Composition and Predominance of Fusarium Species Causing Fusarium Head Blight in Winter Wheat Grain Depending on Cultivar Susceptibility and Meteorological Factors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Area Surveyed and Survey Strategy
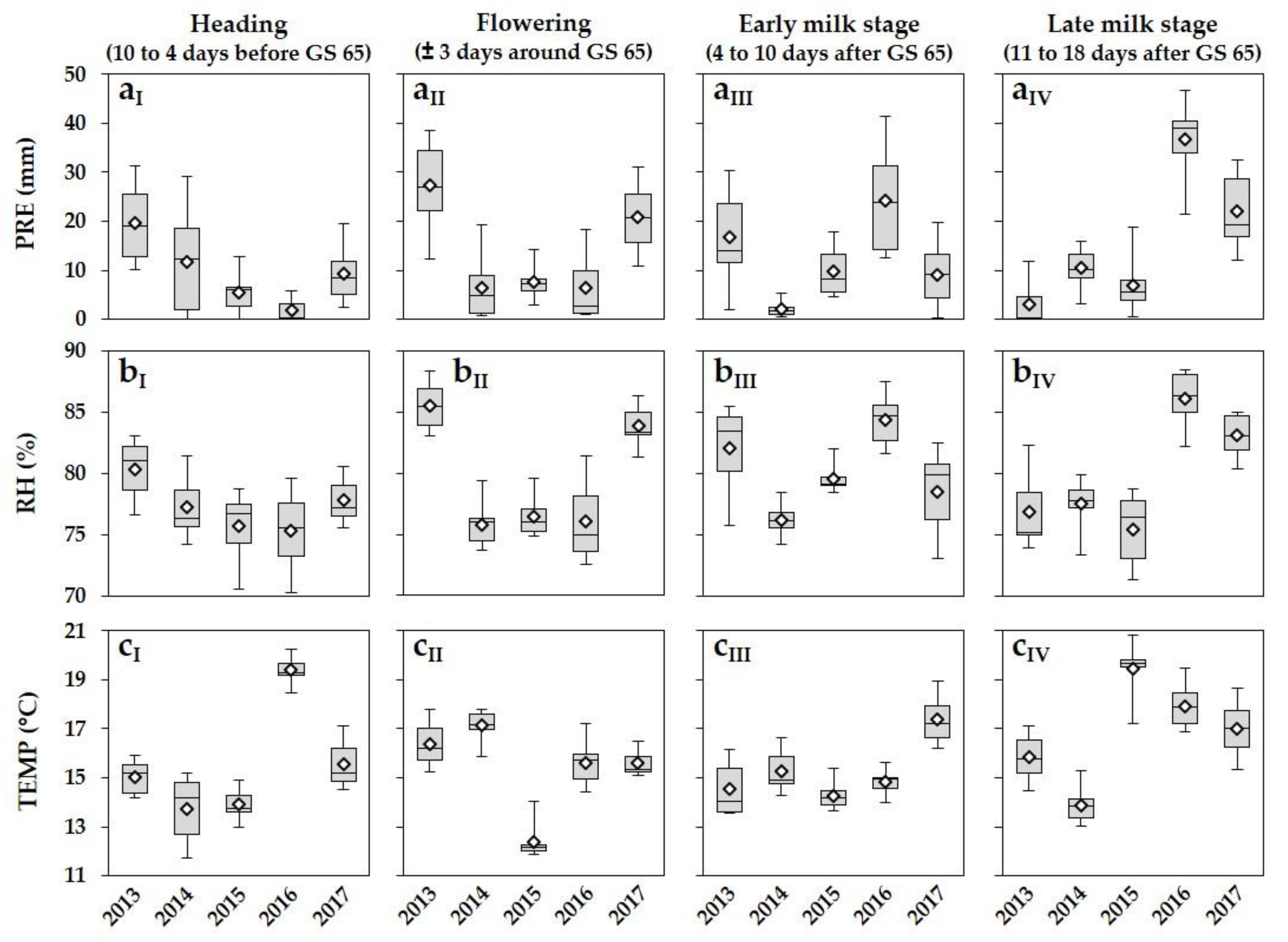
2.2. Meteorological Data
2.3. Sample Preparation
2.4. Fungal Isolates
2.5. DNA Extraction
2.6. Quantitative PCR (qPCR)
2.7. Analysis of Mycotoxins
2.8. Statistical Analysis
3. Results
3.1. Meteorological Conditions
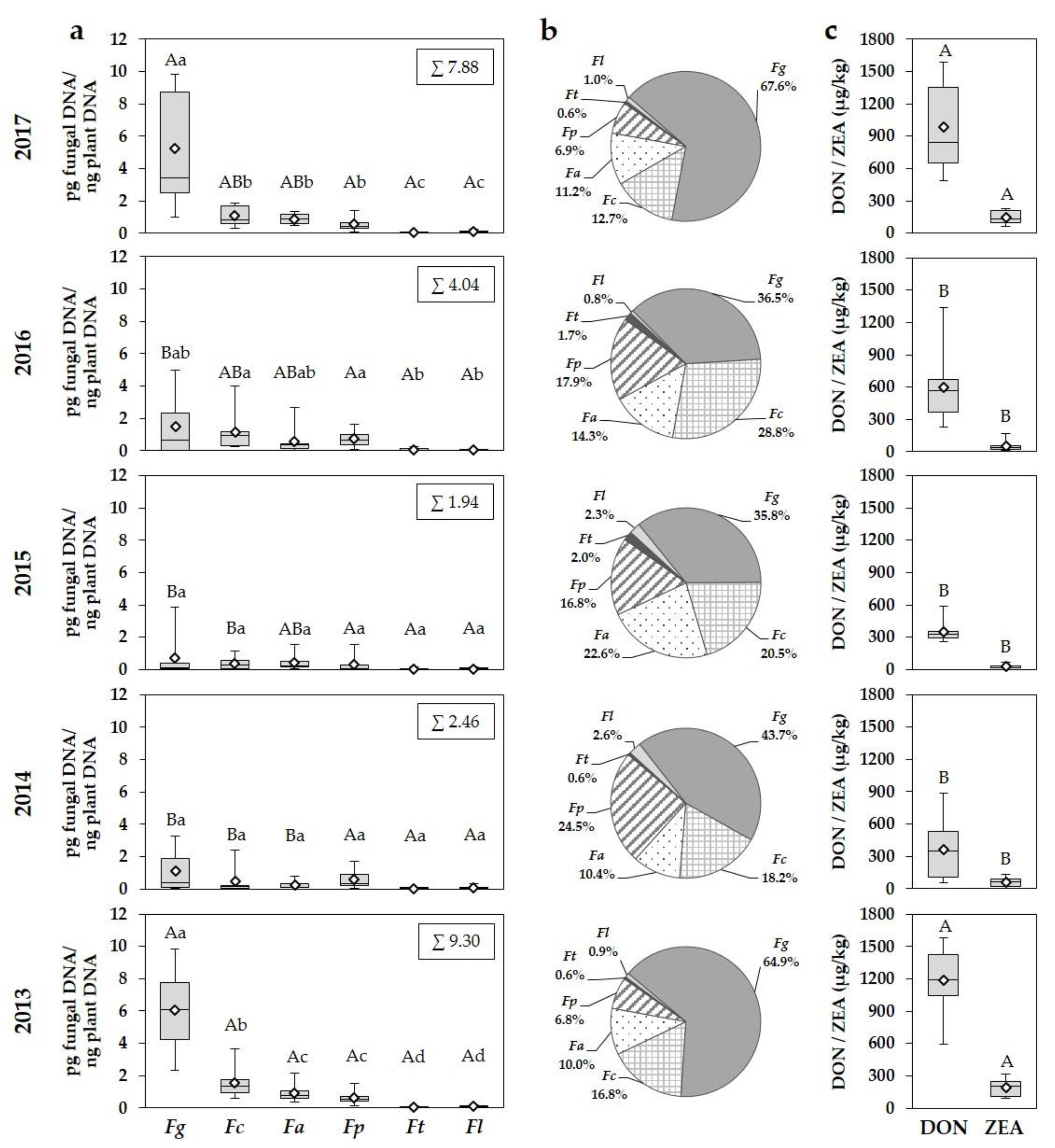
3.2. Fusarium Species in Wheat Grain
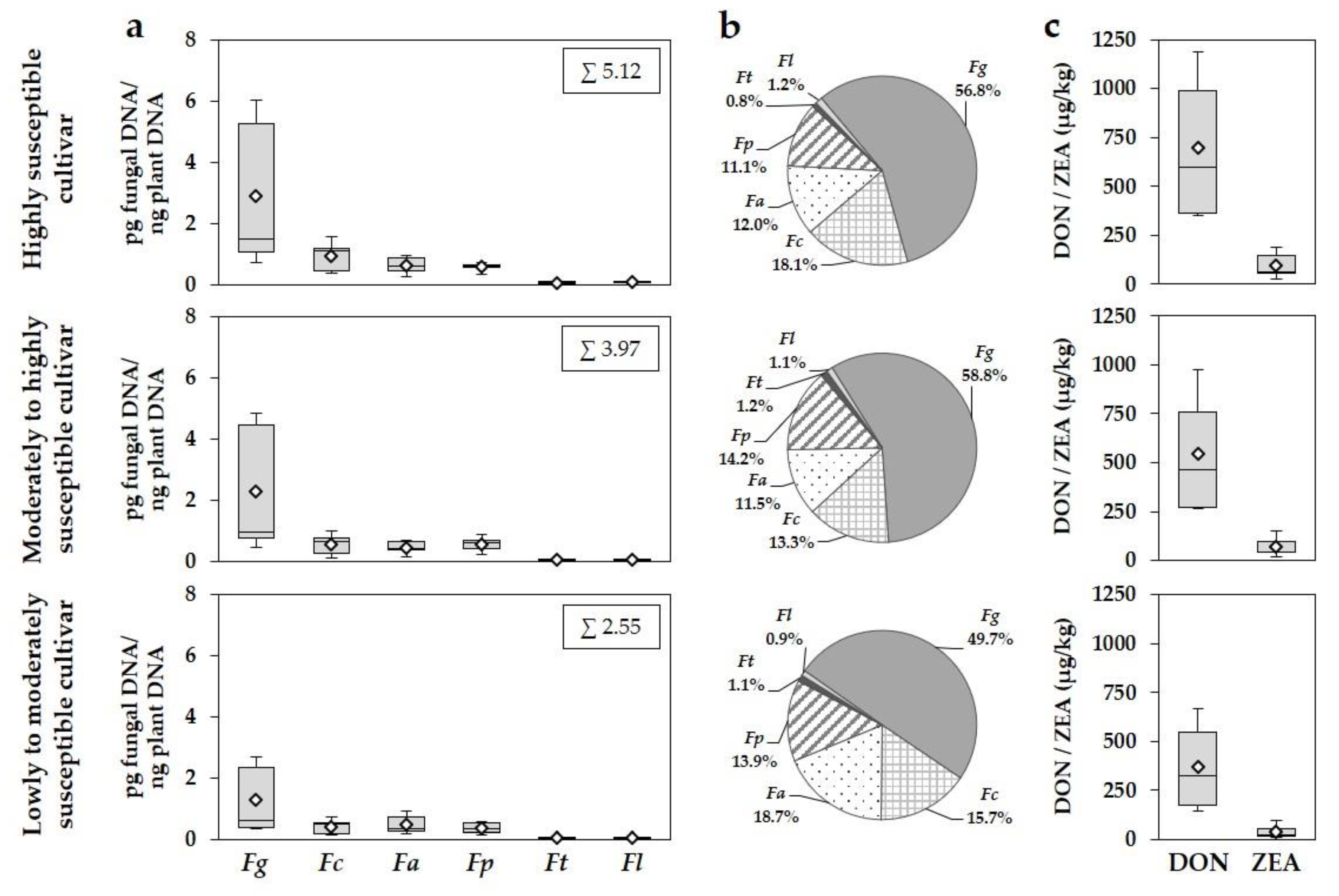
3.3. Effect of Year and Cultivar Susceptibility on Fusarium DNA Amounts, DON and ZEA Concentrations
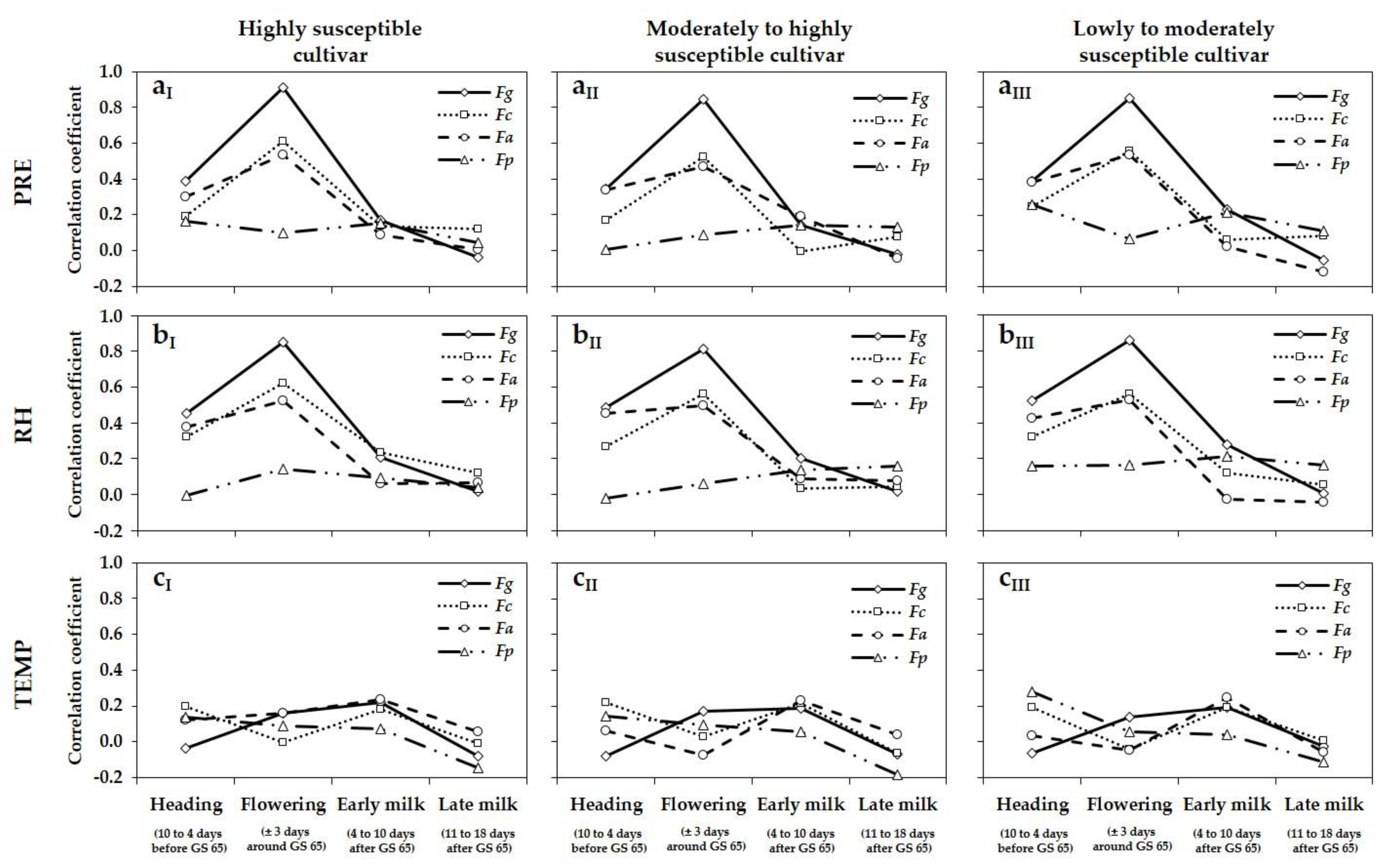
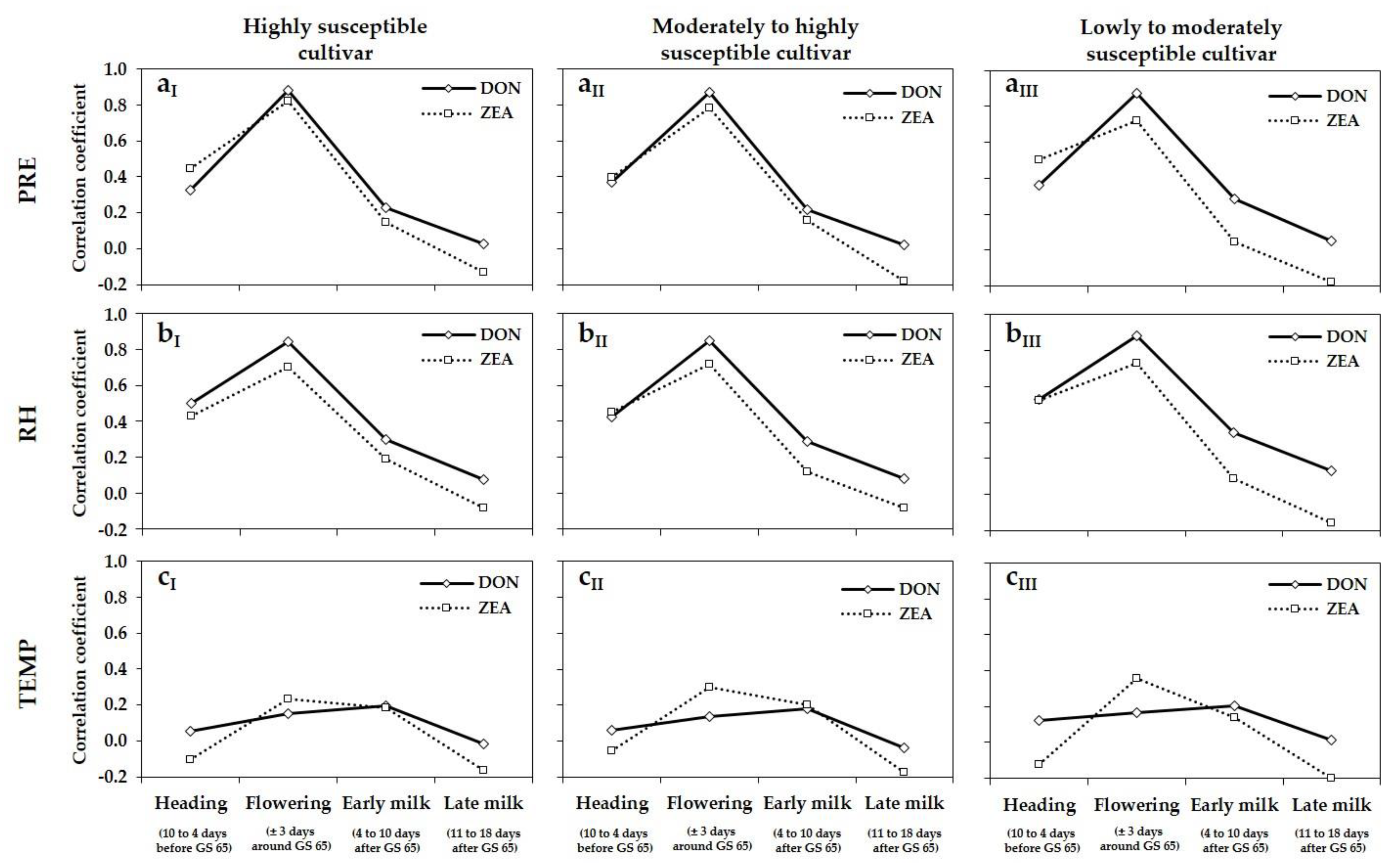
3.4. Relationship between Meteorological Variables, DNA Amounts, and DON and ZEA Concentrations
3.5. Relationship between DNA Amounts and DON and ZEA Concentrations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals-a review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Logrieco, A.; Mulè, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G. Influence of agricultural practices on fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett. 2004, 153, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef]
- Martínez, M.; Ramírez Albuquerque, L.; Arata, A.F.; Biganzoli, F.; Fernández Pinto, V.; Stenglein, S.A. Effects of Fusarium graminearum and Fusarium poae on disease parameters, grain quality and mycotoxins contamination in bread wheat (Part I). J. Sci. Food Agric. 2020, 100, 863–873. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics and Biology; APS Press (The American Phytopathological Society): St. Paul, MN, USA, 2006. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1126/2007of 28 September 2007 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants infoodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union 2007, L255, 14–17. [Google Scholar]
- Xu, X.-M.; Parry, D.W.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Edwards, S.G.; Cooke, B.M.; Doohan, F.M.; Brennan, J.M.; Moretti, A.; et al. Predominance and association of pathogenic fungi causing Fusarium ear blight in wheat in four European countries. Eur. J. Plant Pathol. 2005, 112, 143–154. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Czaban, J.; Wróblewska, B.; Sułek, A.; Mikos, M.; Boguszewska, E.; Podolska, G.; Nieróbca, A. Colonisation of winter wheat grain by Fusarium spp. and mycotoxin content as dependent on a wheat variety, crop rotation, a crop management system and weather conditions. Food Addit. Contam. Part A 2015, 32, 874–910. [Google Scholar] [CrossRef] [PubMed]
- Waalwijk, C.; Kastelein, P.; Vries, I.d.; Kerényi, Z.; van der Lee, T.; Hesselink, T.; Köhl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Jenkinson, P.; Parry, D.W. Splash dispersal of conidia of Fusarium culmorum and Fusarium avenaceum. Mycol. Res. 1994, 98, 506–510. [Google Scholar] [CrossRef]
- Maldonado-Ramirez, S.L.; Schmale, D.G.; Shields, E.J.; Bergstrom, G.C. The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of Fusarium head blight. Agric. For. Meteorol. 2005, 132, 20–27. [Google Scholar] [CrossRef]
- Keller, M.D.; Bergstrom, G.C.; Shields, E.J. The aerobiology of Fusarium graminearum. Aerobiologia 2014, 30, 123–136. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Xu, X.-M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Brennan, J.; Monaghan, S.; Moretti, A.; Mule, G.; et al. Relationship between the fungal complex causing Fusarium head blight of wheat and environmental conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef]
- Hooker, D.C.; Schaafsma, A.W.; Tamburic-Ilincic, L. Using weather variables pre- and post-heading to predict deoxynivalenol content in winter wheat. Plant Dis. 2002, 86, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Franz, E.; Booij, K.; van der Fels-Klerx, I. Prediction of deoxynivalenol content in dutch winter wheat. J. Food Prot. 2009, 72, 2170–2177. [Google Scholar] [CrossRef] [PubMed]
- Váňová, M.; Klem, K.; Matušinský, P.; Trnka, M. Prediction model for deoxynivalenol in wheat grain based on weather conditions. Plant Protect. Sci. 2010, 45, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Kriss, A.B.; Paul, P.A.; Xu, X.; Nicholson, P.; Doohan, F.M.; Hornok, L.; Rietini, A.; Edwards, S.G.; Madden, L.V. Quantification of the relationship between the environment and Fusarium head blight, Fusarium pathogen density, and mycotoxins in winter wheat in Europe. Eur. J. Plant Pathol. 2012, 133, 975–993. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Schaafsma, A.W.; Tamburic-Ilincic, L.; Hooker, D.C. Effect of previous crop, tillage, field size, adjacent crop, and sampling direction on airborne propagules of Gibberella zeae/Fusarium graminearum, fusarium head blight severity, and deoxynivalenol accumulation in winter wheat. Can. J. Plant Pathol. 2005, 27, 217–224. [Google Scholar] [CrossRef]
- Beyer, M.; Klix, M.B.; Klink, H.; Verreet, J.-A. Quantifying the effects of previous crop, tillage, cultivar and triazole fungicides on the deoxynivalenol content of wheat grain—A review. J. Plant Dis. Prot. 2006, 113, 241–246. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Bartók, T.; Kászonyi, G.; Varga, M.; Tóth, B.; Varga, J. Common resistance to different Fusarium spp. causing Fusarium head blight in wheat. Eur. J. Plant Pathol. 2005, 112, 267–281. [Google Scholar] [CrossRef]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Rossi, V.; Terzi, V.; Moggi, F.; Morcia, C.; Faccioli, P.; Haidukowski, M.; Pascale, M. Assessment of Fusarium infection in wheat heads using a quantitative polymerase chain reaction (qPCR) assay. Food Addit. Contam. 2007, 24, 1121–1130. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- Deutscher Wetterdienst (DWD). Klimareport Schleswig-Holstein. Available online: https://www.dwd.de/DE/leistungen/klimareport_sh/download_report_2017.pdf?__blob=publicationFile&v=5 (accessed on 10 March 2020).
- Statistisches Amt für Hamburg und Schleswig-Holstein. Die Bodennutzung in Schleswig-Holstein 2017. Available online: http://epub.sub.uni-hamburg.de/epub/volltexte/2018/78253/pdf/C_I_1_j_17_SH_e.pdf (accessed on 10 March 2020).
- Verreet, J.A.; Klink, H.; Hoffmann, G.M. Regional monitoring for disease prediction and optimization of plant protection measuares: The IPM wheat model. Plant Dis. 2000, 84, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birr, T.; Verreet, J.-A.; Klink, H. Prediction of deoxynivalenol and zearalenone in winter wheat grain in a maize-free crop rotation based on cultivar susceptibility and meteorological factors. J. Plant Dis. Prot. 2019, 126, 13–27. [Google Scholar] [CrossRef]
- Bundessortenamt. Beschreibende Sortenliste. Available online: https://www.bundessortenamt.de/bsa/media/Files/BSL/bsl_getreide_2008.pdf (accessed on 10 March 2020).
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Multcomp: Simultaneous Inference in General Parametric Models. R Package Version 1.4-10. Available online: https://cran.r-project.org/src/contrib/Archive/multcomp/ (accessed on 3 June 2019).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-140. Available online: https://cran.r-project.org/src/contrib/Archive/nlme/ (accessed on 3 June 2019).
- Laird, N.M.; Ware, J.H. Random-effects models for longitudinal data. Biometrics 1982, 38, 963–974. [Google Scholar] [CrossRef]
- Verbeke, G.; Molenberghs, G. Linear Mixed Models for Longitudinal Data; Springer: New York, NY, USA, 2000. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Schaarschmidt, F.; Vaas, L. Analysis of trials with complex treatment structure using multiple contrast tests. HortScience 2009, 44, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Bretz, F.; Hothorn, T.; Westfall, P.H. Multiple Comparisons Using R; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Lacey, J.; Bateman, G.L.; Mirocha, C.J. Effects of infection time and moisture on development of ear blight and deoxynivalenol production by Fusarium spp. in wheat. Ann. Appl. Biol. 1999, 134, 277–283. [Google Scholar] [CrossRef]
- Del Ponte, E.M.; Fernandes, J.M.C.; Bergstrom, G.C. Influence of growth stage on Fusarium head blight and deoxynivalenol production in wheat. J. Phytopathol. 2007, 155, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Nakajima, T. Deoxynivalenol and nivalenol accumulation in wheat infected with Fusarium graminearum during grain development. Phytopathology 2010, 100, 763–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isebaert, S.; de Saeger, S.; Devreese, R.; Verhoeven, R.; Maene, P.; Heremans, B.; Haesaert, G. Mycotoxin-producing Fusarium species occurring in winter wheat in Belgium (Flanders) during 2002–2005. J. Phytopathol. 2009, 157, 108–116. [Google Scholar] [CrossRef]
- Chandelier, A.; Nimal, C.; André, F.; Planchon, V.; Oger, R. Fusarium species and DON contamination associated with head blight in winter wheat over a 7-year period (2003–2009) in Belgium. Eur. J. Plant Pathol. 2011, 130, 403–414. [Google Scholar] [CrossRef]
- Klix, M.B.; Beyer, M.; Verreet, J.-A. Effects of cultivar, agronomic practices, geographic location, and meteorological conditions on the composition of selected Fusarium species on wheat heads. Can. J. Plant Pathol. 2008, 30, 46–57. [Google Scholar] [CrossRef]
- Audenaert, K.; van Broeck, R.; Bekaert, B.; de Witte, F.; Heremans, B.; Messens, K.; Höfte, M.; Haesaert, G. Fusarium head blight (FHB) in Flanders: Population diversity, inter-species associations and DON contamination in commercial winter wheat varieties. Eur. J. Plant Pathol. 2009, 125, 445–458. [Google Scholar] [CrossRef]
- Giraud, F.; Pasquali, M.; El Jarroudi, M.; Vrancken, C.; Brochot, C.; Cocco, E.; Hoffmann, L.; Delfosse, P.; Bohn, T. Fusarium head blight and associated mycotoxin occurrence on winter wheat in Luxembourg in 2007/2008. Food Addit. Contam. Part A 2010, 27, 825–835. [Google Scholar] [CrossRef]
- Talas, F.; Parzies, H.K.; Miedaner, T. Diversity in genetic structure and chemotype composition of Fusarium graminearum sensu stricto populations causing wheat head blight in individual fields in Germany. Eur. J. Plant Pathol. 2011, 131, 39–48. [Google Scholar] [CrossRef]
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected Fusarium toxins in Swedish wheat - Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Hellin, P.; Dedeurwaerder, G.; Duvivier, M.; Scauflaire, J.; Huybrechts, B.; Callebaut, A.; Munaut, F.; Legrève, A. Relationship between Fusarium spp. diversity and mycotoxin contents of mature grains in southern Belgium. Food Addit. Contam. Part A 2016, 33, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Hietaniemi, V.; Rämö, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sieviläinen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. Part A 2016, 33, 831–848. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Aamot, H.U.; Torp, T.; Jestoi, M.; Lattanzio, V.M.T.; Klemsdal, S.S.; Waalwijk, C.; van der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers’ fields in Norway over a six-year period. World Mycotoxin J. 2016, 9, 365–378. [Google Scholar] [CrossRef]
- Karlsson, I.; Friberg, H.; Kolseth, A.-K.; Steinberg, C.; Persson, P. Agricultural factors affecting Fusarium communities in wheat kernels. Int. J. Food Microbiol. 2017, 252, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzdraliński, A.; Nowak, M.; Szczerba, H.; Dudziak, K.; Muszyńska, M.; Leśniowska-Nowak, J. The composition of Fusarium species in wheat husks and grains in south-eastern Poland. J. Integr. Agric. 2017, 16, 1530–1536. [Google Scholar] [CrossRef] [Green Version]
- Manstretta, V.; Rossi, V. Effects of weather variables on ascospore discharge from Fusarium graminearum perithecia. PLoS ONE 2015, 10, e0138860. [Google Scholar] [CrossRef] [Green Version]
- ZMP—Zentrale Markt- und Preisberichtsstelle. ZMP-Marktbilanz Getreide, Ölsaaten, Futtermittel 2001; ZMP—Zentrale Markt- und Preisberichtsstelle GmbH: Bonn, Germany, 2001. [Google Scholar]
- Statistisches Bundesamt. Landwirtschaftliche Bodennutzung. Available online: https://www.destatis.de/GPStatistik/servlets/MCRFileNodeServlet/DEHeft_derivate_00004307/2030312118004.pdf (accessed on 10 March 2020).
- Fernandez, M.R.; Chen, Y. Pathogenicity of Fusarium species on different plant parts of spring wheat under controlled conditions. Plant Dis. 2005, 89, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-M.; Monger, W.; Ritieni, A.; Nicholson, P. Effect of temperature and duration of wetness during initial infection periods on disease development, fungal biomass and mycotoxin concentrations on wheat inoculated with single, or combinations of, Fusarium species. Plant Pathol. 2007, 56, 943–956. [Google Scholar] [CrossRef]
- Xu, X.; Nicholson, P.; Ritieni, A. Effects of fungal interactions among Fusarium head blight pathogens on disease development and mycotoxin accumulation. Int. J. Food Microbiol. 2007, 119, 67–71. [Google Scholar] [CrossRef]
- Lemmens, M.; Buerstmayr, H.; Krska, R.; Schuhmacher, R.; Grausgruber, H.; Ruckenbauer, P. The effect of inoculation treatment and long-term application of moisture on Fusarium head blight symptoms and deoxynivalenol contamination in wheat grains. Eur. J. Plant Pathol. 2004, 110, 299–308. [Google Scholar] [CrossRef]
- Lenc, L.; Czecholiński, G.; Wyczling, D.; Turów, T.; Kaźmierczak, A. Fusarium head blight (FHB) and Fusarium spp. on grain of spring wheat cultivars grown in Poland. J. Plant Prot. Res. 2015, 55, 266–277. [Google Scholar] [CrossRef]
- Rossi, V.; Ravanetti, A.; Pattori, E.; Giosuè, S. Influence of temperature and humidity on the infection of wheat spikes by some fungi causing Fusarium head blight. J. Plant Pathol. 2001, 83, 189–198. [Google Scholar]
- Thrane, U.; Adler, A.; Clasen, P.-E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial Location | Coordinates (ETRS89) | Crop Rotation | Previous Crop | Soil Cultivation | |
|---|---|---|---|---|---|
| xETRS | yETRS | ||||
| Barlt | 1004362 | 7173698 | SB-WW-WW | WW | Plough |
| Elskop | 1060012 | 7133704 | OR-WW-WW | WW | Plough |
| Futterkamp | 1183896 | 7225601 | OR-WW-WB | OR | Reduced tillage |
| Kastorf | 1175513 | 7123486 | OR-WW-WW | WW | Plough |
| Kluvensiek | 1092115 | 7234219 | OR-WW-WB | OR | Reduced tillage |
| Loit | 1080326 | 7286046 | OR-WW-WB | OR | Reduced tillage |
| Sönke-Nissen-Koog | 987384 | 7291393 | OR-WW-WW | WW | Plough |
| Species | 2013 | 2014 | 2015 | 2016 | 2017 | 2013–2017 |
|---|---|---|---|---|---|---|
| Fg | 100 | 76 | 57 | 71 | 100 | 81 |
| Fc | 100 | 71 | 57 | 100 | 100 | 86 |
| Fa | 100 | 67 | 71 | 62 | 100 | 80 |
| Fp | 100 | 81 | 52 | 95 | 100 | 86 |
| Ft | 71 | 67 | 48 | 67 | 81 | 67 |
| Fl | 76 | 62 | 52 | 57 | 76 | 65 |
| Effect | df | DNA Amount (pg Fungal DNA/ng Plant DNA) | DON (µg/kg) | ZEA (µg/kg) | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| Year (Y) | 4 | 5.315 | 0.0003 | 154.317 | <0.0001 | 129.643 | <0.0001 |
| Cultivar (Cv) | 2 | 8.932 | 0.0042 | 48.486 | <0.0001 | 64.051 | <0.0001 |
| Species (Spp) | 5 | 128.769 | <0.0001 | ||||
| Y × Cv | 8 | 1.328 | 0.2250 | 3.134 | 0.0022 | 5.740 | <0.0001 |
| Y × Spp | 20 | 19.284 | <0.0001 | ||||
| Cv × Spp | 10 | 8.453 | <0.0001 | ||||
| Y × Cv × Spp | 40 | 2.317 | <0.0001 | ||||
| Fa | Fc | Fg | Fl | Fp | |
|---|---|---|---|---|---|
| Fc | 0.562 | ||||
| Fg | 0.294 | 0.485 | |||
| Fl | 0.285 | 0.044 | 0.065 | ||
| Fp | 0.198 | 0.180 | 0.123 | 0.246 | |
| Ft | 0.287 | 0.079 | 0.056 | 0.284 | 0.473 |
| Cultivar Susceptibility | DNA Amount (pg Fungal DNA/ng Plant DNA) | Mycotoxin Content (µg/kg) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fg | Fc | Fa | Fp | Ft | Fl | DON | ZEA | ||
| 2017 | High | 5.25 ± 3.70 a | 1.08 ± 0.55 a | 0.88 ± 0.34 a | 0.54 ± 0.43 a | 0.05 ± 0.03 a | 0.08 ± 0.08 a | 987 ± 438 a | 146 ± 66 a |
| Moderate to high | 4.46 ± 3.53 ab | 0.64 ± 0.40 ab | 0.68 ± 0.40 a | 0.42 ± 0.33 a | 0.07 ± 0.03 a | 0.04 ± 0.03 a | 761 ± 357 a | 99 ± 50 a | |
| Low to moderate | 2.34 ± 1.41 b | 0.48 ± 0.29 b | 0.72 ± 0.46 a | 0.33 ± 0.25 a | 0.03 ± 0.04 a | 0.02 ± 0.03 a | 546 ± 226 b | 54 ± 26 b | |
| 2016 | High | 1.48 ± 1.93 a | 1.16 ± 1.32 a | 0.58 ± 0.94 a | 0.73 ± 0.55 a | 0.07 ± 0.09 a | 0.03 ± 0.04 a | 607 ± 392 a | 54 ± 49 a |
| Moderate to high | 0.96 ± 1.44 a | 0.79 ± 0.86 a | 0.38 ± 0.37 a | 0.89 ± 0.64 a | 0.09 ± 0.11 a | 0.02 ± 0.03 a | 463 ± 324 ab | 40 ± 42 a | |
| Low to moderate | 0.61 ± 0.99 a | 0.47 ± 0.89 a | 0.32 ± 0.59 a | 0.53 ± 0.53 a | 0.02 ± 0.03 a | 0.01 ± 0.02 a | 308 ± 252 b | 22 ± 25 a | |
| 2015 | High | 0.69 ± 1.40 a | 0.38 ± 0.42 a | 0.44 ± 0.52 a | 0.33 ± 0.56 a | 0.04 ± 0.05 a | 0.05 ± 0.05 a | 356 ± 103 a | 29 ± 22 a |
| Moderate to high | 0.46 ± 0.56 a | 0.18 ± 0.16 a | 0.42 ± 0.53 a | 0.22 ± 0.30 a | 0.04 ± 0.06 a | 0.05 ± 0.05 a | 273 ± 71 a | 18 ± 13 a | |
| Low to moderate | 0.34 ± 0.61 a | 0.17 ± 0.29 a | 0.27 ± 0.37 a | 0.14 ± 0.22 a | 0.02 ± 0.04 a | 0.04 ± 0.05 a | 113 ± 75 a | 11 ± 8 a | |
| 2014 | High | 1.08 ± 1.27 a | 0.45 ± 0.87 a | 0.26 ± 0.28 a | 0.60 ± 0.64 a | 0.01 ± 0.01 a | 0.07 ± 0.02 a | 366 ± 283 a | 62 ± 42 a |
| Moderate to high | 0.75 ± 1.03 a | 0.29 ± 0.42 a | 0.14 ± 0.16 a | 0.61 ± 0.86 a | 0.02 ± 0.02 a | 0.03 ± 0.02 a | 267 ± 193 a | 44 ± 30 a | |
| Low to moderate | 0.37 ± 0.71 a | 0.15 ± 0.27 a | 0.16 ± 0.10 a | 0.23 ± 0.28 a | 0.01 ± 0.01 a | 0.01 ± 0.02 a | 131 ± 98 a | 27 ± 17 a | |
| 2013 | High | 6.04 ± 2.69 a | 1.57 ± 0.60 a | 0.93 ± 0.59 a | 0.64 ± 0.44 a | 0.05 ± 0.06 a | 0.09 ± 0.06 a | 1186 ± 344 a | 189 ± 91 a |
| Moderate to high | 4.85 ± 2.13 ab | 1.01 ± 0.63 ab | 0.66 ± 0.39 a | 0.68 ± 0.39 a | 0.03 ± 0.03 a | 0.07 ± 0.04 a | 975 ± 212 a | 154 ± 58 a | |
| Low to moderate | 2.68 ± 1.18 b | 0.73 ± 0.58 b | 0.90 ± 0.87 a | 0.54 ± 0.56 a | 0.06 ± 0.12 a | 0.03 ± 0.03 a | 669 ± 116 b | 95 ± 34 b | |
| Species | Mycotoxin | Cultivar Susceptibility | ||
|---|---|---|---|---|
| High | Moderate to High | Low to Moderate | ||
| Fg | DON | 0.911 | 0.883 | 0.889 |
| Fc | DON | 0.645 | 0.622 | 0.589 |
| Fg | ZEA | 0.877 | 0.832 | 0.854 |
| Fc | ZEA | 0.497 | 0.596 | 0.418 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birr, T.; Hasler, M.; Verreet, J.-A.; Klink, H. Composition and Predominance of Fusarium Species Causing Fusarium Head Blight in Winter Wheat Grain Depending on Cultivar Susceptibility and Meteorological Factors. Microorganisms 2020, 8, 617. https://doi.org/10.3390/microorganisms8040617
Birr T, Hasler M, Verreet J-A, Klink H. Composition and Predominance of Fusarium Species Causing Fusarium Head Blight in Winter Wheat Grain Depending on Cultivar Susceptibility and Meteorological Factors. Microorganisms. 2020; 8(4):617. https://doi.org/10.3390/microorganisms8040617
Chicago/Turabian StyleBirr, Tim, Mario Hasler, Joseph-Alexander Verreet, and Holger Klink. 2020. "Composition and Predominance of Fusarium Species Causing Fusarium Head Blight in Winter Wheat Grain Depending on Cultivar Susceptibility and Meteorological Factors" Microorganisms 8, no. 4: 617. https://doi.org/10.3390/microorganisms8040617
APA StyleBirr, T., Hasler, M., Verreet, J.-A., & Klink, H. (2020). Composition and Predominance of Fusarium Species Causing Fusarium Head Blight in Winter Wheat Grain Depending on Cultivar Susceptibility and Meteorological Factors. Microorganisms, 8(4), 617. https://doi.org/10.3390/microorganisms8040617