Identification and Toxigenic Potential of Fungi Isolated from Capsicum Peppers

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
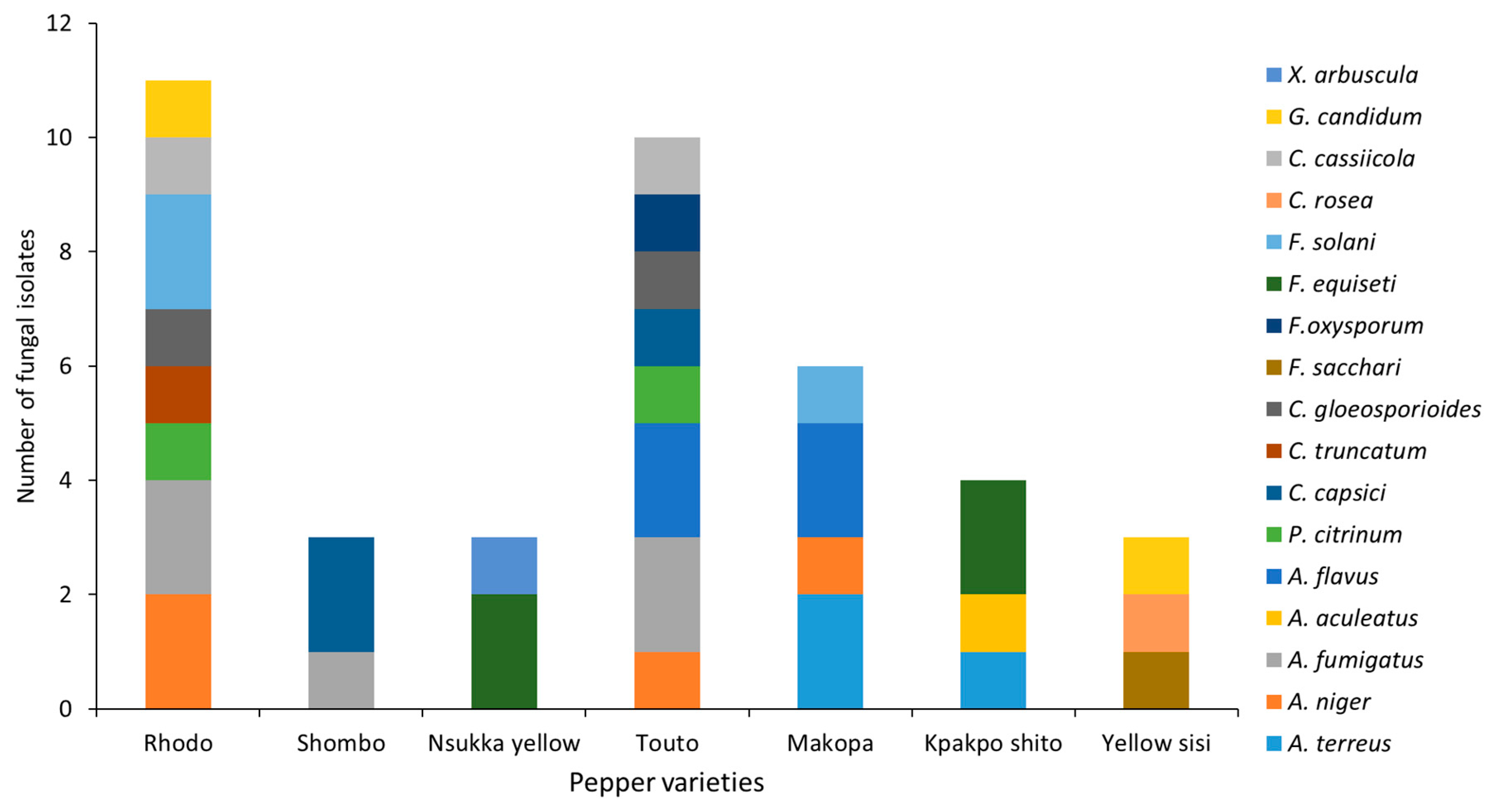
2.1. Sample Collection and Isolation of Fungi
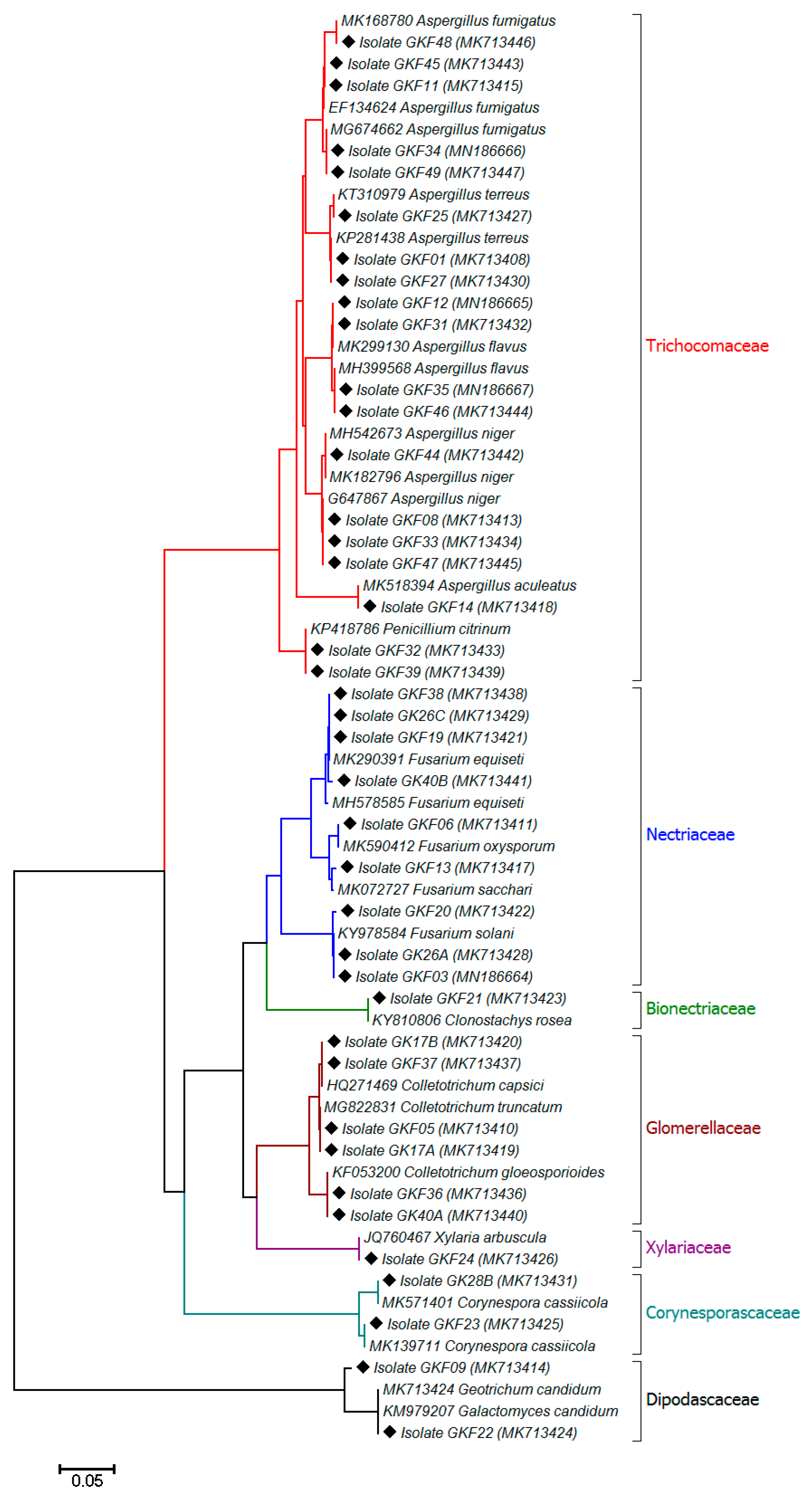
2.2. Morphological and Molecular Identification of the Fungi
2.3. Pathogenicity Test
2.4. Mycotoxins Analysis
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonsu, K.O.; Oduro, C.A.; Tetteh, J.P. Traditional cocoa agroforestry: Species encountered in the cocoa ecosystem of a typical cocoa growing district in Ghana. In Proceedings of the 14th Internationald Cocoa Research Conference, Accra, Ghana, 13–18 October 2003. [Google Scholar]
- Diao, Y.-Z.; Zhang, C.; Liu, F.; Wang, W.-Z.; Liu, L.; Cai, L.; Liu, X.-L. Colletotrichum species causing anthracnose disease of chili in China. Persoonia–Mol. Phylogeny Evol. Fungi 2017, 38, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Caires, N.P.; Pinho, D.B.; Souza, J.S.C.; Silva, M.A.; Lisboa, D.O.; Pereira, O.L.; Furtado, G.Q. First report of Anthracnose on pepper fruit caused by Colletotrichum scovillei in Brazil. Plant Dis. 2014, 98, 1437. [Google Scholar] [CrossRef] [PubMed]
- Widodo; Hidayat, S.H. Identification of Colletotrichum species associated with chili Anthracnose in Indonesia by morphological characteristics and Species-Specific Primers. Asian J. Plant Pathol. 2018, 12, 7–15. [Google Scholar] [CrossRef]
- Than, P.P.; Jeewon, R.; Hyde, K.D.; Pongsupasamit, S.; Mongkolporn, O.; Taylor, P.W.J. Characterization and pathogenicity of Colletotrichum species associated with anthracnose on chilli (Capsicum spp.) in Thailand. Plant Pathol. 2008, 57, 562–572. [Google Scholar] [CrossRef]
- Oo, M.M.; Lim, G.; Jang, H.A.; Oh, S.-K. Characterization and pathogenicity of new record of Anthracnose on various chili varieties caused by Colletotrichum scovillei in Korea. Mycobiology 2017, 45, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Lema, A.A.; Mudansiru, A.; Alexander, B.; Sakinatu, M.; Sakinatu, M. Evaluation of fungal species isolated from three different varieties of pepper (Capsicum chinense, C. frutescens and C. annum L.) in Dutsinma, Katsina State. Ann. Biol. Sci. 2018, 6, 13–17. [Google Scholar] [CrossRef]
- Fatimoh, A.O.; Moses, A.A.; Adekunle, O.B.; Dare, O.E. Isolation and identification of rot fungi on post-harvest of pepper (Capsicum annuum L.) fruits. J. Biol. 2017, 3, 24–29. [Google Scholar]
- Nsabiyera, V.; Ochwo-Ssemakula, M.; Sseruwagi, P. Hot pepper reaction to field diseases. African Crop. Sci. J. 2012, 20, 77–97. [Google Scholar]
- Miller, J.D. Fungi and Mycotoxins in Grain: Implications for Stored Product Research. J. Stored Prod. Res. 1995, 31, 1–16. [Google Scholar] [CrossRef]
- Mandeel, Q.A. Fungal contamination of some imported spices. Mycopathologia 2005, 159, 291–298. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Benito, M.J.; Martín, A.; Aranda, E.; Hernández, A.; Córdoba, M.G. Characterization of molds isolated from smoked paprika by PCR-RFLP and micellar electrokinetic capillary electrophoresis. Food Microbiol. 2009, 26, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Marín, S.; Mateo, E.M.; Gil-Serna, J.; Valle-Algarra, F.M.; Patiño, B.; Ramos, A.J. Mycobiota and co-occurrence of mycotoxins in Capsicum powder. Int. J. Food Microbiol. 2011, 151, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Q.; Amjad, M.; Asi, M.R.; Ariño, A. Assessment of hot peppers for aflatoxin and mold proliferation during storage. J. Food Prot. 2011, 74, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Cotty, P.J. Aflatoxin contamination of dried red chilies: Contrasts between the United States and Nigeria, two markets differing in regulation enforcement. Food Control. 2017, 80, 374–379. [Google Scholar] [CrossRef]
- Costa, J.; Rodríguez, R.; Garcia-Cela, E.; Medina, A.; Magan, N.; Lima, N.; Battilani, P.; Santos, C. Overview of fungi and mycotoxin contamination in Capsicum pepper and in its derivatives. Toxins 2019, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins; IARC Scientific Publication: Lyon, France, 1993; pp. 19–23. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Available online: http://www.fao.org/fao-who-codexalimentarius (accessed on 21 July 2019).
- de Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of clinical fungi; de Hoog, G.S., Guarro, J., Gené, J., Figueras, M.J., Eds.; Centraalbureau voor Schimmelcultures: Baarn, The Netherlands, 1995. [Google Scholar]
- Sadhasivam, S.; Britzi, M.; Zakin, V.; Kostyukovsky, M.; Trostanetsky, A.; Quinn, E.; Sionov, E. Rapid detection and identification of mycotoxigenic fungi and mycotoxins in stored wheat grain. Toxins 2017, 9, 302. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Gell, R.M.; Carbone, I. HPLC quantitation of aflatoxin B1 from fungal mycelium culture. J. Microbiol. Methods 2019, 158, 14–17. [Google Scholar] [CrossRef]
- Cambaza, E.; Koseki, S.; Kawamura, S. Fusarium graminearum colors and Deoxynivalenol synthesis at different water activity. Foods 2019, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Maor, U.; Sadhasivam, S.; Zakin, V.; Prusky, D.; Sionov, E. The effect of ambient pH modulation on ochratoxin A accumulation by Aspergillus carbonarius. World Mycotoxin J. 2017, 10, 339–348. [Google Scholar] [CrossRef]
- Guo, W.; Zhao, M.; Chen, Q.; Huang, L.; Mao, Y.; Xia, N.; Teng, J.; Wei, B. Citrinin produced using strains of Penicillium citrinum from Liupao tea. Food Biosci. 2019, 28, 183–191. [Google Scholar] [CrossRef]
- Pena, G.A.; Pereyra, C.M.; Armando, M.R.; Chiacchiera, S.M.; Magnoli, C.E.; Orlando, J.L.; Dalcero, A.M.; Rosa, C.A.R.; Cavaglieri, L.R. Aspergillus fumigatus toxicity and gliotoxin levels in feedstuff for domestic animals and pets in Argentina. Lett. Appl. Microbiol. 2010, 50, 77–81. [Google Scholar] [CrossRef]
- Udoh, I.P.; Eleazar, C.I.; Ogeneh, B.O.; Ohanu, M.E. Studies on fungi responsible for the spoilage/deterioration of some edible fruits and vegetables. Adv. Microbiol. 2015, 05, 285–290. [Google Scholar] [CrossRef]
- Adebanjo, A.; Shopeju, E. Sources and Mycoflora associated with some sundried vegetables in storage. Int. Biodeterior. Biodegradation 1993, 31, 255–263. [Google Scholar] [CrossRef]
- Dixon, L.J.; Schlub, R.L.; Pernezny, K.; Datnoff, L.E. Host specialization and phylogenetic diversity of Corynespora cassiicola. Phytopathology 2009, 99, 1015–1027. [Google Scholar] [CrossRef]
- Ferreira, A.F.T.A.F.e.; da Silva Bentes, J.L. Pathogenicity of Corynespora cassiicola on different hosts in Amazonas State, Brazil. Summa Phytopathol. 2017, 43, 63–65. [Google Scholar] [CrossRef]
- Ahmed, F.A.; Alam, N.; Khair, A. Incidence and biology of Corynespora cassiicola (Berk. & Curt.) Wei. disease of okra in Bangladesh. Bangladesh J. Bot. 2014, 42, 265–272. [Google Scholar]
- Fulmer, A.M.; Walls, J.T.; Dutta, B.; Parkunan, V.; Brock, J.; Kemerait, R.C. First report of target spot caused by Corynespora cassiicola on cotton in Georgia. Plant. Dis. 2012, 96, 1066. [Google Scholar] [CrossRef]
- Furukawa, T.; Ushiyama, K.; Kishi, K. Corynespora leaf spot of scarlet sage caused by Corynespora cassiicola. J. Gen. Plant Pathol. 2008, 74, 117–119. [Google Scholar] [CrossRef]
- Shimomoto, Y.; Adachi, R.; Morita, Y.; Yano, K.; Kiba, A.; Hikichi, Y.; Takeuchi, S. Corynespora blight of sweet pepper (Capsicum annuum) caused by Corynespora cassiicola (Berk. & Curt.) Wei. J. Gen. Plant Pathol. 2008, 74, 335–337. [Google Scholar]
- Chai, A.-L.; Du, G.-F.; Shi, Y.-X.; Xie, X.-W.; Li, B.-J. Corynespora spot of hot pepper caused by Corynespora cassiicola in China. Can. J. Plant Pathol. 2014, 36, 407–411. [Google Scholar] [CrossRef]
- Omolo, M.A.; Wong, Z.-Z.; Borh, W.G.; Hedblom, G.A.; Dev, K.; Baumler, D.J. Comparative analysis of capsaicin in twenty nine varieties of unexplored Capsicum and its antimicrobial activity against bacterial and fungal pathogens. J. Med. Plants Res. 2018, 12, 544–556. [Google Scholar]
- Cichewicz, R.H.; Thorpe, P.A. The antimicrobial properties of chile peppers (Capsicum species) and their uses in Mayan medicine. J. Ethnopharmacol. 1996, 52, 61–70. [Google Scholar] [CrossRef]
- Varga, J.; Frisvad, J.C.; Samson, R.A. Two new aflatoxin producing species, and an overview of Aspergillus section Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, S.M.; Mäki-Paakkanen, J.; Hirvonen, M.-R.; Roponen, M.; von Wright, A. Genotoxicity of gliotoxin, a secondary metabolite of Aspergillus fumigatus, in a battery of short-term test systems. Mutat. Res. Toxicol. Environ. Mutagen. 2002, 520, 161–170. [Google Scholar] [CrossRef]
- Bouakline, A.; Lacroix, C.; Roux, N.; Gangneux, J.P.; Derouin, F. Fungal contamination of food in hematology units. J. Clin. Microbiol. 2000, 38, 4272–4273. [Google Scholar]
- Kabak, B.; Dobson, A.D.W. Mycotoxins in spices and herbs–An update. Crit. Rev. Food Sci. Nutr. 2017, 57, 18–34. [Google Scholar] [CrossRef]
- Gambacorta, L.; Magistà, D.; Perrone, G.; Murgolo, S.; Logrieco, A.F.; Solfrizzo, M. Co-occurrence of toxigenic moulds, aflatoxins, ochratoxin A, Fusarium and Alternaria mycotoxins in fresh sweet peppers (Capsicum annuum) and their processed products. World Mycotoxin J. 2018, 11, 159–174. [Google Scholar] [CrossRef]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.; De Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A.; et al. Mycotoxigenic potentials of Fusarium species in various culture matrices revealed by mycotoxin profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate | Species | Mycotoxins (ng/g agar ± SD) a | ||||
|---|---|---|---|---|---|---|
| AFB1 | OTA | Gliotoxin | DON | Citrinin | ||
| GKF12 | A. flavus | 49.3 ± 3.2 | - | - | - | - |
| GKF31 | A. flavus | nd | - | - | - | - |
| GKF35 | A. flavus | nd | - | - | - | - |
| GKF46 | A. flavus | nd | - | - | - | - |
| GKF08 | A. niger | - | nd | - | - | - |
| GKF33 | A. niger | - | nd | - | - | - |
| GKF44 | A. niger | - | nd | - | - | - |
| GKF47 | A. niger | - | nd | - | - | - |
| GKF11 | A. fumigatus | - | - | 1015.55 ± 30.3 | - | - |
| GKF34 | A. fumigatus | - | - | 1917.17 ± 61.5 | - | - |
| GKF45 | A. fumigatus | - | - | 1260.13 ± 42.6 | - | - |
| GKF48 | A. fumigatus | - | - | 1394.98 ± 38.5 | - | - |
| GKF49 | A. fumigatus | - | - | 1318.23 ± 41.1 | - | - |
| GKF03 | F. solani | - | - | - | 8127.85 ± 196.2 | - |
| GKF20 | F. solani | - | - | - | 6744.66 ± 150.7 | - |
| GK26A | F. solani | - | - | - | 3903.92 ± 109.3 | - |
| GKF19 | F. equiseti | - | - | - | 3780.28 ± 93.01 | - |
| GK26C | F. equiseti | - | - | - | nd | - |
| GKF38 | F. equiseti | - | - | - | nd | - |
| GK40B | F. equiseti | - | - | - | nd | - |
| GKF32 | P. citrinum | - | - | - | - | 273.25 ± 13.5 |
| GKF39 | P. citrinum | - | - | - | - | nd |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frimpong, G.K.; Adekunle, A.A.; Ogundipe, O.T.; Solanki, M.K.; Sadhasivam, S.; Sionov, E. Identification and Toxigenic Potential of Fungi Isolated from Capsicum Peppers. Microorganisms 2019, 7, 303. https://doi.org/10.3390/microorganisms7090303
Frimpong GK, Adekunle AA, Ogundipe OT, Solanki MK, Sadhasivam S, Sionov E. Identification and Toxigenic Potential of Fungi Isolated from Capsicum Peppers. Microorganisms. 2019; 7(9):303. https://doi.org/10.3390/microorganisms7090303
Chicago/Turabian StyleFrimpong, Gabriel Kojo, Adedotun Adeyinka Adekunle, Oluwatoyin Temitayo Ogundipe, Manoj Kumar Solanki, Sudharsan Sadhasivam, and Edward Sionov. 2019. "Identification and Toxigenic Potential of Fungi Isolated from Capsicum Peppers" Microorganisms 7, no. 9: 303. https://doi.org/10.3390/microorganisms7090303
APA StyleFrimpong, G. K., Adekunle, A. A., Ogundipe, O. T., Solanki, M. K., Sadhasivam, S., & Sionov, E. (2019). Identification and Toxigenic Potential of Fungi Isolated from Capsicum Peppers. Microorganisms, 7(9), 303. https://doi.org/10.3390/microorganisms7090303