Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand



Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Yeast Isolation
2.2. Yeast Identification
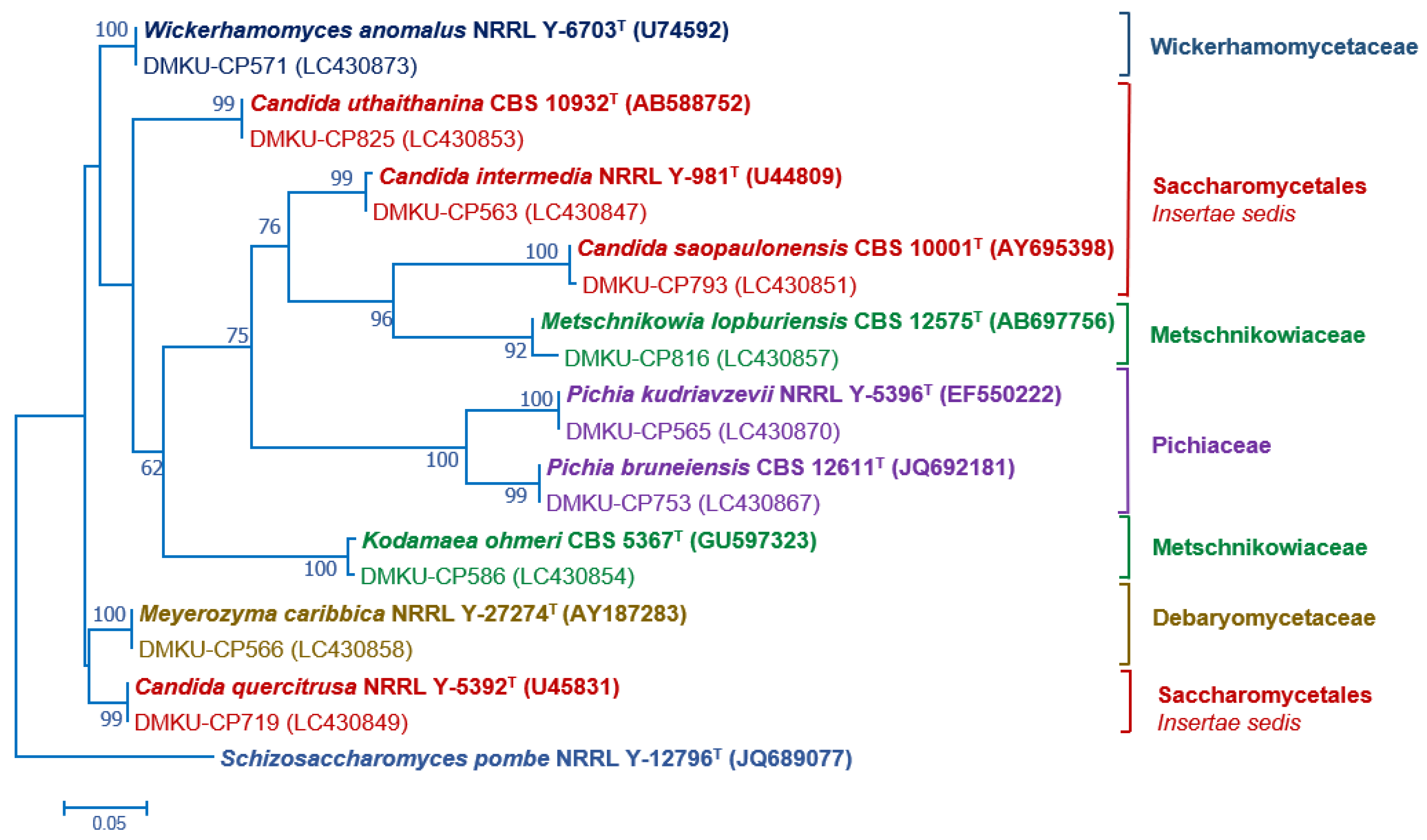
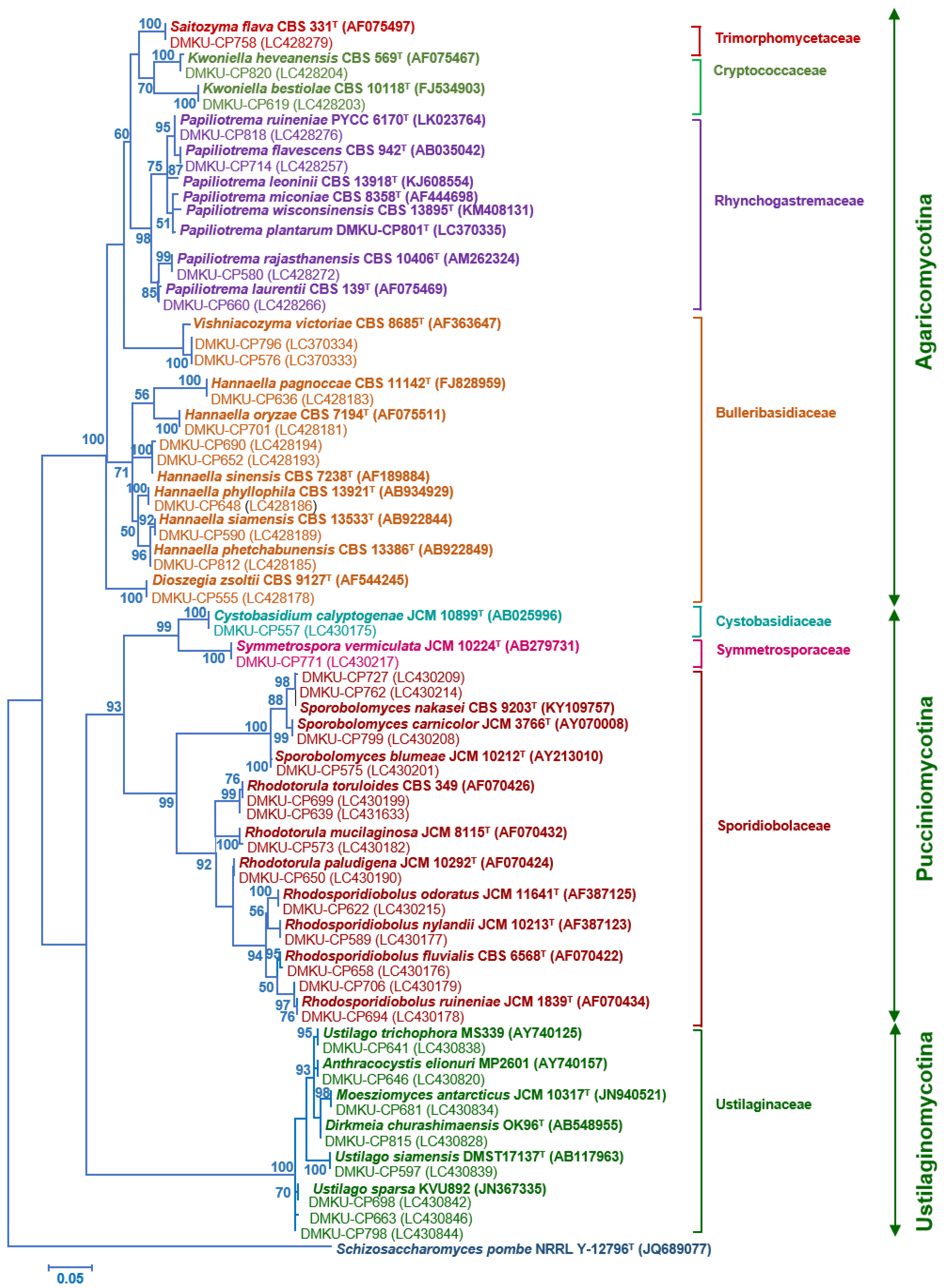
2.3. Phylogenetic Analysis
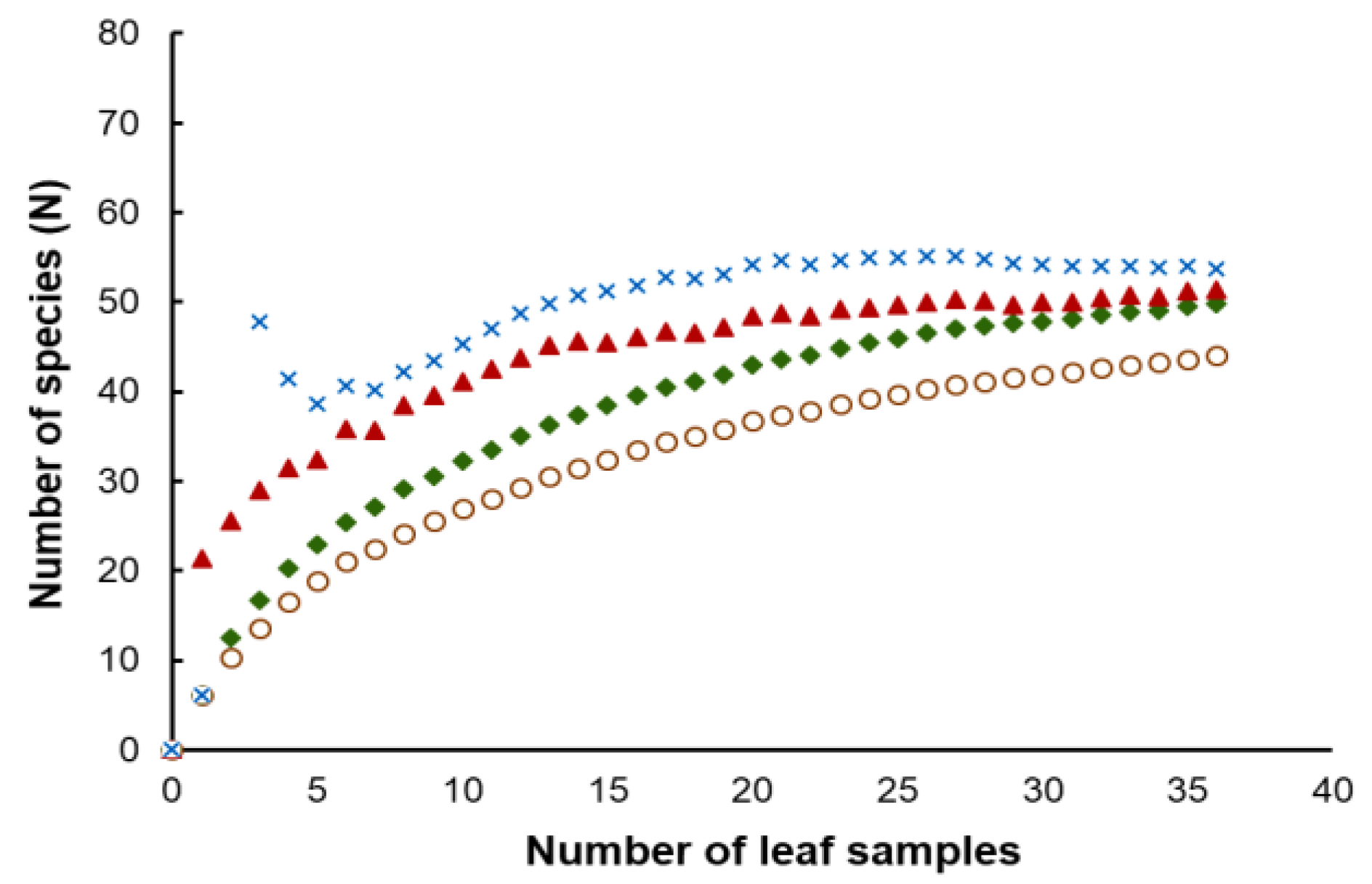
2.4. Biodiversity Analyses
3. Results
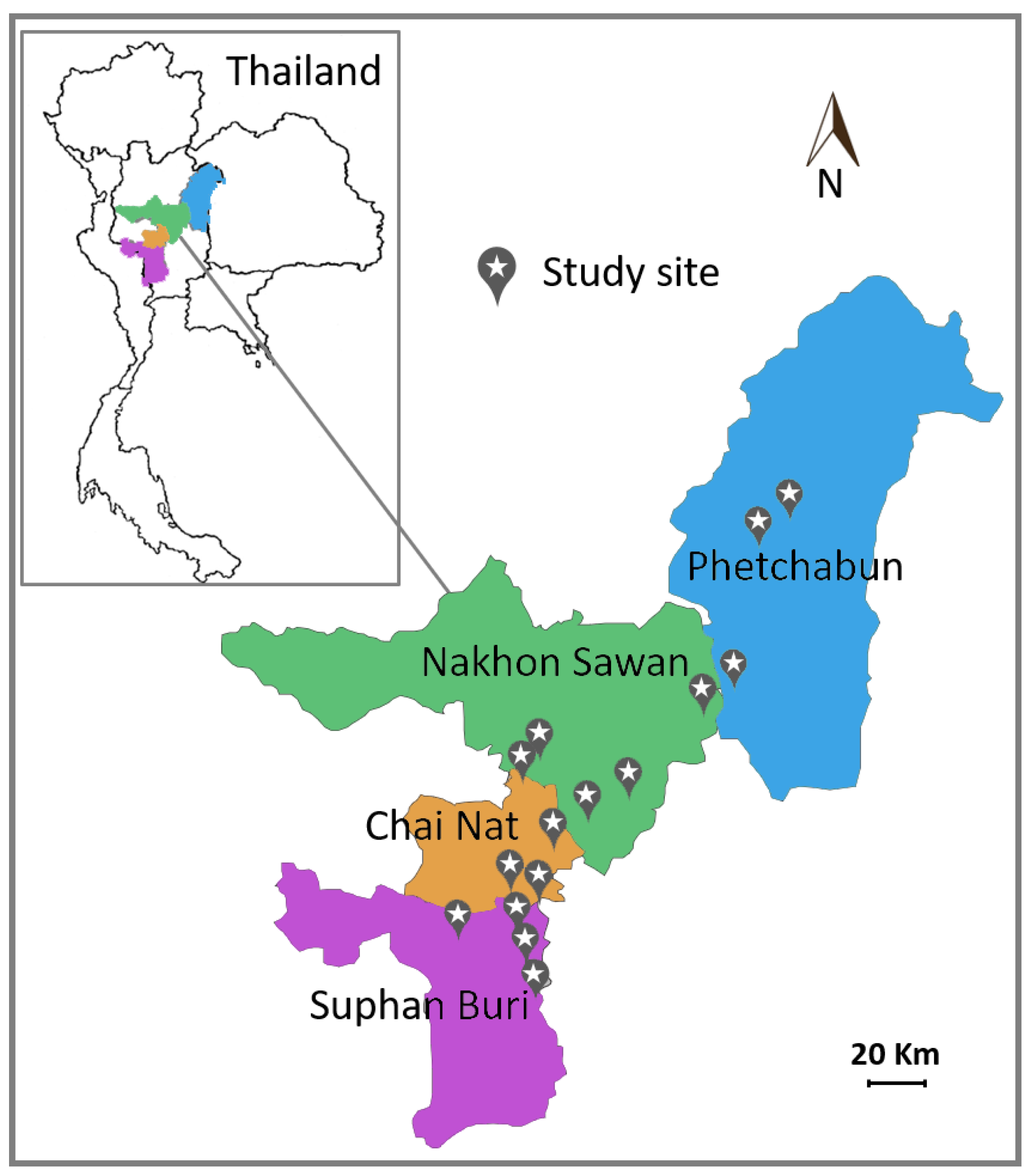
3.1. Sample Collection and Yeast Isolation
3.2. Yeast Identification
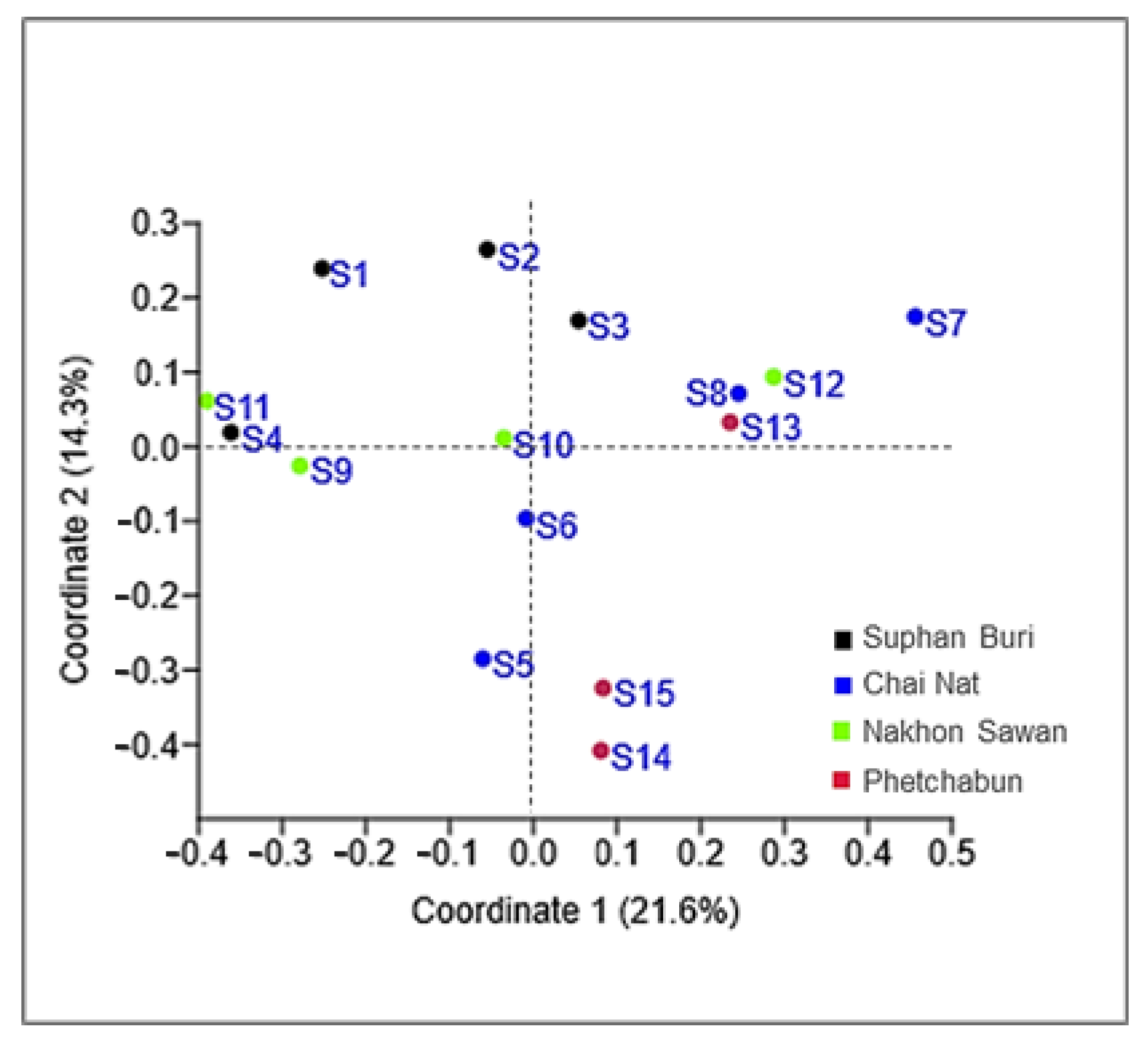
3.3. Yeast Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whipps, J.; Hand, P.; Pink, D.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 105, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Phaff, H.J.; Starmer, W.T. Yeasts associated with plants, insects and soil. In The Yeasts, 2nd ed.; Rose, A.H., Harrison, J.S., Eds.; Academic Press: London, UK, 1987; pp. 123–180. [Google Scholar]
- Fonseca, Á.; Inácio, J. Phylloplane yeasts. In Biodiversity and Ecophysiology of Yeasts; Rosa, C., Péter, G., Eds.; Springer Science: Berlin/Heidelberg, Germany, 2006; pp. 263–301. [Google Scholar]
- Fiala, V.; Glad, C.; Martin, M.; Jolivet, E.; Derridj, S. Occurrence of soluble carbohydrates on thephylloplane of maize (Zea mays L.): Variations in relation to leaf heterogeneity and position on the plant. New Phytol. 1990, 115, 609–615. [Google Scholar] [CrossRef]
- Xin, G.; Glawe, D.; Doty, S.L. Characterization of three endophytic, indole-3-acetic acidproducing yeasts occurring in Populus trees. Mycol. Res. 2009, 113, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.H.; Harris, R.E. The ecology and biogeography of microorganisms on plantsurfaces. Ann. Rev. Phytopathol. 2000, 38, 145–180. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- de Azeredo, L.A.I.; Gomes, E.A.T.; Mendonca-Hagler, L.C.; Hagler, A.N. Yeast communities associated with sugarcane in Campos, Rio de Janeiro, Brazil. Int. Microbiol. 1998, 1, 205–208. [Google Scholar]
- Glushakova, A.M.; Yurkov, A.M.; Chernov, I.Y. Massive isolation of anamorphous ascomycete yeasts Candida oleophila from plant phyllosphere. Microbiology 2007, 76, 799–803. [Google Scholar] [CrossRef]
- Peter, G.; Tornai-Lehoczki, J.; Dlauchy, D. Ogataea allantospora sp. nov., anascomycetous yeast species from phylloplane. Antonie Leeuwenhoek 2007, 92, 443–448. [Google Scholar] [CrossRef]
- Slavikova, E.; Vadkertiova, R.; Vranova, D. Yeasts colonizing the leaves of fruit trees. Ann. Microbiol. 2009, 59, 419–424. [Google Scholar] [CrossRef]
- Inácio, J.; Portugal, L.; Spencer-Martins, I.; Fonseca, Á. Phylloplane yeasts from Portugal: Seven novel anamorphic species in the Tremellales lineage of the Hymenomycetes (Basidiomycota) producing orange-coloured colonies. FEMS Yeast Res. 2005, 5, 1167–1183. [Google Scholar] [CrossRef]
- Nakase, T.; Takashima, M.; Itoh, M.; Fungsin, B.; Potacharoen, W.; Atthasampunna, P.; Komagata, K. Ballistoconidium-forming yeasts found in the phyllosphere of Thailand. Microbiol. Cult. Collect. 2001, 17, 23–33. [Google Scholar]
- Santos, A.R.; Faria, E.S.; Lachance, M.A.; Rosa, C.A. Ogataea mangiferae sp. nov., a methylotrophic yeast isolated from mango leaves. Int. J. Syst. Evol. Microbiol. 2015, 65, 1855–1859. [Google Scholar] [CrossRef] [PubMed]
- Koowadjanakul, N.; Jindamorakot, S.; Yongmanitchai, W.; Limtong, S. Ogataea phyllophila sp. nov., Candida chumphonensis sp. nov. and Candida mattranensis sp. nov., three methylotrophic yeast species from phylloplane in Thailand. Antonie Leeuwenhoek 2011, 100, 207–217. [Google Scholar] [CrossRef]
- Limtong, S.; Kaewwichian, R.; Groenewald, M. Ogataea kanchanaburiensis sp. nov. and Ogataea wangdongensis sp. nov., two novel methylotrophic yeast species from phylloplane in Thailand. Antonie Leeuwenhoek 2013, 103, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Limtong, S.; Kaewwichian, R.; Yongmanitchai, W.; Kawasaki, H. Diversity of culturable yeasts in phylloplane of sugarcane in Thailand and their capability to produce indole-3-acetic acid. World J. Microbiol. Biotechnol. 2014, 30, 1785–1796. [Google Scholar] [CrossRef]
- Limtong, S.; Kaewwichian, R. Candida phyllophila sp. nov. and Candida vitiphila sp. nov., two novel yeast species from grape phylloplane in Thailand. J. Gen. Appl. Microbiol. 2013, 59, 191–197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Limtong, S.; Kaewwichian, R. The diversity of culturable yeasts in the phylloplane of rice in Thailand. Ann. Microbiol. 2015, 65, 667–675. [Google Scholar] [CrossRef]
- Nasanit, R.; Krataithong, K.; Tantirungkij, M.; Limtong, S. Assessment of epiphytic yeast diversity in rice (Oryza sativa) phyllosphere in Thailand by a culture-independent approach. Antonie Leeuwenhoek 2015, 107, 1475–1490. [Google Scholar] [CrossRef]
- Nasanit, R.; Tangwong-o-thai, A.; Tantirungkij, M.; Limtong, S. The assessment of epiphytic yeast diversity in sugarcane phyllosphere in Thailand by culture-independent method. Fungal Biol. 2015, 119, 1145–1157. [Google Scholar] [CrossRef]
- Nasanit, R.; Jaibangyang, S.; Tantirungkij, M.; Limtong, S. Yeast diversity and novel yeast D1/D2 sequences from corn phylloplane obtained by a culture-independent approach. Antonie Leeuwenhoek 2016, 109, 1615–1634. [Google Scholar] [CrossRef]
- Srisuk, N.; Nutaratat, P.; Surussawadee, J.; Limtong, S. Yeast Communities in Sugarcane Phylloplane. Microbiology (Mosc.) 2019, 88, 353–369. [Google Scholar] [CrossRef]
- Yurkov, A.; Inácio, J.; Chernov, I.; Fonseca, Á. Yeast biogeography and the effects of species recognition approaches: The case study of widespread basidiomycetous species from birch forests in Russia. Curr. Microbiol. 2015, 70, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Jumpponen, A.; Jones, K.L. Seasonally dynamic fungal communities in the Quercus macrocarpa phyllosphere differ between urban and nonurban environments. New Phytol. 2010, 186, 496–513. [Google Scholar] [CrossRef] [PubMed]
- Glushakova, A.M.; Chernov, I.Y. Seasonal dynamic of the numbers of epiphytic yeast communities. Microbiology 2010, 79, 830–839. [Google Scholar] [CrossRef]
- Surussawadee, J.; Jindamorakot, S.; Nakase, T.; Lee, C.F.; Limtong, S. Hannaella phyllophila sp. nov., a novel basidiomycetous yeast species associated with plants in Thailand and Taiwan. Int. J. Syst. Evol. Microbiol. 2015, 65, 2135–2140. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Kemler, M.; Begerow, D. Species accumulation curves and incidence-based species richness estimators to appraise the diversity of cultivable yeasts from beech forest soils. PLoS ONE 2011, 6, e23671. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Kemler, M.; Begerow, D. Assessment of yeast diversity in soils under different management regimes. Fungal Ecol. 2012, 5, 24–35. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Gadanho, M.; Santos, S. Polyphasic taxonomy of the basidiomycetous yeast genus Rhodosporidium: Rhodosporidium kratochvilovae and related anamorphic species. Int. J. Syst. Evol. Microbiol. 2001, 51, 687–697. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph, Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycete yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Into, P.; Pontes, A.; Jacques, N.; Casaregola, S.; Limtong, S.; Sampaio, J.P. Papiliotrema plantarum sp. nov., a novel tremellaceous sexual yeast species. Int. J. Syst. Evol. Microbiol. 2018, 68, 1937–1941. [Google Scholar] [CrossRef]
- Li, M.X. Studies on Sporobolomycetaceae I. Taxonomy of Bullera. Acta Microbiol. Sin. 1982, 36, 17–25. [Google Scholar]
- Nakase, T.; Suzuki, M. Ballistospore-forming yeasts found on the surface of the Japanese rice plants Oryza sativa L. J. Gen. Appl. Microbiol. 1985, 31, 457–474. [Google Scholar] [CrossRef]
- Bai, F.Y.; Takashima, M.; Nakase, T. Description of Bullera kunmingensis sp. nov., and clarification of the taxonomic status of Bullera sinensis and its synonyms based on molecular phylogenetic analysis. FEMS Yeast Res. 2001, 1, 103–109. [Google Scholar] [CrossRef][Green Version]
- Hoshino, Y.T.; Matsumoto, N. Changes in fungal community structure in bulk soil and spinach rhizosphere soil after chemical fumigation as revealed by 18S rDNA PCR-DGGE. Soil Sci. Plant Nutr. 2007, 53, 40–55. [Google Scholar] [CrossRef]
- Chou, J.Y.; Chen, H.W.; Lin, C.C.; Wen, Y.D.; Wang, W.L. Yeast diversity associated with the biting midge Forcipomyia taiwana in Taiwan. Nova Hedwigia 2015, 101, 519–527. [Google Scholar] [CrossRef]
- Camargo, F.P.; Araujo, A.C.V.; de Moraes, E.M.; Dos Santos, A.C.A. A comparison between cactophilic yeast communities isolated from Cereus hildmannianus and Praecereus euchlorus necrotic cladodes. Fungal Biol. 2016, 120, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Khunnamwong, P.; Jindamorakot, S.; Limtong, S. Endophytic yeast diversity in leaf tissue of rice, corn and sugarcane cultivated in Thailand assessed by a culture-dependent approach. Fungal Biol. 2018, 122, 785–799. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Vustin, M.M.; Tyaglov, B.V.; Maksimova, I.A.; Sineokiy, S.P. Pigmented basidiomycetous yeasts are a promising source of carotenoids and ubiquinone Q10. Microbiology 2008, 77, 1–6. [Google Scholar] [CrossRef]
- Morita, T.; Ogura, Y.; Takashima, M.; Hirose, N.; Fukuoka, T.; Imura, T.; Kondo, Y.; Kitamoto, D. Isolation of Pseudozyma churashimaensis sp. nov., a novel ustilaginomycetous yeast species as a producer of glycolipid biosurfactants, mannosyl erythritol lipids. J. Biosci. Bioeng. 2011, 112, 137–144. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Röhl, O.; Pontes, A.; Carvalho, C.; Maldonado, C.; Sampaio, J.P. Local climatic conditions constrain soil yeast diversity patterns in Mediterranean forests, woodlands and scrub biome. FEMS Yeast Res. 2016, 16, fov103. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chi, Z.; Liu, G.L.; Madzak, C.; Chi, Z.M. Both decrease in ACL1 gene expression and increase in ICL1 gene expression in marine-derived yeast Yarrowia lipolytica expressing INU1 gene enhance citric acid production from inulin. Mar. Biotechnol. 2013, 15, 26–36. [Google Scholar] [CrossRef]
- Wu, J.; Hu, J.; Zhao, S.; He, M.; Hu, G.; Ge, X.; Peng, N. Single-cell protein and xylitol production by a novel yeast strain Candida intermedia FL023 from lignocellulosic hydrolysates and xylose. Appl. Biochem. Biotechnol. 2018, 185, 163–178. [Google Scholar] [CrossRef]
- Rachamontree, P.; Phusantisampan, T.; Woravutthikul, N.; Pornwongthong, P.; Sriariyanun, M. Selection of Pichia kudriavzevii Strain for the Production of Single-Cell Protein from Cassava Processing Waste. World Acad. Sci. Eng. Technol. Int. J. Biol. Food Veter. Agric. Eng. 2015, 9, 460–464. [Google Scholar]
- Robak, K.; Balcerek, M. Review of second generation bioethanol production from residual biomass. Food Technol. Biotechnol. 2018, 56, 174. [Google Scholar] [CrossRef] [PubMed]
- Yuangsaard, N.; Yongmanitchai, W.; Yamada, M.; Limtong, S. Selection and characterization of a newly isolated thermotolerant Pichia kudriavzevii strain for ethanol production at high temperature from cassava starch hydrolysate. Antonie Leeuwenhoek 2013, 103, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, G.L.; Chi, Z.; Chi, Z.M. Single cell oil production from hydrolysate of cassava starch by marine-derived yeast Rhodotorula mucilaginosa TJY15a. Biomass Bioenerg. 2010, 34, 101–107. [Google Scholar] [CrossRef]
- Poontawee, R.; Yongmanitchai, W.; Limtong, S. Efficient oleaginous yeasts for lipid production from lignocellulosic sugars and effects of lignocellulose degradation compounds on growth and lipid production. Process Biochem. 2017, 53, 44–60. [Google Scholar] [CrossRef]
- Matsui, T.; Otsuka, K.Y.; Sato, S. Microbial oil production from carbohydrates using Sporobolomyces carnicolor strain O33. Ann. Microbiol. 2012, 62, 861–864. [Google Scholar] [CrossRef]
- Nutaratat, P.; Amsri, W.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Indole-3-acetic acid production by newly isolated red yeast Rhodosporidium paludigenum. J. Gen. Appl. Microbiol. 2015, 61, 1–9. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Production of itaconic acid by Pseudozyma antarctica NRRL Y-7808 under nitrogen-limited growth conditions. Enzym. Microb. Technol. 2006, 39, 824–827. [Google Scholar] [CrossRef]
- Do, D.T.H.; Theron, C.W.; Fickers, P. Organic Wastes as Feedstocks for Non-Conventional Yeast-Based Bioprocesses. Microorganisms 2019, 7, 229. [Google Scholar] [CrossRef]
- Carneiro, C.V.G.; de Paula e Silva, F.C.; Almeida, J.R. Xylitol Production: Identification and Comparison of New Producing Yeasts. Microorganisms 2019, 7, 484. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rywińska, A.; Gładkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef]
- Trichez, D.; Steindorff, A.S.; Soares1, C.E.V.F.; Formighieri, E.F.; Almeida, J.R.M. Physiological and comparative genomic analysis of new isolated 2 yeasts Spathaspora sp. JA1 and Meyerozyma caribbica JA9 reveal 3 insights into xylitol production. FEMS Yeast Res. 2019, 19, foz034. [Google Scholar] [CrossRef] [PubMed]
- Stratilová, B.; Klaudiny, J.; Řehulka, P.; Stratilová, E.; Mészárosová, C.; Garajová, S.; Pavlatovská, B.; Řehulková, H.; Kozmon, S.; Šesták, S.; et al. Characterization of a long-chain α-galactosidase from Papiliotrema flavescens. World J. Microbiol. Biotechnol. 2018, 34, 19. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | No. of Sample | No. of Strain | |||||
|---|---|---|---|---|---|---|---|
| Site | Province District | Location | Date (dd mm yy) | Atmospheric Temperature a (°C) | Rainfall b (mm) | ||
| Suphan Buri | |||||||
| S1 | Doembangnangbuat | 14°50′27.1″ N 100°07′51.5″ E | 2016-07-18 | 29.2–29.8 | 100.2–157.0 | 2 | 19 |
| S2 | Sam Chuk | 14°47′18.4″ N 100°09′14.1″ E | 2016-07-30 | 3 | 22 | ||
| S3 | Nong Ya Sai | 14°49′22.6″ N 99°55′46.0″ E | 2016-08-06 | 2 | 9 | ||
| S4 | Si Prachan | 14°38′36.8″ N 100°08′53.1″ E | 2016-08-06 | 2 | 13 | ||
| Total | 9 | 63 | |||||
| Chai Nat | |||||||
| S5 | Hankha | 14°59′55.3″ N 100°05′45.6″ E | 2016-08-06 | 28.1–28.6 | 208.8–358.6 | 4 | 22 |
| S6 | Sankhaburi | 14°57′37.8″ N 100°10′57.6″ E | 2016-08-06 | 2 | 17 | ||
| S7 | Manorom | 15°23′11.6″ N 100°08′29.6″ E | 2016-08-13 | 1 | 4 | ||
| S8 | Sapphaya | 15°08′15.5″ N 100°14′44.7″ E | 2016-09-17 | 2 | 13 | ||
| Total | 9 | 56 | |||||
| Nakhon Sawan | |||||||
| S9 | Phayuha Khiri | 15°28′06.3″ N 100°12′17.7″ E | 2016-09-16 | 28.1–28.6 | 374.7 | 4 | 19 |
| S10 | Phaisali | 15°38′20.6″ N 100°46′49.6″ E | 2016-09-16 | 2 | 16 | ||
| S11 | Tak Fa | 15°21′21.2″ N 100°30′10.0″ E | 2016-09-17 | 2 | 12 | ||
| S12 | Takhli | 15°16′19.5″ N 100°21′48.1″ E | 2016-09-17 | 1 | 5 | ||
| Total | 9 | 52 | |||||
| Phetchabun | |||||||
| S13 | Bueng Sam Phan | 15°43′51.0″ N 100°52′34.9″ E | 2016-09-16 | 27.2–27.8 | 290.5 | 2 | 10 |
| S14 | Mueang Phetchabun | 16°18′08.2″ N 101°03′58.2″ E | 2016-09-17 | 3 | 15 | ||
| S15 | Chon Daen | 16°12′32.6″ N 100°57′48.9″ E | 2016-09-17 | 4 | 21 | ||
| Total | 9 | 46 | |||||
| Taxa | Number of Strain | Total | FO (%) a | |||
|---|---|---|---|---|---|---|
| Suphan Buri | Chai Nat | Nakhon Sawan | Phetchabun | |||
| Phylum Ascomycota Subphylum Saccharomycotina | ||||||
| Candida intermedia | 1 | 1 | 3 | - | 5 | 13.9 |
| Candida quercitrusa | - | - | - | 1 | 1 | 2.8 |
| Candida saopaulonensis | - | 2 | - | 2 | 4 | 11.1 |
| Candida uthaithanina | - | 1 | - | - | 1 | 2.8 |
| Kodamaea ohmeri | 1 | - | - | - | 1 | 2.8 |
| Metschnikowia lopburiensis | - | 1 | 1 | 1 | 3 | 8.3 |
| Meyerozyma caribbica | 2 | 3 | 5 | 4 | 14 | 38.9 |
| Pichia bruneiensis | - | 2 | - | 4 | 6 | 16.7 |
| Pichia kudriavzevii | 1 | - | 1 | - | 2 | 5.6 |
| Wickerhamomyces anomalus | 2 | 1 | - | - | 3 | 8.3 |
| Phylum Basidiomycota Subphylum Agaricomycotina | ||||||
| Dioszegia zsoltii | 1 | 1 | 1 | - | 3 | 8.3 |
| Hannaella oryzae | - | - | 2 | - | 2 | 5.6 |
| Hannaella pagnoccae | - | 2 | - | 1 | 3 | 8.3 |
| Hannaella phetchabunensis | - | 1 | - | - | 1 | 2.8 |
| Hannaella phyllophila | - | 1 | 1 | 1 | 3 | 8.3 |
| Hannaella siamensis | 1 | 2 | - | 2 | 5 | 13.9 |
| Hannaella sinensis | 5 | 8 | 6 | 5 | 24 | 66.7 |
| Potential new species closest to Vishniacozyma heimaeyensis | 1 | 1 | - | - | 2 | 5.6 |
| Kwoniella bestiolae | 1 | - | - | - | 1 | 2.8 |
| Kwoniella heveanensis | - | 1 | - | - | 1 | 2.8 |
| Papiliotrema flavescens | 1 | 5 | 2 | 6 | 14 | 38.9 |
| Papiliotrema laurentii | 3 | 1 | 1 | - | 5 | 13.9 |
| Papiliotrema rajasthanensis | 5 | 2 | 3 | 1 | 11 | 30.6 |
| Papiliotrema ruineniae | - | 1 | 1 | - | 2 | 5.6 |
| Plapiliotrema plantarum | - | 1 | - | - | 1 | 2.8 |
| Saitozyma flava | 1 | - | - | 1 | 2 | 5.6 |
| Subphylum Pucciniomycotina | ||||||
| Cystobasidium calyptogenae | 1 | - | - | - | 1 | 2.8 |
| Symmetrospora vermiculata | 1 | 1 | 1 | - | 3 | 8.3 |
| Rhodotorula mucilaginosa | 2 | - | - | - | 2 | 5.6 |
| Rhodotorula paludigena | 5 | 4 | 5 | 4 | 18 | 50.0 |
| Rhodotorula toruloides | - | - | 1 | - | 1 | 2.8 |
| Potential new species closest to Rhodotorula toruloides | 1 | 1 | - | - | 2 | 5.6 |
| Rhodosporidiobolus nylandii | 1 | - | - | - | 1 | 2.8 |
| Rhodosporidiobolus ruineniae | - | - | 2 | 1 | 3 | 8.3 |
| Rhodosporidiobolus fluvialis | - | 1 | - | - | 1 | 2.8 |
| Rhodosporidiobolus odoratus | 1 | - | - | - | 1 | 2.8 |
| Sporobolomyces blumeae | 5 | - | 2 | - | 7 | 19.4 |
| Sporobolomyces nakasei | - | - | - | 6 | 6 | 16.7 |
| Sporobolomyces carnicolor | - | 1 | - | 1 | 2 | 5.6 |
| Subphylum Ustilaginomycotina | ||||||
| Anthracocystis elionuri | 1 | 3 | 2 | - | 6 | 16.7 |
| Dirkmeia churashimaensis | 8 | 4 | 4 | 1 | 17 | 47.2 |
| Moesziomyces antarcticus | 3 | - | 5 | 2 | 10 | 27.8 |
| Ustilago trichophora | 1 | 1 | - | - | 2 | 5.6 |
| Ustilago siamensis | 1 | - | - | - | 1 | 2.8 |
| Ustilago sparsa | 6 | 2 | 3 | 2 | 13 | 36.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Into, P.; Pontes, A.; Sampaio, J.P.; Limtong, S. Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand. Microorganisms 2020, 8, 80. https://doi.org/10.3390/microorganisms8010080
Into P, Pontes A, Sampaio JP, Limtong S. Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand. Microorganisms. 2020; 8(1):80. https://doi.org/10.3390/microorganisms8010080
Chicago/Turabian StyleInto, Parichat, Ana Pontes, José Paulo Sampaio, and Savitree Limtong. 2020. "Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand" Microorganisms 8, no. 1: 80. https://doi.org/10.3390/microorganisms8010080
APA StyleInto, P., Pontes, A., Sampaio, J. P., & Limtong, S. (2020). Yeast Diversity Associated with the Phylloplane of Corn Plants Cultivated in Thailand. Microorganisms, 8(1), 80. https://doi.org/10.3390/microorganisms8010080