Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation


Abstract
:1. Introduction
2. Materials and Methods
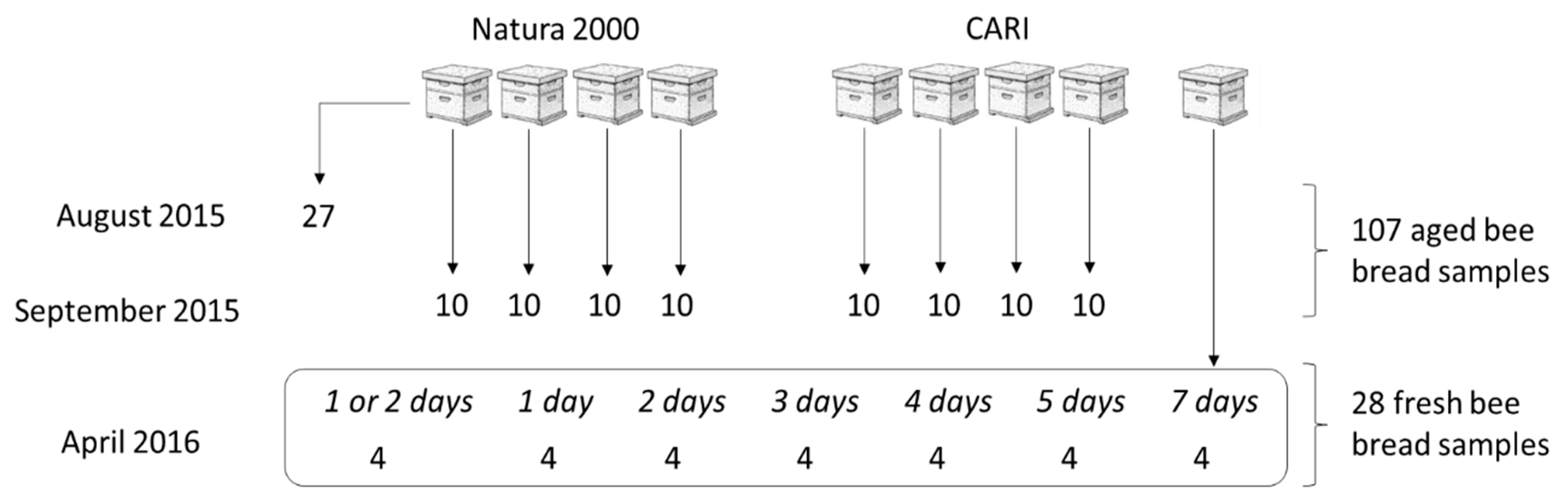
2.1. Samples
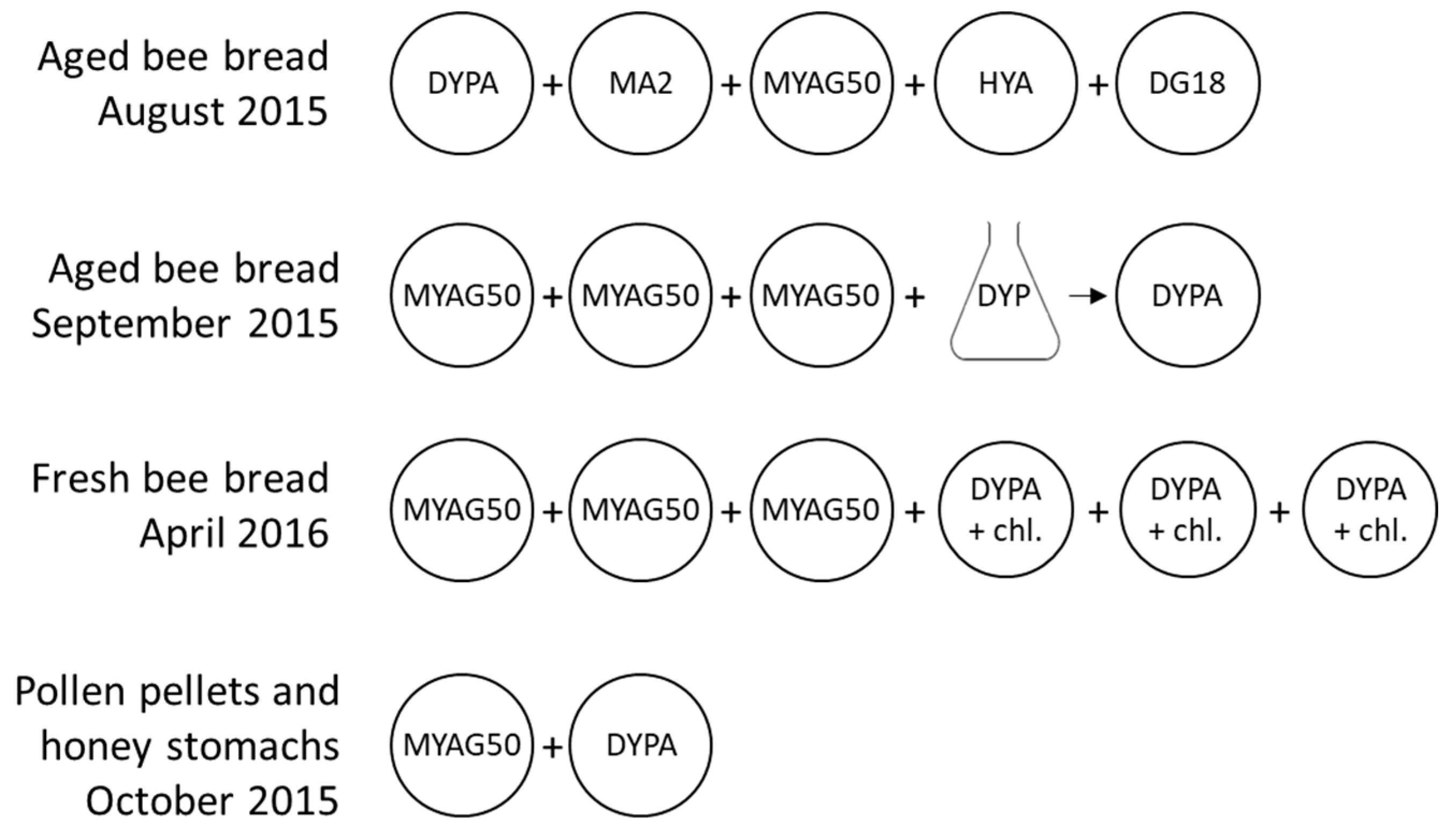
2.2. Cultures
2.3. Isolates and Identifications
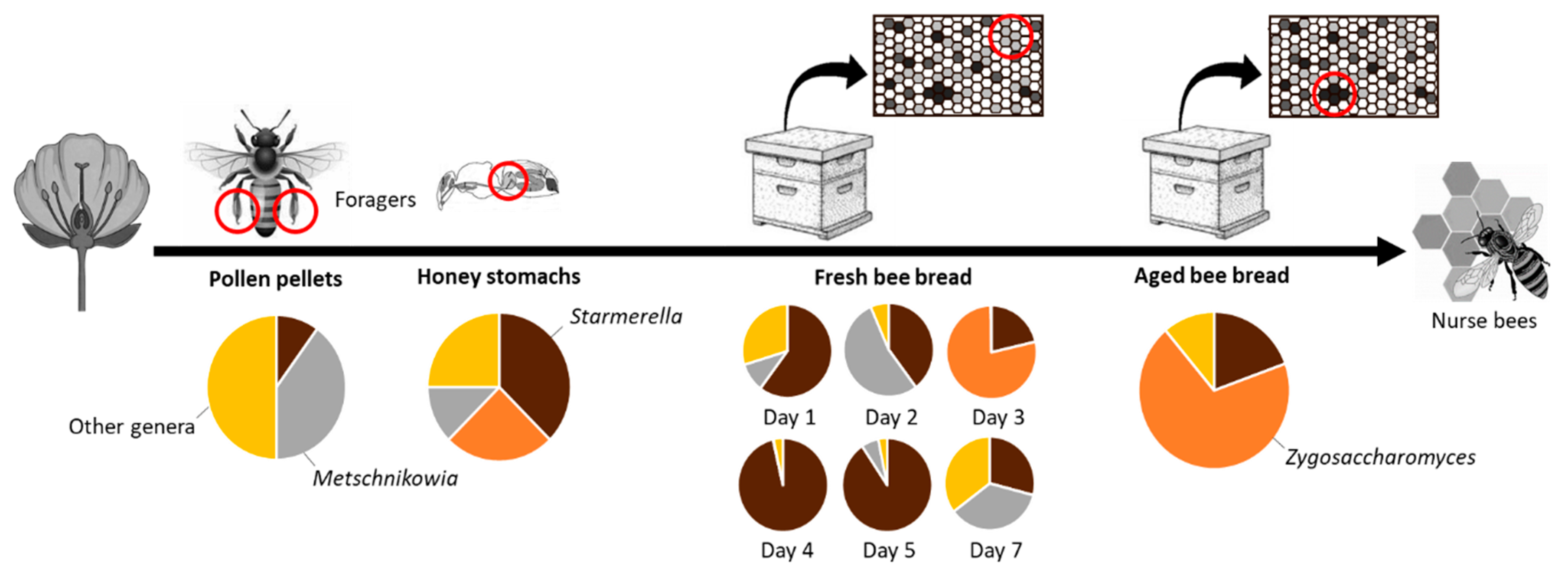
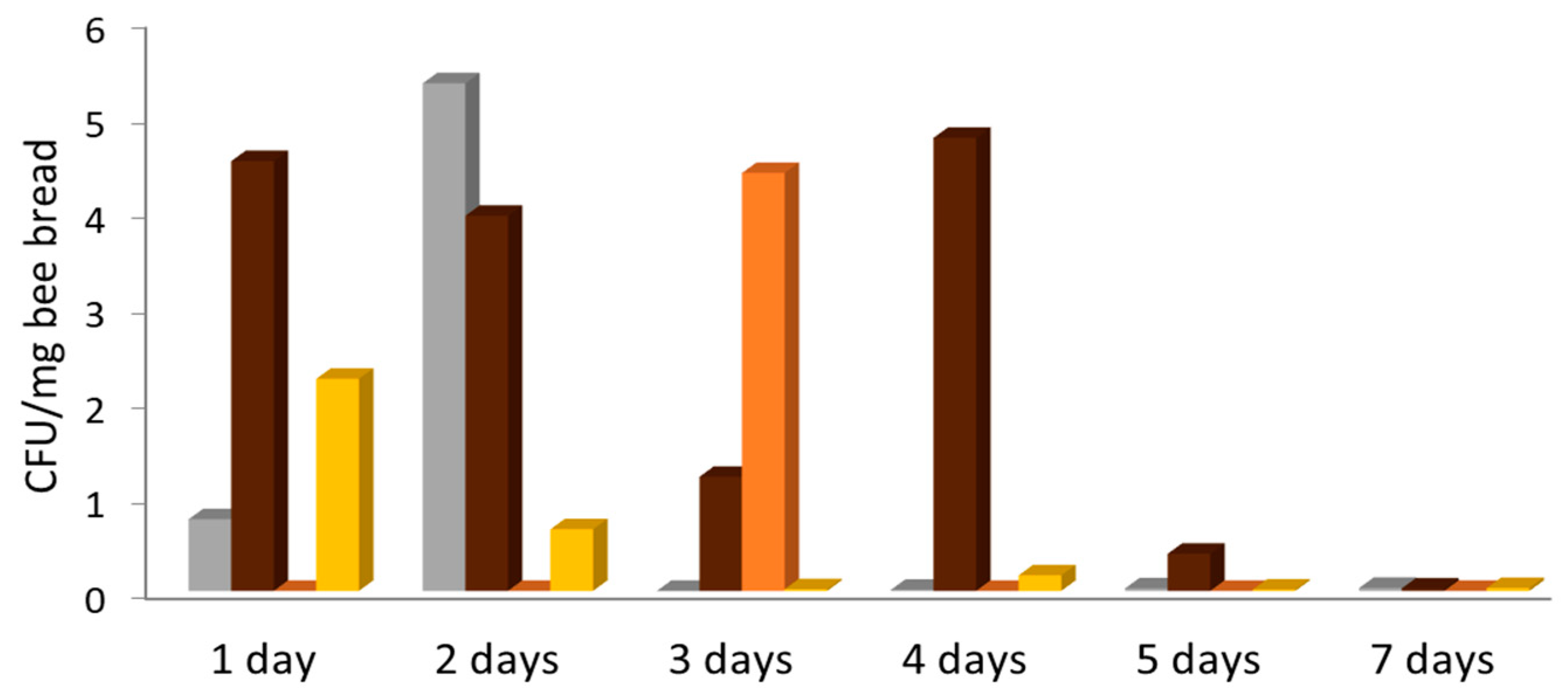
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Parker, R.L. The Collection and Utilization of Pollen by the Honeybee. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, September 1925. [Google Scholar]
- Podrižnik, B.; Božič, J. Maturation and stratification of antibacterial activity and total phenolic content of bee bread in honey comb cells. J. Apic. Res. 2015, 54, 81–92. [Google Scholar] [CrossRef]
- Vanengelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J.; Pettis, J.S. “Entombed Pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant. Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H.; Buchmann, S.L. What governs protein content of pollen: Pollinator preferences, pollen-pistil interactions, or phylogeny? Ecol. Monogr. 2000, 70, 617. [Google Scholar] [CrossRef]
- Pain, J.; Maugenet, J. Recherches biochimiques et physiologiques sur le pollen emmagasiné par les abeilles. Apidologie 1966, 9, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Herbert, E.W., Jr.; Shimanuki, H. Chemical composition and nutritive value of bee-collected and bee-stored pollen. Apidologie 1978, 9, 33–40. [Google Scholar] [CrossRef] [Green Version]
- DeGrandi-Hoffman, G.; Eckholm, B.J.; Huang, M.H. A comparison of bee bread made by Africanized and European honey bees (Apis mellifera) and its effects on hemolymph protein titers. Apidologie 2012, 44, 52–63. [Google Scholar] [CrossRef]
- Beutler, R.; Opfinger, E. Pollenernährung und nosemabefall der honigbiene (Apis Mellifica). J. Comp. Physiol. A 1950, 32, 383–421. [Google Scholar] [CrossRef]
- Cremonez, T.M.; De Jong, D.; Bitondi, M.M.G. Quantification of hemolyph proteins as a fast method for testing protein diets for honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1998, 91, 1284–1289. [Google Scholar] [CrossRef]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-stored pollen of honey bees: Many lines of evidence are consistent with pollen preservation, not nutrient conversion. Mol. Ecol. 2014, 23, 5904–5917. [Google Scholar] [CrossRef]
- Carroll, M.J.; Brown, N.; Goodall, C.; Downs, A.M.; Sheenan, T.H.; Anderson, K.E. Honey bees preferentially consume freshly-stored pollen. PLoS ONE 2017, 12, e0175933. [Google Scholar] [CrossRef] [Green Version]
- Maes, P.; Rodrigues, P.A.P.; Oliver, R.; Mott, B.M.; Anderson, K.E. Diet-related gut bacterial dysbiosis correlates with impaired development, increased mortality and Nosema disease in the honeybee (Apis mellifera). Mol. Ecol. 2016, 25, 5439–5450. [Google Scholar] [CrossRef]
- Vásquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- De Vuyst, L.; Lefeber, T.; Papalexandratou, Z.; Camu, N. The functional role of lactic acid bacteria in cocoa bean fermentation. In Biotechnology of Lactic Acid Bacteria; Wiley: Hoboken, NJ, USA, 2010; pp. 301–325. [Google Scholar]
- Chevtchik, V. Mikrobiologie pylového kvašení. Publ. Fac. Sci. Univ. Masaryk 1950, 323, 103–130. [Google Scholar]
- Gilliam, M. Microbiology of pollen and bee bread: The yeasts. Apidologie 1979, 10, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; Hansen, A.K.; Powell, J.E.; Sabree, Z.L. Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS ONE 2012, 7, e36393. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial ecology of the hive and pollination landscape: Bacterial associates from floral nectar, the alimentary tract and stored food of honey bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.A.; Zemolin, A.P.P.; Franco, J.L.; Boldo, J.T.; Stefenon, V.M.; Triplett, E.W.; Camargo, F.A.D.O.; Roesch, L.F.W. Relationship between honeybee nutrition and their microbial communities. Antonie van Leeuwenhoek 2015, 107, 921–933. [Google Scholar] [CrossRef]
- Khan, K.A.; Al-Ghamdi, A.A.; Ghramh, H.A.; Ansari, M.J.; Ali, H.; Alamri, S.A.; Kahtani, S.N.A.; Adgaba, N.; Qasim, M.; Hafeez, M. Structural diversity and functional variability of gut microbial communities associated with honey bees. Microb. Pathog. 2020, 138, 103793. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; Vanengelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.-H.; Jung, M.-J.; Kim, P.S.; Bae, J.-W. Social status shapes the bacterial and fungal gut communities of the honey bee. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pozo, I.M.; Van Kemenade, G.; Van Oystaeyen, A.; Aledón-Catalá, T.; Benavente, A.; Ende, W.V.D.; Wäckers, F.; Jacquemyn, H. The impact of yeast presence in nectar on bumble bee behavior and fitness. Ecol. Monogr. 2019, 90, 01393. [Google Scholar] [CrossRef]
- Carreck, N.L.; Andree, M.; Brent, C.S.; Cox-Foster, D.; Dade, A.H.; Ellis, J.D.; Hatjina, F.; Van Englesdorp, D. Standard methods forApis melliferaanatomy and dissection. J. Apic. Res. 2013, 52, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Hennebert, G.; Université catholique de Louvain, Louvain-la-Neuve, Belgium. Personal communication/Internal quality document I013N, 2019.
- Hocking, A.D.; Pitt, I.J. Dichloran-glycerol medium for enumeration of xerophilic fungi from low-moisture foods. Appl. Environ. Microbiol. 1980, 39, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Moncalvo, J.-M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Daniel, H. Evaluation of ribosomal RNA and actin gene sequences for the identification of ascomycetous yeasts. Int. J. Food Microbiol. 2003, 86, 61–78. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Lachance, M.-A. Metschnikowia: Half tetrads, a regicide and the fountain of youth. Yeast 2016, 33, 563–574. [Google Scholar] [CrossRef]
- Lachance, M.-A. Starmerella Rosa & Lachance (1998). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 2, pp. 811–815. [Google Scholar]
- Lachance, M.-A.; Dobson, J.; Wijayanayaka, D.N.; Smith, A.M.E. The use of parsimony network analysis for the formal delineation of phylogenetic species of yeasts: Candida apicola, Candida azyma, and Candida parazyma sp. nov., cosmopolitan yeasts associated with floricolous insects. Antonie van Leeuwenhoek 2010, 97, 155–170. [Google Scholar] [CrossRef]
- Haydak, M.H. Value of pollen substitutes for brood rearing of honeybees. J. Econ. Entomol. 1945, 38, 484–487. [Google Scholar] [CrossRef]
- Rosa, C.A.; Lachance, M.-A.; Silva, J.O.; Teixeira, A.C.P.; Marini, M.M.; Antonini, Y.; Martins, R.P. Yeast communities associated with stingless bees. FEMS Yeast Res. 2003, 4, 271–275. [Google Scholar] [CrossRef]
- Lachance, M.-A.; Starmer, W.T.; Rosa, C.A.; Bowles, J.M.; Barker, J.F.; Janzen, D.H. Biogeography of the yeasts of ephemeral flowers and their insects. FEMS Yeast Res. 2001, 1, 1–8. [Google Scholar] [CrossRef]
- Inglis, G.; Sigler, L.; Goette, M. Aerobic microorganisms associated with alfalfa leafcutter bees (Megachile rotundata). Microb. Ecol. 1993, 26, 125–143. [Google Scholar] [CrossRef]
- Spencer, J.F.T.; Gorin, P.A.J.; Hobbs, G.A.; Cooke, D.A. Yeasts isolated from bumblebee honey from Western Canada: Identification with the aid of proton magnetic resonance spectra of their mannose-containing polysaccharides. Can. J. Microbiol. 1970, 16, 117–119. [Google Scholar] [CrossRef]
- Egorova, A.I.; Bab’eva, I.P. Yeast flora of the honey bee (Apis mellifica L.). Izvestija Sibirskogo otdelenija Akademii nauk SSSR. Ser. Biol. Med. Nauk 1967, 2, 127–132. [Google Scholar]
- Teixeira, A.C.P.; Marini, M.M.; Nicoli, J.R.; Antonini, Y.; Martins, R.P.; Lachance, M.-A.; Rosa, C.A. Starmerella meliponinorum sp. nov., a novel ascomycetous yeast species associated with stingless bees. Int. J. Syst. Evol. Microbiol. 2003, 53, 339–343. [Google Scholar] [CrossRef]
- Pimentel, M.R.; Antonini, Y.; Martins, R.P.; Lachance, M.-A.; Rosa, C.A. Candida riodocensis and Candida cellae, two new yeast species from the clade associated with solitary bees in the Atlantic rain forest of Brazil. FEMS Yeast Res. 2005, 5, 875–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, H.-M.; Rosa, C.A.; Thiago-Calaça, P.S.S.; Antonini, Y.; Bastos, E.M.A.F.; Evrard, P.; Huret, S.; Fidalgo-Jiménez, A.; Lachance, M.-A. Starmerella neotropicalis f. a., sp. nov., a yeast species found in bees and pollen. Int. J. Syst. Evol. Microbiol. 2013, 63, 3896–3903. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.A.; Viana, E.M.; Martins, R.P.; Antonini, Y.; Lachance, M.-A. Candida batistae, a new yeast species associated with solitary digger nesting bees in Brazil. Mycologia 1999, 91, 428. [Google Scholar] [CrossRef]
- Monrocher, R. Quelques nouvelles espèces et variétés du genre Candida (levures asporogènes). Rev. Mycol. 1967, 32, 69–92. [Google Scholar]
- Brysch-Herzberg, M. Ecology of yeasts in plant–bumblebee mutualism in Central Europe. FEMS Microbiol. Ecol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Dharampal, P.S.; Diaz-Garcia, L.; Haase, M.A.B.; Zalapa, J.E.; Currie, C.R.; Hittinger, C.T.; Steffan, S.A. Microbial diversity associated with the pollen stores of captive-bred bumble bee colonies. Insects 2020, 11, 250. [Google Scholar] [CrossRef]
- Grilione, P.; Federici, F.; Miller, M. Yeasts from honey bees (Apis Mellifera L.). In Advances in Biotechnology, Proceedings of the Fifth International Yeast Symposium, London, Canada, July 20–25 1980; Elsevier BV: Amsterdam, The Netherlands, 1981; pp. 599–605. [Google Scholar] [CrossRef]
- Lavie, P. Essais de lutte biologique contre l’Acarapis woodi, agent de l’acariose de l’abeille. C. R. Hebd. Acad. Sci. 1954, 238, 947–948. [Google Scholar]
- Neto, D.J.D.C.; De Morais, P.B. The vectoring of Starmerella species and other yeasts by stingless bees in a Neotropical savanna. Fungal Ecol. 2020, 47, 100973. [Google Scholar] [CrossRef]
- Parle, J.N.; Di Menna, M.E. The source of yeasts in New Zealand wines. N. Z. J. Agric. Res. 1966, 9, 98–107. [Google Scholar] [CrossRef]
- Rosa, C.A.; Lachance, M.-A. The yeast genus Starmerella gen. nov. and Starmerella bombicola sp. nov., the teleomorph of Candida bombicola (Spencer, Gorin & Tullock) Meyer & Yarrow. Int. J. Syst. Bacteriol. 1998, 48, 1413–1417. [Google Scholar] [CrossRef]
- Jones, D.F. Novel macrocyclic glycolipids from Torulopsis gropengiesseri. J. Chem. Soc. C 1967, 6, 479–484. [Google Scholar] [CrossRef]
- Konishi, M.; Fukuoka, T.; Morita, T.; Imura, T.; Kitamoto, D. Production of new types of sophorolipids by Candida batistae. J. Oleo Sci. 2008, 57, 359–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzman, C.P.; Price, N.P.; Ray, K.J.; Kuo, T.-M. Production of sophorolipid biosurfactants by multiple species of the Starmerella (Candida) bombicola yeast clade. FEMS Microbiol. Lett. 2010, 311, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Morita, T.; Fukuoka, T.; Imura, T.; Uemura, S.; Iwabuchi, H.; Kitamoto, D. efficient production of acid-form sophorolipids from waste glycerol and fatty acid methyl esters by Candida floricola. J. Oleo Sci. 2018, 67, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Hommel, R.; Weber, L.; Weiss, A.; Himmelreich, U.; Rilke, O.; Kleber, H.-P. Production of sophorose lipid by Candida (Torulopsis) apicola grown on glucose. J. Biotechnol. 1994, 33, 147–155. [Google Scholar] [CrossRef]
- Van Bogaert, I.N.A.; Saerens, K.; De Muynck, C.; Develter, D.; Soetaert, W.; Vandamme, E.J. Microbial production and application of sophorolipids. Appl. Microbiol. Biotechnol. 2007, 76, 23–34. [Google Scholar] [CrossRef]
- Marchant, R.; Banat, I.M. Microbial biosurfactants: Challenges and opportunities for future exploitation. Trends Biotechnol. 2012, 30, 558–565. [Google Scholar] [CrossRef]
- Pacini, E.; Hesse, M. Pollenkitt—its composition, forms and functions. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 399–415. [Google Scholar] [CrossRef]
- James, S.A.; Stratford, M. Zygosaccharomyces Barker (1901). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 2, pp. 937–947. [Google Scholar]
- Solieri, L.; Dakal, T.C.; Giudici, P. Zygosaccharomyces sapae sp. nov., isolated from Italian traditional balsamic vinegar. Int. J. Syst. Evol. Microbiol. 2013, 63, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Sinpoo, C.; Williams, G.R.; Chantawannakul, P. Dynamics of fungal communities in corbicular pollen and bee bread. Chiang Mai J. Sci. 2017, 44, 1244–1256. [Google Scholar]
- Čadež, N.; Fülöp, L.; Dlauchy, D.; Péter, G. Zygosaccharomyces favi sp. nov., an obligate osmophilic yeast species from bee bread and honey. Antonie van Leeuwenhoek 2014, 107, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Péter, G.; National Collection of Agricultural and Industrial Microorganisms (NCAIM), Szent István University, Budapest, Hungary. Personal communication, 2017.
- James, S.A.; Stratford, M. Spoilage yeasts with emphasis on the genus Zygosaccharomyces. In Yeasts in Food; Elsevier BV: Amsterdam, The Netherlands, 2003; pp. 171–191. [Google Scholar]
- Grüẞ, J. Die Anpassung eines Pilzes (Anthomyces Reukaufii) an den Blütenbau und den Bienenrüssel. Ber. Deut. Bot. Ges. 1918, 35, 746–762. [Google Scholar]
- Pozo, M.I.; Lachance, M.-A.; Herrera, C.M. Nectar yeasts of two southern Spanish plants: The roles of immigration and physiological traits in community assembly. FEMS Microbiol. Ecol. 2012, 80, 281–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, B.; Lachance, M.-A.; Herrera, C.M. Phylogenetic analysis of the angiosperm-floricolous insect–yeast association: Have yeast and angiosperm lineages co-diversified? Mol. Phylogenetics Evol. 2013, 68, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey bee gut microbiome is altered by in-hive pesticide exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Dapporto, L.; Legras, J.-L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [Green Version]
- Hajsig, M. Contribution to the knowledge of yeasts from the intestinal content of bees. Vet. Arhiv 1959, 29, 145–156. [Google Scholar]
- Sandhu, D.K.; Waraich, M.K. Yeasts associated with pollinating bees and flower nectar. Microb. Ecol. 1985, 11, 51–58. [Google Scholar] [CrossRef]
- Batra, L.R.; Batra, S.W.T.; Bohart, G.E. The mycoflora of domesticated and wild bees (Apoidea). Mycopathol. 1973, 49, 13–44. [Google Scholar] [CrossRef]
- Burnside, C.E. Fungous Disease of the Honeybee; Technical Report for United States Department of Agriculture: Washington, DC, USA, 1930; Volume 149, pp. 1–41. [Google Scholar]
- Gilliam, M.; Wickerham, L.; Morton, H.L.; Martin, R.D. Yeasts isolated from honey bees, Apis mellifera, fed 2,4-D and antibiotics. J. Invertebr. Pathol. 1974, 24, 349–356. [Google Scholar] [CrossRef]
- Gilliam, M.; Morton, H.L.; Prest, D.B.; Martin, R.D.; Wickerham, L. The mycoflora of adult worker honeybees, Apis mellifera: Effects of 2,4,5-T and caging of bee colonies. J. Invertebr. Pathol. 1977, 30, 50–54. [Google Scholar] [CrossRef]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Paleolog, J. Impact of nosemosis on the intestinal yeast flora of honey bees. Med. Weter. 2013, 69, 726–729. [Google Scholar]
- Torto, B.; Boucias, I.G.; Arbogast, R.T.; Tumlinson, J.H.; Teal, P.E.A. Multitrophic interaction facilitates parasite-host relationship between an invasive beetle and the honey bee. Proc. Natl. Acad. Sci. USA 2007, 104, 8374–8378. [Google Scholar] [CrossRef] [Green Version]
- Benda, N.D. Detection and characterization of kodamaea ohmeri associated with small hive beetle Aethina tumida infesting honey bee hives. J. Apic. Res. 2008, 47, 194–201. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Composition | Application |
|---|---|---|
| DYPA | Glucose 2% (Rocc, Sart-Eustache, Belgium) | General |
| Dextrose Yeast Extract Peptone Agar | Peptone 1% (Duchefa, Haarlem, Netherlands) | |
| Yeast extract 0.5% (Oxoid, Basingstoke, United Kingdom) | ||
| Agar 2% (Rocc, Sart-Eustache, Belgium) | ||
| MA2 | Malt extract 2% (Duchefa, Haarlem, Netherlands) | General, rich in complex nutrients |
| Malt Agar 2% | Agar 1.5% | |
| MYAG50 | Glucose 50% | Osmophiles |
| Malt Yeast Agar Glucose 50% | Peptone 0.5% | |
| Yeast extract 0.3% | ||
| Malt extract 0.3% | ||
| Agar 2% | ||
| HYA | Honey without preservatives 1 3% | Basidiomycetous bee yeasts |
| Honey Yeast Agar [28] | Yeast extract 0.5% | |
| Agar 2% | ||
| DG18 | Glucose 1% | Xerophilic fungi and yeasts |
| Dichloran-Glycerol Agar [29] | Peptone 0.5% | |
| Dipotassium phosphate 0.1% (VWR, Radnor, USA) | ||
| Magnesium sulfate 0.05% (VWR, Radnor, USA) | ||
| Chloramphenicol 0.01% (AppliChem, Darmstadt, Germany) | ||
| Glycerol 22% (VWR, Radnor, USA) | ||
| Agar 1.5% |
| Fresh Bee Bread 1 n = 28 | Aged Bee Bread 2 n = 107 | Pollen Pellets n = 12 | Honey Stomach n = 14 | |||||
|---|---|---|---|---|---|---|---|---|
| Samples | Colonies 3 | Samples | Colonies 3 | Samples | Colonies | Samples | Colonies | |
| Metschnikowia | ||||||||
| Metschnikowia (Candida) rancensis | 3 | 100 | 1 | 50 | ||||
| Metschnikowia cf. pulcherrima | 9 | 74.4 | 1 | 10 | ||||
| Starmerella | ||||||||
| Starmerella (Candida) apicola | 10 | 36.5 | 1 | 0.7 | ||||
| Starmerella (Candida) apis | 18 | 229.8 | ||||||
| Starmerella (Candida) bombi | 2 | 4.7 | 2 | 2 | 1 | 1 | ||
| Starmerella (Candida) magnoliae | 10 | 62.4 | 1 | 2 | 1 | 2 | 2 | 5 |
| Starmerella bombicola | 5 | 26.5 | 3 | >200 | ||||
| Zygosaccharomyces | ||||||||
| Zygosaccharomyces favi | 6 | 3.3 | ||||||
| Zygosaccharomyces mellis | 2 | 40.3 | 3 | 121 | ||||
| Zygosaccharomyces rouxii | 2 | 46 | 6 | 24.3 | 1 | 1 | ||
| Zygosaccharomyces sapae | 5 | 102 | ||||||
| Zygosaccharomyces siamensis | 1 | 0.3 | 5 | 14.3 | 1 | 2 | ||
| Other | ||||||||
| Aureobasidium pullulans | 1 | 0.7 | ||||||
| Candida parapsilosis | 1 4 | |||||||
| Debaryomyces hansenii | 9 | 5 | ||||||
| Debaryomyces sp. | 2 | 0.7 | ||||||
| Debaryomyces maramus | 2 | 4 | ||||||
| Dothiora prunorum | 2 | 0.6 | ||||||
| Hanseniaspora uvarum | 2 | 4 | 1 | 5 | ||||
| Kluyveromyces dobzhanskii | 1 | 3 | 1 | 3 | ||||
| Kodamaea ohmeri | 8 | 35.4 | ||||||
| Meyerozyma guilliermondii | 1 | 0.7 | ||||||
| Naganishia (Cryptococcus) diffluens | 1 | 1 | ||||||
| Rhodotorula mucilaginosa | 2 4 | |||||||
| Saccharomyces cerevisiae | 1 | >200 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detry, R.; Simon-Delso, N.; Bruneau, E.; Daniel, H.-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms 2020, 8, 1789. https://doi.org/10.3390/microorganisms8111789
Detry R, Simon-Delso N, Bruneau E, Daniel H-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms. 2020; 8(11):1789. https://doi.org/10.3390/microorganisms8111789
Chicago/Turabian StyleDetry, Roxane, Noa Simon-Delso, Etienne Bruneau, and Heide-Marie Daniel. 2020. "Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation" Microorganisms 8, no. 11: 1789. https://doi.org/10.3390/microorganisms8111789