pegueroles, C.; Gómez-Molero, E.; Saus, E.; Iraola-Guzmán, S.; Loska, D.; Bader, O.; Fairhead, C.; Gabaldón, T. Patterns of genomic variation in the opportunistic pathogen Candida glabrata suggest the existence of mating and a secondary association with humans. Curr. Biol. 2018, 28, 15–27.e7. [Google Scholar] [CrossRef] [PubMed]
- Koszul, R.; Malpertuy, A.; Frangeul, L.; Bouchier, C.; Wincker, P.; Thierry, A.; Duthoy, S.; Ferris, S.; Hennequin, C.; Dujon, B. The complete mitochondrial genome sequence of the pathogenic yeast Candida (Torulopsis) glabrata. FEBS Lett. 2003, 534, 39–48. [Google Scholar] [CrossRef]
- De Groot, P.W.J.; Kraneveld, E.A.; Qing, Y.Y.; Dekker, H.L.; Groß, U.; Crielaard, W.; De Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell 2008, 7, 1951–1964. [Google Scholar] [CrossRef]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A yeast by any other name: Candida glabrata and its interaction with the host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef]
- De Las Peñas, A.; Pan, S.J.; Castaño, I.; Alder, J.; Cregg, R.; Cormack, B.P. Virulence-related surface glycoproteins in the yeast pathogen Candida glabrata are encoded in subtelomeric clusters and subject to RAP1- and SIR-dependent transcriptional silencing. Genes Dev. 2003, 17, 2245–2258. [Google Scholar] [CrossRef]
- Domergue, R. Nicotinic acid limitation regulates silencing of Candida adhesins during UTI. Science 2005, 308, 866–870. [Google Scholar] [CrossRef]
- López-Fuentes, E.; Gutiérrez-Escobedo, G.; Timmermans, B.; Van Dijck, P.; De Las Peñas, A.; Castaño, I. Candida glabrata’s genome plasticity confers a unique pattern of expressed cell wall proteins. J. Fungi 2018, 4, 67. [Google Scholar] [CrossRef]
- Cormack, B.P. An adhesin of the yeast pathogen Candida glabrata mediating adherence to human epithelial cells. Science 1999, 285, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.M.; Vyas, V.K. The Candida glabrata adhesin Epa1p causes adhesion, phagocytosis, and cytokine secretion by innate immune cells. FEMS Yeast Res. 2012, 12, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Vale-Silva, L.A.; Moeckli, B.; Torelli, R.; Posteraro, B.; Sanglard, D. Upregulation of the adhesin gene EPA1 mediated by PDR1 in Candida glabrata leads to enhanced host colonization. mSphere 2016, 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, M.L.; Frieman, M.; Smith, D.; Alvarez, R.A.; Cummings, R.D.; Cormack, B.P. Glycan microarray analysis of Candida glabrata adhesin ligand specificity. Mol. Microbiol. 2008, 68, 547–559. [Google Scholar] [CrossRef]
- Castaño, I.; Pan, S.J.; Zupancic, M.; Hennequin, C.; Dujon, B.; Cormack, B.P. Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata. Mol. Microbiol. 2005, 55, 1246–1258. [Google Scholar] [CrossRef]
- Iraqui, I.; Garcia-Sanchez, S.; Aubert, S.; Dromer, F.; Ghigo, J.M.; D’Enfert, C.; Janbon, G. The Yak1p kinase controls expression of adhesins and biofilm formation in Candida glabrata in a Sir4p-dependent pathway. Mol. Microbiol. 2005, 55, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schüller, C. Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef]
- Mundy, R.D.; Cormack, B. Expression of Candida glabrata adhesins after exposure to chemical preservatives. J. Infect. Dis. 2009, 199, 1891–1898. [Google Scholar] [CrossRef]
- Kraneveld, E.A.; de Soet, J.J.; Deng, D.M.; Dekker, H.L.; de Koster, C.G.; Klis, F.M.; Crielaard, W.; de Groot, P.W.J. Identification and differential gene expression of adhesin-like wall proteins in Candida glabrata biofilms. Mycopathologia. 2011, 172, 415–427. [Google Scholar] [CrossRef]
- Gómez-Molero, E.; de Boer, A.D.; Dekker, H.L.; Moreno-Martínez, A.; Kraneveld, E.A.; Ichsan; Chauhan, N.; Weig, M.; de Soet, J.J.; de Koster, C.G.; et al. Proteomic analysis of hyperadhesive Candida glabrata clinical isolates reveals a core wall proteome and differential incorporation of adhesins. FEMS Yeast Res. 2015, 15, fov098. [Google Scholar]
- Juárez-Cepeda, J.; Orta-Zavalza, E.; Cañas-Villamar, I.; Arreola-Gómez, J.; Pérez-Cornejo, G.P.; Hernández-Carballo, C.Y.; Gutiérrez-Escobedo, G.; Castaño, I.; De Las Peñas, A. The EPA2 adhesin encoding gene is responsive to oxidative stress in the opportunistic fungal pathogen Candida glabrata. Curr. Genet. 2015, 61, 529–544. [Google Scholar] [CrossRef]
- Maestre-reyna, M.; Diderrich, R.; Stefan, M.; Eulenburg, G.; Kalugin, V.; Brückner, S. Structural basis for promiscuity and specificity during Candida glabrata invasion of host epithelia. Proc. Natl. Acad. Sci. USA 2012, 109, 16864–16869. [Google Scholar] [CrossRef]
- Ielasi, F.S.; Alioscha-perez, M.; Donohue, D.; Claes, S.; Sahli, H.; Schols, D. Lectin-glycan interaction network-based identification of host receptors of microbial pathogenic adhesins. mBio 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Diderrich, R.; Kock, M.; Maestre-Reyna, M.; Keller, P.; Steuber, H.; Rupp, S.; Essen, L.O.; Mösch, H.U. Structural hot spots determine functional diversity of the Candida glabrata epithelial adhesin family. J. Biol. Chem. 2015, 290, 19597–19613. [Google Scholar] [CrossRef]
- Linde, J.; Duggan, S.; Weber, M.; Horn, F.; Sieber, P.; Hellwig, D.; Riege, K.; Marz, M.; Martin, R.; Guthke, R.; et al. Defining the transcriptomic landscape of Candida glabrata by RNA-Seq. Nucleic Acids Res. 2015, 43, 1392–1406. [Google Scholar] [CrossRef]
- Thierry, A.; Bouchier, C.; Dujon, B.; Richard, G.F. Megasatellites: A peculiar class of giant minisatellites in genes involved in cell adhesion and pathogenicity in Candida glabrata. Nucleic Acids Res. 2008, 36, 5970–5982. [Google Scholar] [CrossRef]
- Vale-Silva, L.; Beaudoing, E.; Tran, V.D.T.; Sanglard, D. Comparative genomics of two sequential Candida glabrata clinical isolates. G3 2017, 7, 2413–2426. [Google Scholar] [CrossRef]
- Salazar, S.B.; Wang, C.; Musterkotter, M.; Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; Lopes, M.M.; Güldener, U.; Butler, G.; Mira, N.P. Comparative genomic and transcriptomic analyses unveil novel features of azole resistance and adaptation to the human host in Candida glabrata. FEMS Yeast Res. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- D’Enfert, C.; Janbon, G. Biofilm formation in Candida glabrata: What have we learnt from functional genomics approaches? FEMS Yeast Res. 2015, 16, 1–13. [Google Scholar]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Taff, H.T.; Cuevas, M.A.; Reinicke, E.L.; Sanchez, H.; Andes, D.R. Role of matrix β-1,3 glucan in antifungal resistance of non-albicans Candida biofilms. Antimicrob. Agents Chemother. 2013, 57, 1918–1920. [Google Scholar] [CrossRef]
- Riera, M.; Mogensen, E.; d’Enfert, C.; Janbon, G. New regulators of biofilm development in Candida glabrata. Res. Microbiol. 2012, 163, 297–307. [Google Scholar] [CrossRef]
- Schwarzmuller, T.; Ma, B.; Hiller, E.; Istel, F.; Tscherner, M.; Brunke, S.; Ames, L.; Firon, A.; Green, B.; Cabral, V.; et al. Systematic phenotyping of a large-scale Candida glabrata deletion collection reveals novel antifungal tolerance genes. PLoS Pathog. 2014, 10, e1004211. [Google Scholar] [CrossRef]
- Kucharíková, S.; Neirinck, B.; Sharma, N.; Vleugels, J.; Lagrou, K.; Van Dijck, P. In vivo Candida glabrata biofilm development on foreign bodies in a rat subcutaneous model. J. Antimicrob. Chemother. 2015, 70, 846–856. [Google Scholar] [CrossRef]
- Jayampath Seneviratne, C.; Wang, Y.; Jin, L.; Abiko, Y.; Samaranayake, L.P. Proteomics of drug resistance in Candida glabrata biofilms. Proteomics 2010, 10, 1444–1454. [Google Scholar] [CrossRef]
- Sharma, V.; Purushotham, R.; Kaur, R. The phosphoinositide 3-kinase regulates retrograde trafficking of the iron permease CgFtr1 and iron homeostasis in Candida glabrata. J. Biol. Chem. 2016, 291, 24715–24734. [Google Scholar] [CrossRef]
- Rasheed, M.; Battu, A.; Kaur, R. Aspartyl proteases in Candida glabrata are required for suppression of the host innate immune response. J. Biol. Chem. 2018, 293, 6410–6433. [Google Scholar] [CrossRef]
- Kantarcioǧlu, A.S.; Yücel, A. Phospholipase and protease activities in clinical Candida isolates with reference to the sources of strains. Mycoses 2002, 45, 160–165. [Google Scholar] [CrossRef]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef]
- Bairwa, G.; Kaur, R. A novel role for a glycosylphosphatidylinositol-anchored aspartyl protease, CgYps1, in the regulation of pH homeostasis in Candida glabrata. Mol. Microbiol. 2011, 79, 900–913. [Google Scholar] [CrossRef]
- Miyazaki, T.; Izumikawa, K.; Yamauchi, S.; Inamine, T.; Nagayoshi, Y.; Saijo, T.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. The glycosylphosphatidylinositol-linked aspartyl protease Yps1 is transcriptionally regulated by the calcineurin-Crz1 and Slt2 MAPK pathways in Candida glabrata. FEMS Yeast Res. 2011, 11, 449–456. [Google Scholar] [CrossRef]
- Fukuda, Y.; Tsai, H.F.; Myers, T.G.; Bennett, J.E. Transcriptional profiling of Candida glabrata during phagocytosis by neutrophils and in the infected mouse spleen. Infect. Immun. 2013, 81, 1325–1333. [Google Scholar] [CrossRef]
- Bairwa, G.; Rasheed, M.; Taigwal, R.; Sahoo, R.; Kaur, R. GPI (glycosylphosphatidylinositol)-linked aspartyl proteases regulate vacuole homoeostasis in Candida glabrata. Biochem. J. 2014, 458, 323–334. [Google Scholar] [CrossRef]
- Seider, K.; Brunke, S.; Schild, L.; Jablonowski, N.; Wilson, D.; Majer, O.; Barz, D.; Haas, A.; Kuchler, K.; Schaller, M.; et al. The facultative intracellular pathogen Candida glabrata subverts macrophage cytokine production and phagolysosome maturation. J. Immunol. 2011, 187, 3072–3086. [Google Scholar] [CrossRef]
- Quintin, J.; Asmar, J.; Matskevich, A.A.; Lafarge, M.-C.; Ferrandon, D. The Drosophila toll pathway controls but does not clear Candida glabrata infections. J. Immunol. 2013, 190, 2818–2827. [Google Scholar] [CrossRef]
- Enkler, L.; Richer, D.; Marchand, A.L.; Ferrandon, D.; Jossinet, F. Genome engineering in the yeast pathogen Candida glabrata using the CRISPR-Cas9 system. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Lachke, S.A.; Joly, S.; Daniels, K.; Soll, D.R. Phenotypic switching and filamentation in Candida glabrata. Microbiology 2002, 148, 2661–2674. [Google Scholar] [CrossRef]
- Brockert, P.J.; Lachke, S.A.; Srikantha, T.; Pujol, C.; Galask, R.; Soll, D.R. Phenotypic switching and mating type switching of Candida glabrata at sites of colonization. Infect. Immun. 2003, 71, 7109–7118. [Google Scholar] [CrossRef]
- Csank, C.; Haynes, K. Candida glabrata displays pseudohyphal growth. FEMS Microbiol. Lett. 2000, 189, 115–120. [Google Scholar] [CrossRef]
- Sasani, E.; Khodavaisy, S.; Agha Kuchak Afshari, S.; Darabian, S.; Aala, F.; Rezaie, S. Pseudohyphae formation in Candida glabrata due to CO2 exposure. Curr. Med. Mycol. 2016, 2, 49–52. [Google Scholar] [CrossRef]
- Kamran, M.; Calcagno, A.; Findon, H.; Bignell, E.; Jones, M.D.; Warn, P.; Denning, D.W.; Butler, G.; Mühlschlegel, F.A.; Haynes, K.; et al. Inactivation of transcription factor gene ACE2 in the fungal pathogen Candida glabrata results in hypervirulence. Eukaryot. Cell 2004, 3, 546–552. [Google Scholar] [CrossRef]
- MacCallum, D.M.; Findon, H.; Kenny, C.C.; Butler, G.; Haynes, K.; Odds, F.C. Different consequences of ACE2 and SWI5 gene disruptions for virulence of pathogenic and nonpathogenic yeasts. Infect. Immun. 2006, 74, 5244–5248. [Google Scholar] [CrossRef]
- Srikantha, T.; Lachke, S.A.; Soll, D.R. Three mating type-like loci in Candida glabrata. Eukaryot. Cell 2003, 2, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.A.; Heitman, J. Fungal mating-type loci. Curr. Biol. 2003, 13, R792–R795. [Google Scholar] [CrossRef]
- Robledo-Márquez, K.; Gutiérrez-Escobedo, G.; Yáñez-Carrillo, P.; Vidal-Aguiar, Y.; Briones-Martín-del-Campo, M.; Orta-Zavalza, E.; De Las Peñas, A.; Castaño, I. Candida glabrata encodes a longer variant of the mating type (MAT) alpha2 gene in the mating type-like MTL3 locus, which can form homodimers. FEMS Yeast Res. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Ramírez-Zavaleta, C.Y.; Salas-Delgado, G.E.; de Las Peñas, A.; Castaño, I. Subtelomeric silencing of the MTL3 locus of Candida glabrata requires yKu70, yKu80, and Rif1 proteins. Eukaryot. Cell 2010, 9, 1602–1611. [Google Scholar] [CrossRef]
- Boisnard, S.; Li, Y.Z.; Arnaise, S.; Sequeira, G.; Raffoux, X.; Enache-Angoulvant, A.; Bolotin-Fukuhara, M.; Fairhead, C. Efficient mating-type switching in Candida glabrata induces cell death. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef]
- Nikolaou, E.; Agrafioti, I.; Stumpf, M.; Quinn, J.; Stansfield, I.; Brown, A.J. Phylogenetic diversity of stress signalling pathways in fungi. BMC Evol. Biol. 2009, 9, 1–18. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Briones-Martin-del-Campo, M.; Cañas-Villamar, I.; Montalvo-Arredondo, J.; Riego-Ruiz, L.; Castaño, I.; De Las Peñas, A. High resistance to oxidative stress in the fungal pathogen Candida glabrata is mediated by a single catalase, Cta1p, and is controlled by the transcription factors Yap1p, Skn7p, Msn2p, and Msn4p. Eukaryot. Cell 2008, 7, 814–825. [Google Scholar] [CrossRef]
- Briones-Martin-Del-Campo, M.; Orta-Zavalza, E.; Caïas-Villamar, I.; Gutiïrrez-Escobedo, G.; Juïrez-Cepeda, J.; Robledo-Mïrquez, K.; Arroyo-Helguera, O.; Castaïo, I.; De Las Peïas, A. The superoxide dismutases of Candida glabrata protect against oxidative damage and are required for lysine biosynthesis, DNA integrity and chronological life survival. Microbiology 2015, 161, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Desai, P.R.; Rai, M.N.; Kaur, R.; Ganesan, K.; Bachhawat, A.K. Glutathione biosynthesis in the yeast pathogens Candida glabrata and Candida albicans: Essential in C. glabrata, and essential for virulence in C. albicans. Microbiology 2011, 157, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Escobedo, G.; Orta-Zavalza, E.; Castaño, I.; De Las Peñas, A. Role of glutathione in the oxidative stress response in the fungal pathogen Candida glabrata. Curr. Genet. 2013, 59, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Gratz, N.; Kovarik, P.; Schüller, C. Autophagy supports Candida glabrata survival during phagocytosis. Cell. Microbiol. 2010, 12, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Klopf, E.; Gratz, N.; Marcet-Houben, M.; Hiller, E.; Rupp, S.; Gabaldón, T.; Kovarik, P.; Schüller, C. Regulation of Candida glabrata oxidative stress resistance is adapted to host environment. FEBS Lett. 2011, 585, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Saijo, T.; Miyazaki, T.; Izumikawa, K.; Mihara, T.; Takazono, T.; Kosai, K.; Imamura, Y.; Seki, M.; Kakeya, H.; Yamamoto, Y.; et al. Skn7p is involved in oxidative stress response and virulence of Candida glabrata. Mycopathologia 2010, 169, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Orta-Zavalza, E.; Guerrero-Serrano, G.; Gutierrez-Escobedo, G.; Canas-Villamar, I.; Juarez-Cepeda, J.; Castano, I.; De Las Penas, A. Local silencing controls the oxidative stress response and the multidrug resistance in Candida glabrata. Mol. Microbiol. 2013, 88, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Kounatidis, I.; Ames, L.; Mistry, R.; Ho, H.-L.; Haynes, K.; Ligoxygakis, P. A host-pathogen interaction screen identifies ada2 as a mediator of Candida glabrata defences against reactive oxygen species. G3 2018, 8, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Brunke, S.; Seider, K.; Almeida, R.S.; Heyken, A.; Fleck, C.B.; Brock, M.; Barz, D.; Rupp, S.; Hube, B. Candida glabrata tryptophan-based pigment production via the Ehrlich pathway. Mol. Microbiol. 2010, 76, 25–47. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Inamine, T.; Yamauchi, S.; Nagayoshi, Y.; Saijo, T.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. Role of the Slt2 mitogen-activated protein kinase pathway in cell wall integrity and virulence in Candida glabrata. FEMS Yeast Res. 2010, 10, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Shivarathri, R.; Kaur, R. The Rho1 GTPase-activating protein CgBem2 is required for survival of azole stress in Candida glabrata. J. Biol. Chem. 2011, 286, 34311–34324. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, M.; Ito, F.; Aoyama, T.; Sato-Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; Shibata, N. Cooperation between ER stress and calcineurin signaling contributes to the maintenance of cell wall integrity in Candida glabrata. Fungal Biol. 2018, 122, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Yamauchi, S.; Inamine, T.; Nagayoshi, Y.; Saijo, T.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. Roles of calcineurin and Crz1 in antifungal susceptibility and virulence of Candida glabrata. Antimicrob. Agents Chemother. 2010, 54, 1639–1643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Konieczka, J.H.; Springer, D.J.; Bowen, S.E.; Zhang, J.; Silao, F.G.S.; Bungay, A.A.C.; Bigol, U.G.; Nicolas, M.G.; Abraham, S.N.; et al. Convergent evolution of calcineurin pathway roles in thermotolerance and virulence in Candida glabrata. G3 2012, 2, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Jandric, Z.; Gregori, C.; Klopf, E.; Radolf, M.; Schüller, C. Sorbic acid stress activates the Candida glabrata high osmolarity glycerol MAP kinase pathway. Front. Microbiol. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.K.; Suneetha, K.J.; Kaur, R. The mitogen-activated protein kinase CgHog1 is required for iron homeostasis, adherence and virulence in Candida glabrata. FEBS J. 2015, 282, 2142–2166. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Nakayama, H.; Nagayoshi, Y.; Kakeya, H.; Kohno, S. Dissection of Ire1 functions reveals stress response mechanisms uniquely evolved in Candida glabrata. PLoS Pathog. 2013, 9, e1003160. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-J.; Chang, Y.-L.; Chen, Y.-L. Deletion of ADA2 increases antifungal drug susceptibility and virulence in Candida glabrata. Antimicrob. Agents Chemother. 2018, 62, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Castano, I.; Cormack, B.P. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida glabrata: Roles of calcium signaling and mitochondria. Antimicrob. Agents Chemother. 2004, 48, 1600–1613. [Google Scholar] [CrossRef] [PubMed]
- Singh-Babak, S.D.; Babak, T.; Diezmann, S.; Hill, J.A.; Xie, J.L.; Chen, Y.L.; Poutanen, S.M.; Rennie, R.P.; Heitman, J.; Cowen, L.E. Global analysis of the evolution and mechanism of echinocandin resistance in Candida glabrata. PLoS Pathog. 2012, 8, e1002718. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Rogers, P.D. Azole Resistance in Candida glabrata. Curr. Infect. Dis. Rep. 2016, 18, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Messer, S.A.; Rhomberg, P.R.; Pfaller, M.A. Antifungal susceptibility patterns of a global collection of fungal isolates: Results of the SENTRY antifungal surveillance program (2013). Diagn. Microbiol. Infect. Dis. 2016, 85, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.; Perlin, D.S. Fungal resistance to echinocandins and the MDR phenomenon in Candida glabrata. J. Fungi 2018, 4, 105. [Google Scholar]
- Sanglard, D.; Ischer, F.; Calabrese, D.; Majcherczyk, P.A.; Bille, J. The ATP binding cassette transporter gene CgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 1999, 43, 2753–2765. [Google Scholar] [CrossRef]
- Izumikawa, K.; Kakeya, H.; Tsai, H.F.; Grimberg, B.; Bennett, J.E. Function of Candida glabrata ABC transporter gene, PDH1. Yeast 2003, 20, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Cota, J.M.; Grabinski, J.L.; Talbert, R.L.; Burgess, D.S.; Rogers, P.D.; Edlind, T.D.; Wiederhold, N.P. Increases in SLT2 expression and chitin content are associated with incomplete killing of Candida glabrata by caspofungin. Antimicrob. Agents Chemother. 2008, 52, 1144–1146. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Tronchin, G.; Bergès, T.; Hennequin, C.; Chabasse, D.; Bouchara, J.P. Reduced susceptibility to polyenes associated with a missense mutation in the ERG6 gene in a clinical isolate of Candida glabrata with pseudohyphal growth. Antimicrob. Agents Chemother. 2007, 51, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Tronchin, G.; Larcher, G.; Ernoult, E.; Bergès, T.; Chabasse, D.; Bouchara, J.P. A nonsense mutation in the ERG6 gene leads to reduced susceptibility to polyenes in a clinical isolate of Candida glabrata. Antimicrob. Agents Chemother. 2008, 52, 3701–3709. [Google Scholar] [CrossRef]
- Healey, K.R.; Zhao, Y.; Perez, W.B.; Lockhart, S.R.; Sobel, J.D.; Farmakiotis, D.; Kontoyiannis, D.P.; Sanglard, D.; Taj-Aldeen, S.J.; Alexander, B.D.; et al. Prevalent mutator genotype identified in fungal pathogen Candida glabrata promotes multi-drug resistance. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Van Urk, H.; Voll, W.S.L.; Scheffers, W.A.; Van Dijken, J.P. Transient-state analysis of metabolic fluxes in crabtree-positive and crabtree-negative yeasts. Appl. Environ. Microbiol. 1990, 56, 281–287. [Google Scholar]
- Legrand, J.; Bolotin-Fukuhara, M.; Bourgais, A.; Fairhead, C.; Sicard, D. Life-history strategies and carbon metabolism gene dosage in the Nakaseomyces yeasts. FEMS Yeast Res. 2016, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Freese, S.; Vogts, T.; Speer, F.; Schäfer, B.; Passoth, V.; Klinner, U. C- and N-catabolic utilization of tricarboxylic acid cycle-related amino acids by Scheffersomyces stipitis and other yeasts. Yeast 2011, 28, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.S.; Chew, S.Y.; Rangasamy, P.; Mohd Desa, M.N.; Sandai, D.; Chong, P.P.; Than, L.T.L. SNF3 as high affinity glucose sensor and its function in supporting the viability of Candida glabrata under glucose-limited environment. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Conant, G.C.; Wolfe, K.H. Increased glycolytic flux as an outcome of whole-genome duplication in yeast. Mol. Syst. Biol. 2007, 3, 129. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, C.L.; Wykoff, D.D. Candida glabrata PHO4 is necessary and sufficient for Pho2-independent transcription of phosphate starvation genes. Genetics 2009, 182, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Orkwis, B.R.; Davies, D.L.; Kerwin, C.L.; Sanglard, D.; Wykoff, D.D. Novel acid phosphatase in Candida glabrata suggests selective pressure and niche specialization in the phosphate signal transduction pathway. Genetics 2010, 186, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, C.L.; Wykoff, D.D. De novo generation of a phosphate starvation-regulated promoter in Candida glabrata. FEMS Yeast Res. 2012, 12, 980–989. [Google Scholar] [CrossRef]
- He, B.Z.; Zhou, X.; O’Shea, E.K. Evolution of reduced co-activator dependence led to target expansion of a starvation response pathway. eLife 2017, 6, 1–26. [Google Scholar] [CrossRef]
- Nahas, J.V.; Iosue, C.L.; Shaik, N.F.; Selhorst, K.; He, B.Z.; Wykoff, D.D. Dynamic changes in yeast phosphatase families allow for specialization in phosphate and thiamine starvation. G3 2018, 8, 2333–2343. [Google Scholar] [CrossRef]
- Kornitzer, D. Fungal mechanisms for host iron acquisition. Curr. Opin. Microbiol. 2009, 12, 377–383. [Google Scholar] [CrossRef]
- Weissman, Z.; Kornitzer, D. A family of Candida cell surface haem-binding proteins involved in haemin and haemoglobin-iron utilization. Mol. Microbiol. 2004, 53, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.K.; Suneetha, K.J.; Kaur, R. A systematic analysis reveals an essential role for high-affinity iron uptake system, haemolysin and CFEM domain-containing protein in iron homoeostasis and virulence in Candida glabrata. Biochem. J. 2014, 463, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, T.; Thiele, D.J. Host iron withholding demands siderophore utilization for Candida glabrata to survive macrophage killing. PLoS Pathog. 2011, 7, e1001322. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Safyan, A.; Wisgott, S.; Brunke, S.; Kasper, L.; Hube, B. The fungal pathogen Candida glabrata does not depend on surface ferric reductases for iron acquisition. Front. Microbiol. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Safyan, A.; Wisgott, S.; Hille, F.; Kaemmer, P.; Linde, J.; Brunke, S.; Kasper, L.; Hube, B. A novel hybrid iron regulation network combines features from pathogenic and nonpathogenic yeasts. MBio 2016, 7, e01782-16. [Google Scholar] [CrossRef] [PubMed]
- Merhej, J.; Thiebaut, A.; Blugeon, C.; Pouch, J.; Ali Chaouche, M.E.A.; Camadro, J.M.; Le Crom, S.; Lelandais, G.; Devaux, F. A network of paralogous stress response transcription factors in the human pathogen Candida glabrata. Front. Microbiol. 2016, 7, 645. [Google Scholar] [CrossRef]
- Thiébaut, A.; Delaveau, T.; Benchouaia, M.; Boeri, J.; Garcia, M.; Lelandais, G.; Devaux, F. The CCAAT-binding complex controls respiratory gene expression and iron homeostasis in Candida glabrata. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Dongari-Bagtzoglou, A. Oral epithelium- Candida glabrata interactions in vitro. Oral Microbiol. Immunol. 2007, 22, 182–187. [Google Scholar] [CrossRef]
- Silva, S.; Henriques, M.C.; Hayes, A.; Oliveira, R.; Azeredo, J.; Williams, D.W. Candida glabrata and Candida albicans co-infection of an in vitro oral epithelium. J. Oral Pathol. Med. 2011, 40, 421–427. [Google Scholar] [CrossRef]
- Ho, H.L.; Haynes, K. Candida glabrata: New tools and technologies-expanding the toolkit. FEMS Yeast Res. 2015, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bai, J.; Wu, C.-L.; Wu, Y.; Zhao, Y.; Song, W.-M.; Luo, X.-P. Candida glabrata induced infection of rat tracheal epithelial cells is mediated by TLR-2 induced activation of NF-κB. Microb. Pathog. 2015, 91, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Zajac, D.; Karkowska-Kuleta, J.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Interaction of human fibronectin with Candida glabrata epithelial adhesin 6 (Epa6). Acta Biochim. Pol. 2016, 63, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dongari-Bagtzoglou, A. Epithelial GM-CSF induction by Candida glabrata. J. Dent. Res. 2009, 88, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Pärnänen, P.; Meurman, J.H.; Samaranayake, L.; Virtanen, I. Human oral keratinocyte E-cadherin degradation by Candida albicans and Candida glabrata. J. Oral Pathol. Med. 2010, 39, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Swidergall, M.; Solis, N.V.; Lionakis, M.S.; Filler, S.G. EphA2 is an epithelial cell pattern recognition receptor for fungal β-glucans. Nat. Microbiol. 2018, 3, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torrado, R.; Querol, A. Saccharomyces cerevisiae show low levels of traversal across the human blood brain barrier in vitro. F1000 Res. 2017, 6, 944. [Google Scholar] [CrossRef]
- West, L.; Lowman, D.W.; Mora-Montes, H.M.; Grubb, S.; Murdoch, C.; Thornhill, M.H.; Gow, N.A.R.; Williams, D.; Haynes, K. Differential virulence of Candida glabrata glycosylation mutants. J. Biol. Chem. 2013, 288, 22006–22018. [Google Scholar] [CrossRef]
- Torres-Tirado, D.; Knabb, M.; Castaño, I.; Patrón-Soberano, A.; De Las Peñas, A.; Rubio, R. Candida glabrata binds to glycosylated and lectinic receptors on the coronary endothelial luminal membrane and inhibits flow sense and cardiac responses to agonists. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R24–R32. [Google Scholar] [CrossRef]
- Netea, M.G.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Kullberg, B.J.; Van De Veerdonk, F.L. Immune defense against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Johnson, C.J.; Kernien, J.F.; Hoyer, A.R.; Nett, J.E. Mechanisms involved in the triggering of neutrophil extracellular traps (NETs) by Candida glabrata during planktonic and biofilm growth. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Essig, F.; Hünniger, K.; Dietrich, S.; Figge, M.T.; Kurzai, O. Human neutrophils dump Candida glabrata after intracellular killing. Fungal Genet. Biol. 2015, 84, 37–40. [Google Scholar] [CrossRef]
- Duggan, S.; Essig, F.; Hünniger, K.; Mokhtari, Z.; Bauer, L.; Lehnert, T.; Brandes, S.; Häder, A.; Jacobsen, I.D.; Martin, R.; et al. Neutrophil activation by Candida glabrata but not Candida albicans promotes fungal uptake by monocytes. Cell. Microbiol. 2015, 17, 1259–1276. [Google Scholar] [CrossRef] [PubMed]
- Ifrim, D.C.; Bain, J.M.; Reid, D.M.; Oosting, M.; Verschueren, I.; Gow, N.A.R.; van Krieken, J.H.; Brown, G.D.; Kullberg, B.J.; Joosten, L.A.B.; et al. Role of dectin-2 for host defense against systemic infection with Candida glabrata. Infect. Immun. 2014, 82, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Vitenshtein, A.; Charpak-Amikam, Y.; Yamin, R.; Bauman, Y.; Isaacson, B.; Stein, N.; Berhani, O.; Dassa, L.; Gamliel, M.; Gur, C.; et al. NK cell recognition of Candida glabrata through binding of NKp46 and NCR1 to fungal ligands Epa1, Epa6, and Epa7. Cell Host Microbe 2016, 20, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, C.; Majer, O.; Frohner, I.E.; Lesiak-Markowicz, I.; Hildering, K.-S.; Glaser, W.; Stockinger, S.; Decker, T.; Akira, S.; Muller, M.; et al. Conventional dendritic cells mount a Type I IFN response against Candida spp. requiring novel phagosomal TLR7-mediated IFN- Signaling. J. Immunol. 2011, 186, 3104–3112. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.N.; Balusu, S.; Gorityala, N.; Dandu, L.; Kaur, R. Functional genomic analysis of candida glabrata-macrophage interaction: Role of chromatin remodeling in virulence. PLoS Pathog. 2012, 8, e1002863. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Shen, H.; Zhang, T.; Huang, X.; Liu, X.Q.; Guo, S.Y.; Zhao, J.J.; Wang, C.F.; Yan, L.; Xu, G.T.; et al. Dectin-1 plays an important role in host defense against systemic Candida glabrata infection. Virulence 2017, 5594, 1–14. [Google Scholar] [CrossRef]
- Kasper, L.; Seider, K.; Gerwien, F.; Allert, S.; Brunke, S.; Schwarzmüller, T.; Ames, L.; Zubiria-Barrera, C.; Mansour, M.K.; Becken, U.; et al. Identification of Candida glabrata genes involved in pH modulation and modification of the phagosomal environment in macrophages. PLoS ONE 2014, 9, e96015. [Google Scholar] [CrossRef]
- Seider, K.; Gerwien, F.; Kasper, L.; Allert, S.; Brunke, S.; Jablonowski, N.; Schwarzmüller, T.; Barz, D.; Rupp, S.; Kuchler, K.; et al. Immune evasion, stress resistance, and efficient nutrient acquisition are crucial for intracellular survival of Candida glabrata within macrophages. Eukaryot. Cell 2014, 13, 170–183. [Google Scholar] [CrossRef]
- Brunke, S.; Seider, K.; Fischer, D.; Jacobsen, I.D.; Kasper, L.; Jablonowski, N.; Wartenberg, A.; Bader, O.; Enache-Angoulvant, A.; Schaller, M.; et al. One small step for a yeast—Microevolution within macrophages renders Candida glabrata hypervirulent due to a single point mutation. PLoS Pathog. 2014, 10, e1004478. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Candida glabrata | Candida albicans |
|---|---|---|
| Ploidy | Haploid | Diploid |
| Cellular morphology | Yeast | Yeast, pseudohyphae and hyphae |
| Cell size | 1–4 µm | 4–6 µm |
| Phylogeny | Non-CTG clade | CTG clade |
| Phenotypic switching | Present | Present |
| Carbon assimilation | Glucose and trehalose | Glucose, trehalose, maltose and galactose |
| Auxotrophy | Niacin, thiamine, pyridoxine | None |
| Crab tree | Positive | Negative |
| Mitochondrial function | Petite positive | Petite negative |
| Mating genes | Present | Present |
| Haem receptor | Absent | Present |
| Haemoglobin and transferrin utilization | Absent | Present |
| Innate azole resistance | Present | Absent |
| Secretory aspartyl proteases | Absent | Present |
| Life style | Probably commensal, and pathogenic | Commensal and pathogenic |
| Major sites of infection | Vaginal, oral, disseminated | Vaginal, oral, disseminated |
| Major adhesins | Lectins (Epa) | Lectins (Als and Hwp) |
| Biofilm | Present | Present |
| Invasion | Not known | Induced endocytosis and active penetration |
| Damage to host cells | No significant damage | Substantial damage |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, K.; Askari, F.; Sahu, M.S.; Kaur, R. Candida glabrata: A Lot More Than Meets the Eye. Microorganisms 2019, 7, 39. https://doi.org/10.3390/microorganisms7020039
Kumar K, Askari F, Sahu MS, Kaur R. Candida glabrata: A Lot More Than Meets the Eye. Microorganisms. 2019; 7(2):39. https://doi.org/10.3390/microorganisms7020039
Chicago/Turabian StyleKumar, Kundan, Fizza Askari, Mahima Sagar Sahu, and Rupinder Kaur. 2019. "Candida glabrata: A Lot More Than Meets the Eye" Microorganisms 7, no. 2: 39. https://doi.org/10.3390/microorganisms7020039
APA StyleKumar, K., Askari, F., Sahu, M. S., & Kaur, R. (2019). Candida glabrata: A Lot More Than Meets the Eye. Microorganisms, 7(2), 39. https://doi.org/10.3390/microorganisms7020039