
Predatory Bacteria Select for Sustained Prey Diversity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Predator-Prey Coevolution
2.2. Mucoid Frequency Estimation and Clone Isolation
2.3. Productivity and Predation Assays
2.4. Genomic Data
2.5. Statistical Analysis
3. Results
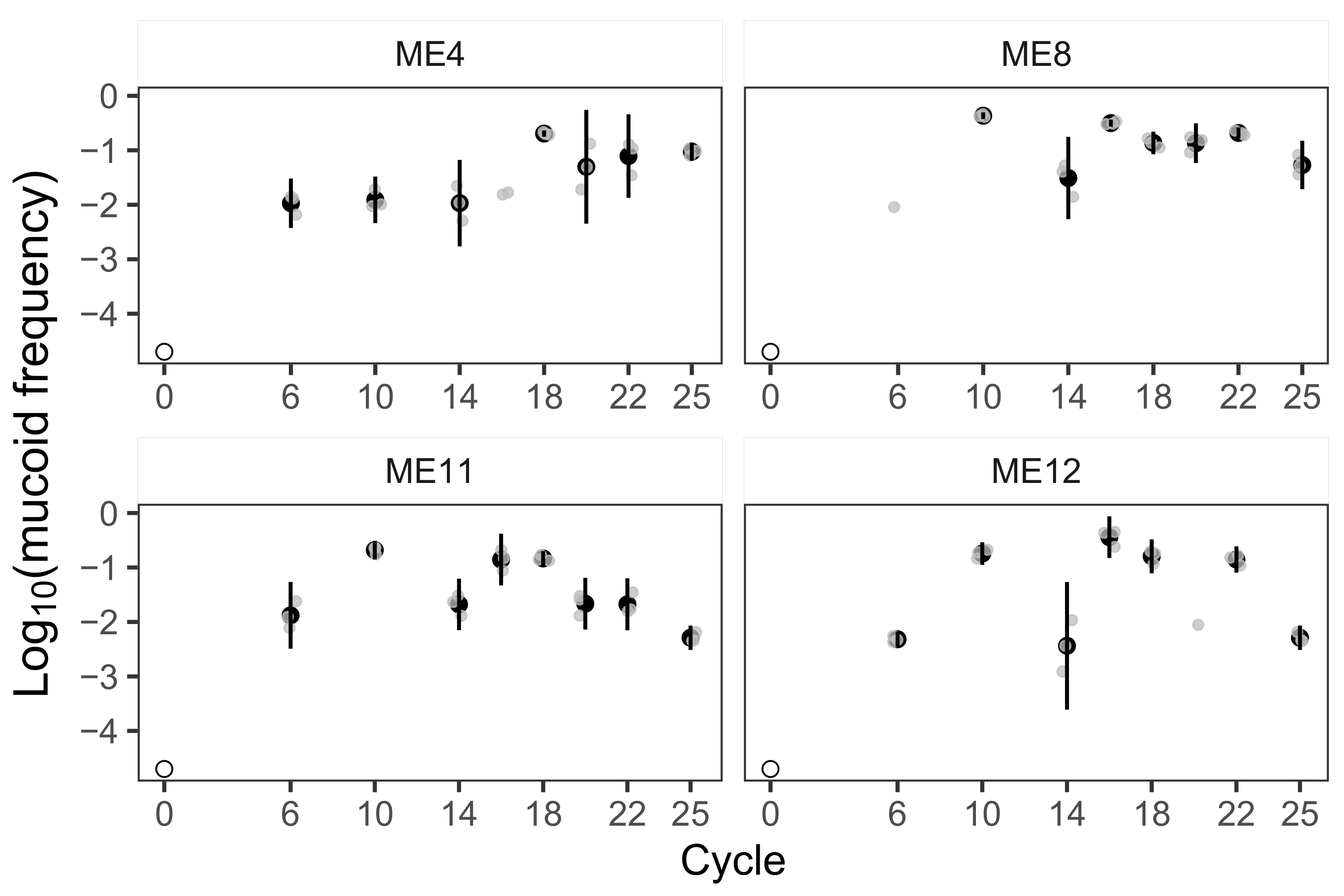
3.1. Mucoid Phenotypes Rise to and Remain within Intermediate Frequency Ranges
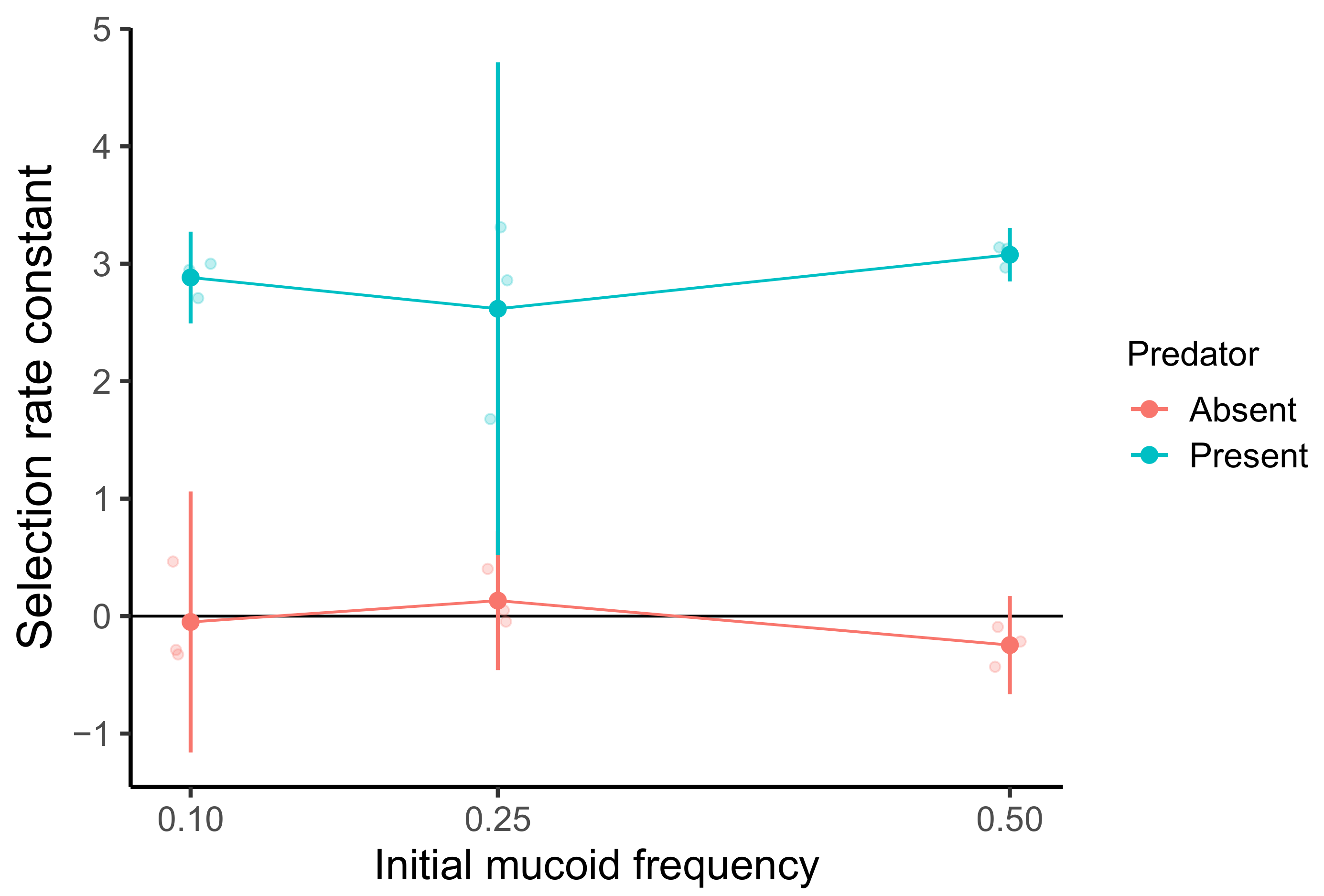
3.2. A Single-Cycle Competition Experiment Reveals a Predation-Specific Advantage to Mucoidy but Not NFD
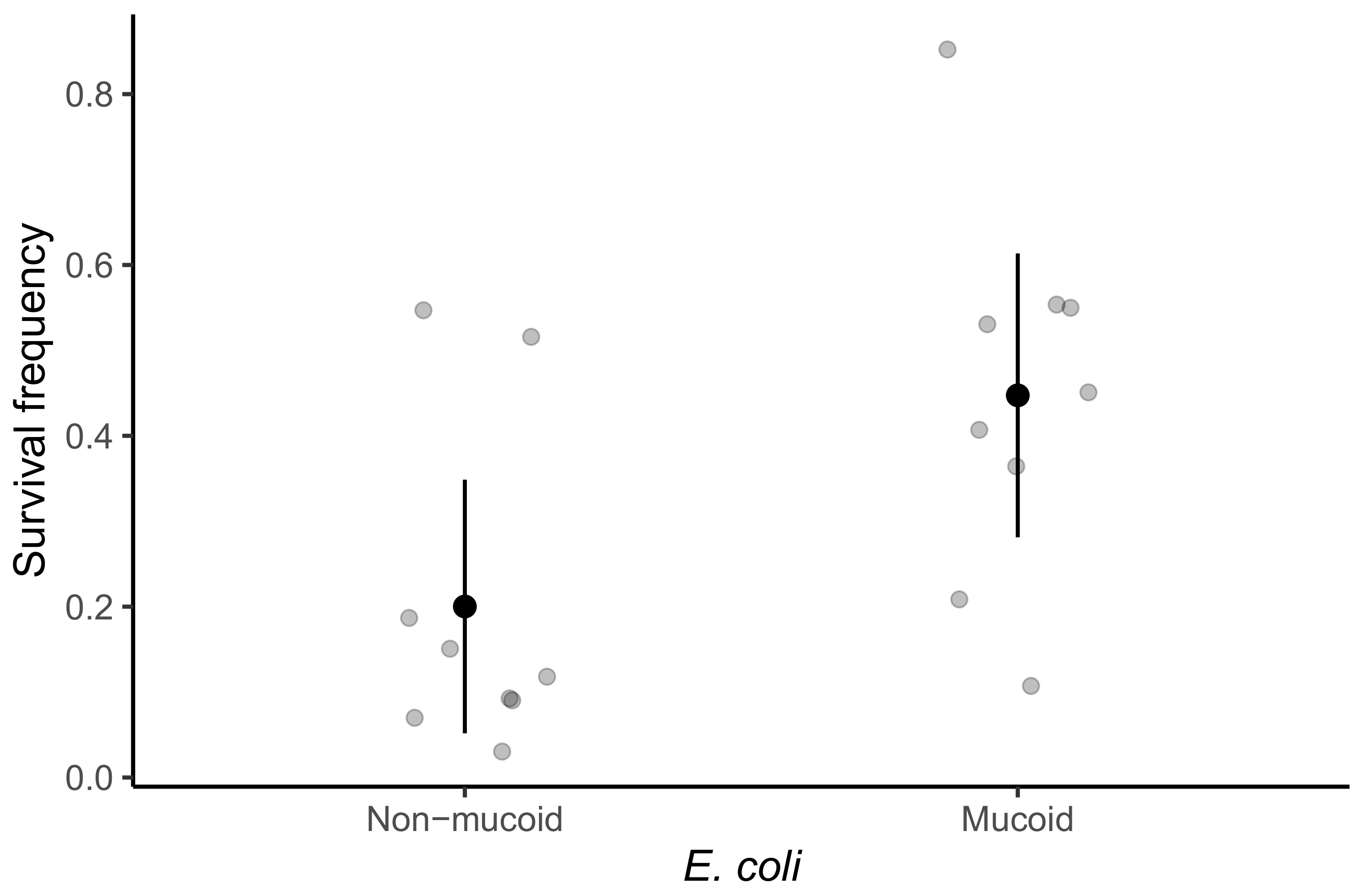
3.3. Mucoidy Is Associated with Low Prey Productivity in the Absence of Predators
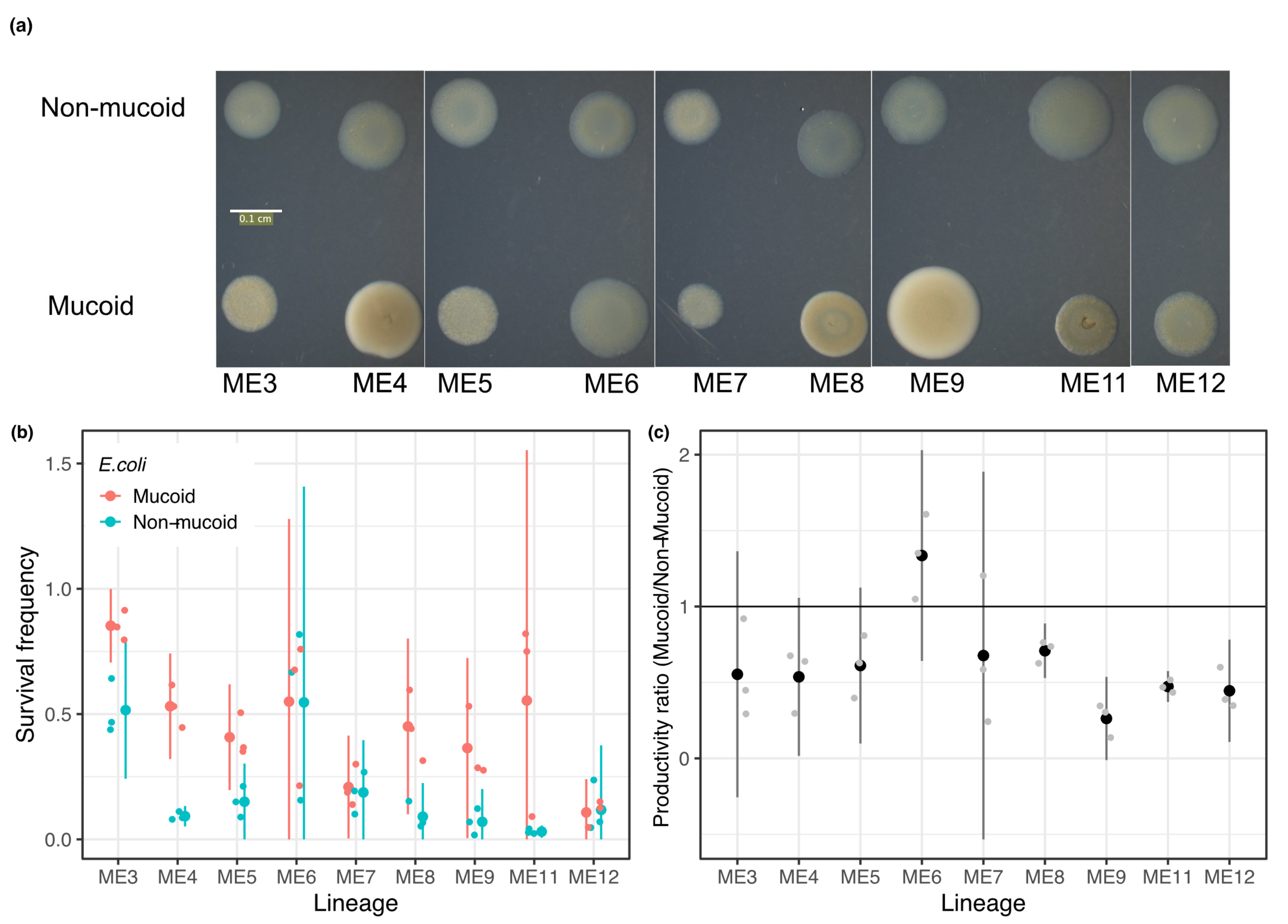
3.4. Mucoidy Is Phenotypically Variable
3.5. Candidate Mutations for Mucoidy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Estes, J.A.; Crooks, K.; Holt, R.D. Predators, Ecological Role of. In Encyclopedia of Biodiversity, 2nd ed.; Academic Press: Waltham, MA, USA, 2013; pp. 229–249. [Google Scholar] [CrossRef]
- Viola, D.V.; Mordecai, E.A.; Jaramillo, A.G.; Sistla, S.A.; Albertson, L.K.; Gosnell, J.S.; Cardinale, B.J.; Levine, J.M. Competition-defense tradeoffs and the maintenance of plant diversity. Proc. Natl. Acad. Sci. USA 2010, 107, 17217–17222. [Google Scholar] [CrossRef]
- Krediet, C.J.; Donahue, M.J. Growth-mortality trade-offs along a depth gradient in Cancer borealis. J. Exp. Mar. Biol. Ecol. 2009, 373, 133–139. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Fortner, A.M.; Fukami, T. The effects of resource enrichment, dispersal, and predation on local and metacommunity structure. Oecologia 2006, 149, 150–157. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Significance of predation by protists in aquatic microbial food webs. Antonie Van Leeuwenhoek 2002, 81, 293–308. [Google Scholar] [CrossRef]
- Rodríguez-Zaragoza, S. Ecology of Free-Living Amoebae. Crit. Rev. Microbiol. 1994, 20, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Shtonda, B.B.; Avery, L. Dietary choice behavior in Caenorhabditis elegans. J. Exp. Biol. 2006, 209, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.; Moraleda-Muñoz, A.; Marcos-Torres, F.J.; Muñoz-Dorado, J. Bacterial predation: 75 years and counting! Environ. Microbiol. 2016, 18, 766–779. [Google Scholar] [CrossRef]
- Williams, H.N.; Lymperopoulou, D.S.; Athar, R.; Chauhan, A.; Dickerson, T.L.; Chen, H.; Laws, E.; Berhane, T.K.; Flowers, A.R.; Bradley, N.; et al. Halobacteriovorax, an underestimated predator on bacteria: Potential impact relative to viruses on bacterial mortality. ISME J. 2016, 10, 491–499. [Google Scholar] [CrossRef]
- Corno, G.; Jurgens, K. Direct and Indirect Effects of Protist Predation on Population Size Structure of a Bacterial Strain with High Phenotypic Plasticity. Appl. Environ. Microbiol. 2006, 72, 78–86. [Google Scholar] [CrossRef]
- Queck, S.Y.; Weitere, M.; Moreno, A.M.; Rice, S.A.; Kjelleberg, S. The role of quorum sensing mediated developmental traits in the resistance of Serratia marcescens biofilms against protozoan grazing. Environ. Microbiol. 2006, 8, 1017–1025. [Google Scholar] [CrossRef]
- Müller, S.; Strack, S.N.; Hoefler, B.C.; Straight, P.D.; Kearns, D.B.; Kirby, J.R. Bacillaene and sporulation protect Bacillus subtilis from predation by Myxococcus xanthus. Appl. Environ. Microbiol. 2014, 80, 5603–5610. [Google Scholar] [CrossRef]
- Mazzola, M.; de Bruijn, I.; Cohen, M.F.; Raaijmakers, J.M. Protozoan-Induced Regulation of Cyclic Lipopeptide Biosynthesis Is an Effective Predation Defense Mechanism for Pseudomonas fluorescens. Appl. Environ. Microbiol. 2009, 75, 6804–6811. [Google Scholar] [CrossRef] [PubMed]
- Jousset, A. Ecological and evolutive implications of bacterial defences against predators. Environ. Microbiol. 2011, 14, 1830–1843. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Buckling, A. Co-evolution with lytic phage selects for the mucoid phenotype of Pseudomonas fluorescens SBW25. ISME J. 2012, 6, 1148–1158. [Google Scholar] [CrossRef]
- Koskella, B. Phage-Mediated Selection on Microbiota of a Long-Lived Host. Curr. Biol. 2013, 23, 1256–1260. [Google Scholar] [CrossRef] [PubMed]
- Wielgoss, S.; Bergmiller, T.; Bischofberger, A.M.; Hall, A.R. Adaptation to Parasites and Costs of Parasite Resistance in Mutator and Nonmutator Bacteria. Mol. Biol. Evol. 2016, 33, 770–782. [Google Scholar] [CrossRef]
- Muñoz-Dorado, J.; Marcos-Torres, F.J.; García-Bravo, E.; Moraleda-Muñoz, A.; Pérez, J. Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Front. Microbiol. 2016, 7, 781. [Google Scholar] [CrossRef]
- Berleman, J.E.; Zemla, M.; Remis, J.P.; Liu, H.; Davis, A.E.; Worth, A.N.; West, Z.; Zhang, A.; Park, H.; Bosneaga, E.; et al. Exopolysaccharide microchannels direct bacterial motility and organize multicellular behavior. ISME J. 2016, 10, 2620–2632. [Google Scholar] [CrossRef]
- Thiery, S.; Kaimer, C. The Predation Strategy of Myxococcus xanthus. Front. Microbiol. 2020, 11, 2. [Google Scholar] [CrossRef]
- Morgan, A.D.; MacLean, R.C.; Hillesland, K.L.; Velicer, G.J. Comparative analysis of Myxococcus predation on soil bacteria. Appl. Environ. Microbiol. 2010, 76, 6920–6927. [Google Scholar] [CrossRef]
- Rendueles, O.; Velicer, G.J. Hidden paths to endless forms most wonderful: Complexity of bacterial motility shapes diversification of latent phenotypes. BMC Evol. Biol. 2020, 20, 145. [Google Scholar] [CrossRef]
- Nair, R.R.; Vasse, M.; Wielgoss, S.; Sun, L.; Yu, Y.T.N.; Velicer, G.J. Bacterial predator-prey coevolution accelerates genome evolution and selects on virulence-associated prey defences. Nat. Commun. 2019, 10, 4301. [Google Scholar] [CrossRef] [PubMed]
- Piña, S.E.; Mattingly, S.J. The Role of Fluoroquinolones in the Promotion of Alginate Synthesis and Antibiotic Resistance in Pseudomonas aeruginosa. Curr. Microbiol. 1997, 35, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Miskinyte, M.; Sousa, A.; Ramiro, R.S.; de Sousa, J.A.M.; Kotlinowski, J.; Caramalho, I.; Magalhães, S.; Soares, M.P.; Gordo, I. The Genetic Basis of Escherichia coli Pathoadaptation to Macrophages. PLoS Pathog. 2013, 9, e1003802. [Google Scholar] [CrossRef] [PubMed]
- Landau, E.; Shapira, R. Effects of Subinhibitory Concentrations of Menthol on Adaptation, Morphological, and Gene Expression Changes in Enterohemorrhagic Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 5361–5367. [Google Scholar] [CrossRef]
- Paine, R.T. The Pisaster-Tegula Interaction: Prey Patches, Predator Food Preference, and Intertidal Community Structure. Ecology 1969, 50, 950–961. [Google Scholar] [CrossRef]
- Soto-Ortiz, L. The Regulation of Ecological Communities Through Feedback Loops: A Review. Res. Zool. 2015, 5, 1–15. [Google Scholar] [CrossRef]
- Menge, B.A.; Iles, A.C.; Freidenburg, T.L. Keystone Species. In Encyclopedia of Biodiversity, 2nd ed.; Academic Press: Waltham, MA, USA, 2013; pp. 442–457. [Google Scholar] [CrossRef]
- Bond, A.B.; Kamil, A.C. Apostatic selection by blue jays produces balanced polymorphism in virtual prey. Nature 1998, 395, 594–596. [Google Scholar] [CrossRef]
- Good, B.H.; McDonald, M.J.; Barrick, J.E.; Lenski, R.E.; Desai, M.M. The dynamics of molecular evolution over 60,000 generations. Nature 2017, 551, 45–50. [Google Scholar] [CrossRef]
- Lang, G.I.; Rice, D.P.; Hickman, M.J.; Sodergren, E.; Weinstock, G.M.; Botstein, D.; Desai, M.M. Pervasive genetic hitchhiking and clonal interference in forty evolving yeast populations. Nature 2013, 500, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Maddamsetti, R.; Lenski, R.E.; Barrick, J.E. Adaptation, clonal interference, and frequency-dependent interactions in a long-term evolution experiment with Escherichia coli. Genetics 2015, 200, 619–631. [Google Scholar] [CrossRef]
- Velicer, G.J. Pleiotropic effects of adaptation to a single carbon source for growth on alternative substrates. Appl. Environ. Microbiol. 1999, 65, 264–269. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Velicer, G.J.; Kroos, L.; Lenski, R.E. Developmental cheating in the social bacterium Myxococcus xanthus. Nature 2000, 404, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Fiegna, F.; Velicer, G.J. Competitive fates of bacterial social parasites: Persistence and self-induced extinction of Myxococcus xanthus cheaters. Proc. Biol. Sci. R. Soc. 2003, 270, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Ferrières, L.; Aslam, S.N.; Cooper, R.M.; Clarke, D.J. The yjbEFGH locus in Escherichia coli K-12 is an operon encoding proteins involved in exopolysaccharide production. Microbiology 2007, 153, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Gottesman, S. The Rcs phosphorelay: A Complex Signal Transduction System. Annu. Rev. Microbiol. 2005, 59, 379–405. [Google Scholar] [CrossRef] [PubMed]
- Corno, G.; Villiger, J.; Pernthaler, J. Coaggregation in a microbial predator-prey system affects competition and trophic transfer efficiency. Ecology 2013, 94, 870–881. [Google Scholar] [CrossRef]
- Menge, B.A.; Foley, M.M.; Robart, M.J.; Richmond, E.; Noble, M.; Chan, F. Keystone predation: Trait-based or driven by extrinsic processes? Assessment using a comparative-experimental approach. Ecol. Monogr. 2021, 91, e01436. [Google Scholar] [CrossRef]
- Vance, R.R. Predation and Resource Partitioning in One Predator—Two Prey Model Communities. Am. Nat. 1978, 112, 797–813. [Google Scholar] [CrossRef]
- Estévez, D.; Galindo, J.; Rolán-Alvarez, E. Negative frequency-dependent selection maintains shell banding polymorphisms in two marine snails (Littorina fabalis and Littorina saxatilis). Ecol. Evol. 2021, 11, 6381–6390. [Google Scholar] [CrossRef]
- Weir, J.C. Search Image-Driven Apostatic Selection and the Evolution of Phenotypic Polymorphism. eLS 2021, 2, 1–10. [Google Scholar] [CrossRef]
- Barrett, R.D.H.; Laurent, S.; Mallarino, R.; Pfeifer, S.P.; Xu, C.C.Y.; Foll, M.; Wakamatsu, K.; Duke-Cohan, J.S.; Jensen, J.D.; Hoekstra, H.E. Linking a mutation to survival in wild mice. Science 2019, 363, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Dragoš, A.; Lakshmanan, N.; Martin, M.; Horváth, B.; Maróti, G.; García, C.F.; Lieleg, O.; Kovács, T. Evolution of exploitative interactions during diversification in Bacillus subtilis biofilms. FEMS Microbiol. Ecol. 2018, 94, fix155. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Belkin, S. Overproduction of Exopolysaccharides by an Escherichia coli K-12 rpoS Mutant in Response to Osmotic Stress. Appl. Environ. Microbiol. 2009, 75, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Belkin, S. Simple quantification of bacterial envelope-associated extracellular materials. J. Microbiol. Methods 2009, 78, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Navasa, N.; Rodríguez-Aparicio, L.; Ferrero, M.; Monteagudo-Mera, A.; Martínez-Blanco, H. Polysialic and colanic acids metabolism in Escherichia coli K92 is regulated by RcsA and RcsB. Biosci. Rep. 2013, 33, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Lueders, T.; Kindler, R.; Miltner, A.; Friedrich, M.W.; Kaestner, M. Identification of bacterial micropredators distinctively active in a soil microbial food web. Appl. Environ. Microbiol. 2006, 72, 5342–5348. [Google Scholar] [CrossRef]
- Johnke, J.; Cohen, Y.; de Leeuw, M.; Kushmaro, A.; Jurkevitch, E.; Chatzinotas, A. Multiple micro-predators controlling bacterial communities in the environment. Curr. Opin. Biotechnol. 2014, 27, 185–190. [Google Scholar] [CrossRef]
- Petters, S.; Groß, V.; Söllinger, A.; Pichler, M.; Reinhard, A.; Bengtsson, M.M.; Urich, T. The soil microbial food web revisited: Predatory myxobacteria as keystone taxa? ISME J. 2021, 15, 2665–2675. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.M.; Novak, M.; Rudolf, V.H.; Schreiber, S.J.; Urban, M.C.; Vasseur, D.A. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar] [CrossRef]
- Padfield, D.; Vujakovic, A.; Paterson, S.; Griffiths, R.; Buckling, A.; Hesse, E. Evolution of diversity explains the impact of pre-adaptation of a focal species on the structure of a natural microbial community. ISME J. 2020, 14, 2877–2889. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Mutation | Gene Function | Population |
|---|---|---|---|
| rcsA | deletion (1 bp) | transcriptional regulator of colanic acid capsular biosynthesis | ME4 |
| yjbG | SNP | extracellular polysaccharide export outer-membrane associated protein | ME4 |
| yjbE | insertion (+9 bp) | extracellular polysaccharide production threonine-rich protein | ME8 |
| yjbH | SNP | DUF940 family extracellular polysaccharide protein | ME11 |
| yjbH | insertion (+8 bp) | DUF940 family extracellular polysaccharide protein | ME12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nair, R.R.; Velicer, G.J. Predatory Bacteria Select for Sustained Prey Diversity. Microorganisms 2021, 9, 2079. https://doi.org/10.3390/microorganisms9102079
Nair RR, Velicer GJ. Predatory Bacteria Select for Sustained Prey Diversity. Microorganisms. 2021; 9(10):2079. https://doi.org/10.3390/microorganisms9102079
Chicago/Turabian StyleNair, Ramith R., and Gregory J. Velicer. 2021. "Predatory Bacteria Select for Sustained Prey Diversity" Microorganisms 9, no. 10: 2079. https://doi.org/10.3390/microorganisms9102079