
Genetic Predisposition to Persistent Human Papillomavirus-Infection and Virus-Induced Cancers



Abstract
:1. Introduction
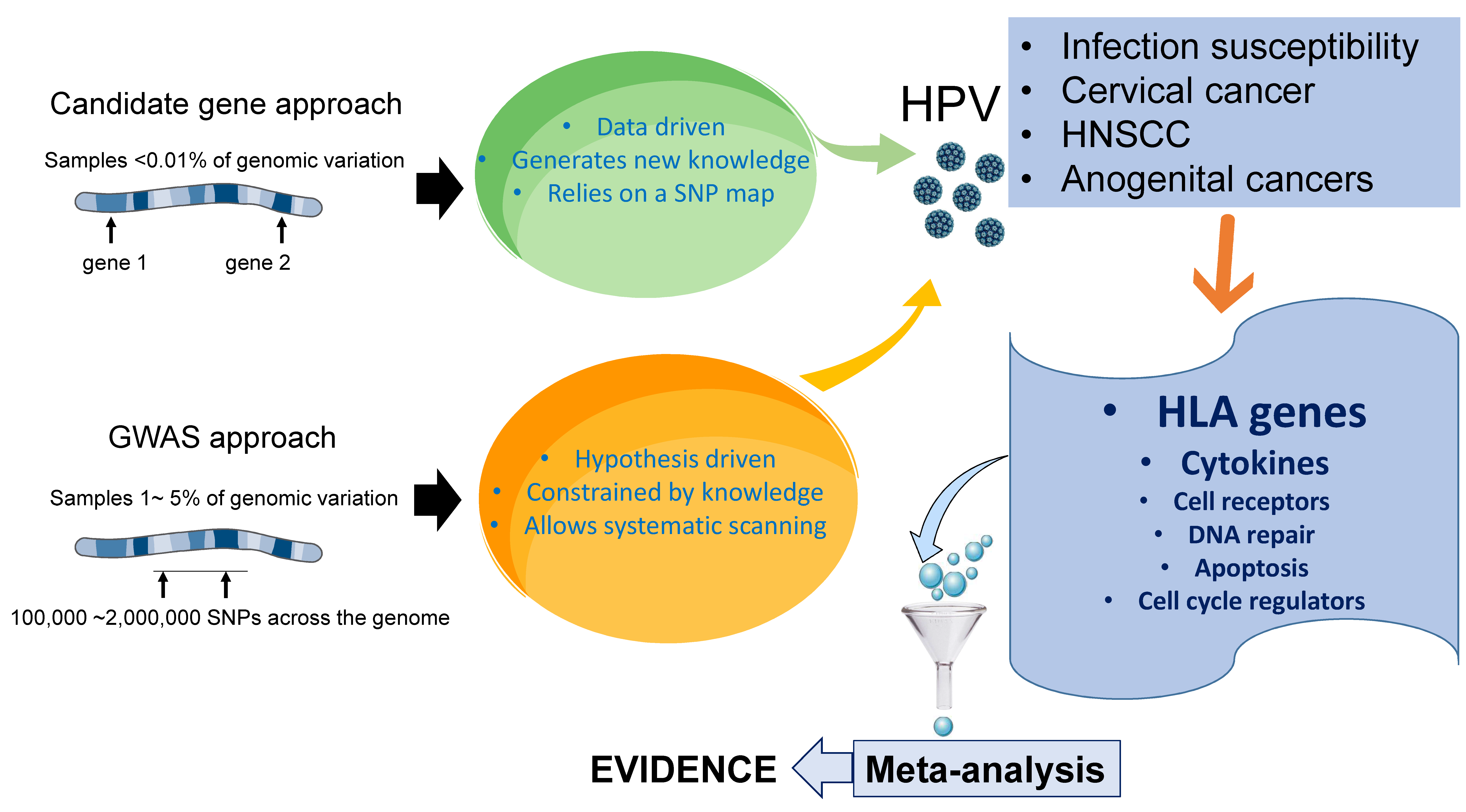
2. Candidate Genes versus Genome-Wide Associations in HPV Research
3. Genetic Variants and Susceptibility to HPV Infections
4. Genetic Variants and HPV-Induced Cervical Cancer
5. Genetic Variants and HPV-Induced Head and Neck Cancer
6. Genetic Variants and HPV-Induced Anogenital Cancers
7. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liebenberg, L.J.P.; McKinnon, L.R.; Yende-Zuma, N.; Garrett, N.; Baxter, C.; Kharsany, A.B.M.; Archary, D.; Rositch, A.; Samsunder, N.; Mansoor, L.E.; et al. HPV infection and the genital cytokine milieu in women at high risk of HIV acquisition. Nat. Commun. 2019, 10, 5227. [Google Scholar] [CrossRef]
- Wakabayashi, R.; Nakahama, Y.; Nguyen, V.; Espinoza, J.L. The Host-Microbe Interplay in Human Papillomavirus-Induced Carcinogenesis. Microorganisms 2019, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Näsman, A.; Du, J.; Dalianis, T. A global epidemic increase of an HPV-induced tonsil and tongue base cancer—Potential benefit from a pan-gender use of HPV vaccine. J. Intern. Med. 2020, 287, 134–152. [Google Scholar] [CrossRef] [Green Version]
- Latsuzbaia, A.; Wienecke-Baldacchino, A.; Tapp, J.; Arbyn, M.; Karabegović, I.; Chen, Z.; Fischer, M.; Mühlschlegel, F.; Weyers, S.; Pesch, P.; et al. Characterization and Diversity of 243 Complete Human Papillomavirus Genomes in Cervical Swabs Using Next Generation Sequencing. Viruses 2020, 12, 1437. [Google Scholar] [CrossRef]
- Della Fera, A.N.; Warburton, A.; Coursey, T.L.; Khurana, S.; McBride, A.A. Persistent Human Papillomavirus Infection. Viruses 2021, 13, 321. [Google Scholar] [CrossRef]
- de Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. J. Cancer 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olusola, P.; Banerjee, H.N.; Philley, J.V.; Dasgupta, S. Human Papilloma Virus-Associated Cervical Cancer and Health Disparities. Cells 2019, 8, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paaso, A.; Jaakola, A.; Syrjänen, S.; Louvanto, K. From HPV Infection to Lesion Progression: The Role of HLA Alleles and Host Immunity. Acta Cytol. 2019, 63, 148–158. [Google Scholar] [CrossRef]
- Bahrami, A.; Hasanzadeh, M.; Shahidsales, S.; Farazestanian, M.; Hassanian, S.M.; Moetamani Ahmadi, M.; Maftouh, M.; Gharib, M.; Yousefi, Z.; Kadkhodayan, S.; et al. Genetic susceptibility in cervical cancer: From bench to bedside. J. Cell Physiol. 2018, 233, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Guidry, J.T.; Scott, R.S. The interaction between human papillomavirus and other viruses. Virus Res. 2017, 231, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wen, X. Seropositivity to herpes simplex virus type 2, but not type 1 is associated with cervical cancer: NHANES (1999-2014). BMC Cancer 2017, 17, 726. [Google Scholar] [CrossRef] [Green Version]
- Escarcega-Tame, M.A.; López-Hurtado, M.; Escobedo-Guerra, M.R.; Reyes-Maldonado, E.; Castro-Escarpulli, G.; Guerra-Infante, F.M. Co-infection between genotypes of the human papillomavirus and. Int. J. STD AIDS 2020, 31, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Brusselaers, N.; Shrestha, S.; van de Wijgert, J.; Verstraelen, H. Vaginal dysbiosis and the risk of human papillomavirus and cervical cancer: Systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2018, 221, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.J.; Bodinier, B.; Kalliala, I.; Zuber, V.; Vuckovic, D.; Doulgeraki, T.; Whitaker, M.D.; Wielscher, M.; Cartwright, R.; Tsilidis, K.K.; et al. Genetic variation in cervical preinvasive and invasive disease: A genome-wide association study. Lancet Oncol. 2021, 22, 548–557. [Google Scholar] [CrossRef]
- Takeuchi, F.; Kukimoto, I.; Li, Z.; Li, S.; Li, N.; Hu, Z.; Takahashi, A.; Inoue, S.; Yokoi, S.; Chen, J.; et al. Genome-wide association study of cervical cancer suggests a role for ARRDC3 gene in human papillomavirus infection. Hum. Mol. Genet. 2019, 28, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Cui, T.; Ek, W.E.; Liu, H.; Wang, H.; Gyllensten, U. Analysis of the genetic architecture of susceptibility to cervical cancer indicates that common SNPs explain a large proportion of the heritability. Carcinogenesis 2015, 36, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Ma, H.; Lu, F.; Yuan, Z.; Wang, R.; Jiang, H.; Hu, Z.; Shen, H.; Chen, N. Genetic variants at 4q23 and 12q24 are associated with head and neck cancer risk in China. Mol. Carcinog. 2013, 52 (Suppl. 1), E2–E9. [Google Scholar] [CrossRef]
- Chen, D.; Juko-Pecirep, I.; Hammer, J.; Ivansson, E.; Enroth, S.; Gustavsson, I.; Feuk, L.; Magnusson, P.K.; McKay, J.D.; Wilander, E.; et al. Genome-wide association study of susceptibility loci for cervical cancer. J. Natl. Cancer Inst. 2013, 105, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Adebamowo, S.N.; Adeyemo, A.A.; Rotimi, C.N.; Olaniyan, O.; Offiong, R.; Adebamowo, C.A.; Group, H.A.A.R. Genome-wide association study of prevalent and persistent cervical high-risk human papillomavirus (HPV) infection. BMC Med. Genet. 2020, 21, 231. [Google Scholar] [CrossRef]
- Amos, W.; Driscoll, E.; Hoffman, J.I. Candidate genes versus genome-wide associations: Which are better for detecting genetic susceptibility to infectious disease? Proc. Biol. Sci. 2011, 278, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Takami, A. Role of non-HLA gene polymorphisms in graft-versus-host disease. Int. J. Hematol. 2013, 98, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, L.E.; Ostacher, M.; Ballon, J. How genome-wide association studies (GWAS) made traditional candidate gene studies obsolete. Neuropsychopharmacology 2019, 44, 1518–1523. [Google Scholar] [CrossRef] [PubMed]
- Alizon, S.; Murall, C.L.; Bravo, I.G. Why Human Papillomavirus Acute Infections Matter. Viruses 2017, 9, 293. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Petersen, J.; Rossjohn, J.; Fugger, L. HLA variation and disease. Nat. Rev. Immunol. 2018, 18, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Wang, C.; Aissani, B.; Wilson, C.M.; Tang, J.; Kaslow, R.A. Interleukin-10 gene (IL10) polymorphisms and human papillomavirus clearance among immunosuppressed adolescents. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 1626–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, E.; Kay, R.; Sillibourne, J.; Patel, P.; Hutchinson, I.; Woo, P. Polymorphic haplotypes of the interleukin-10 5′ flanking region determine variable interleukin-10 transcription and are associated with particular phenotypes of juvenile rheumatoid arthritis. Arthritis Rheum. 1999, 42, 1101–1108. [Google Scholar] [CrossRef]
- Maciag, P.C.; Schlecht, N.F.; Souza, P.S.; Franco, E.L.; Villa, L.L.; Petzl-Erler, M.L. Major histocompatibility complex class II polymorphisms and risk of cervical cancer and human papillomavirus infection in Brazilian women. Cancer Epidemiol. Biomarkers Prev. 2000, 9, 1183–1191. [Google Scholar]
- Bernal-Silva, S.; Granados, J.; Gorodezky, C.; Aláez, C.; Flores-Aguilar, H.; Cerda-Flores, R.M.; Guerrero-González, G.; Valdez-Chapa, L.D.; Morales-Casas, J.; González-Guerrero, J.F.; et al. HLA-DRB1 Class II antigen level alleles are associated with persistent HPV infection in Mexican women; a pilot study. Infect Agent Cancer 2013, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, S.; Roger, M.; Faucher, M.C.; Coutlée, F.; Franco, E.L.; Brassard, P. The association between human leukocyte antigen (HLA)-G polymorphisms and human papillomavirus (HPV) infection in Inuit women of northern Quebec. Hum. Immunol. 2013, 74, 1610–1615. [Google Scholar] [CrossRef]
- Ferguson, R.; Ramanakumar, A.V.; Richardson, H.; Tellier, P.P.; Coutlée, F.; Franco, E.L.; Roger, M. Human leukocyte antigen (HLA)-E and HLA-G polymorphisms in human papillomavirus infection susceptibility and persistence. Hum. Immunol. 2011, 72, 337–341. [Google Scholar] [CrossRef]
- Del Río-Ospina, L.; Camargo, M.; Soto-De León, S.C.; Sánchez, R.; Moreno-Pérez, D.A.; Patarroyo, M.E.; Patarroyo, M.A. Identifying the HLA DRB1-DQB1 molecules and predicting epitopes associated with high-risk HPV infection clearance and redetection. Sci. Rep. 2020, 10, 7306. [Google Scholar] [CrossRef]
- Adebamowo, S.N.; Adeyemo, A.A.; Consortium, A.R.G.a.p.o.t.H.A. Classical HLA alleles are associated with prevalent and persistent cervical high-risk HPV infection in African women. Hum. Immunol. 2019, 80, 723–730. [Google Scholar] [CrossRef]
- Hart, G.T.; Hogquist, K.A.; Jameson, S.C. Krüppel-like factors in lymphocyte biology. J. Immunol. 2012, 188, 521–526. [Google Scholar] [CrossRef]
- Hu, Z.; Zhu, D.; Wang, W.; Li, W.; Jia, W.; Zeng, X.; Ding, W.; Yu, L.; Wang, X.; Wang, L.; et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat. Genet. 2015, 47, 158–163. [Google Scholar] [CrossRef]
- Mainali, B.; Schabath, M.B.; Sudenga, S.L.; Ye, Y.; Wiener, H.W.; Villa, L.L.; Giuliano, A.R.; Shrestha, S. Variants in immune-related genes and genital HPV 16 persistence in men. Papillomavirus Res. 2019, 7, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.A.; Luhn, P.; Gage, J.C.; Bodelon, C.; Dunn, S.T.; Walker, J.; Zuna, R.; Hewitt, S.; Killian, J.K.; Yan, L.; et al. Discovery and validation of candidate host DNA methylation markers for detection of cervical precancer and cancer. Int. J. Cancer 2017, 141, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Blom, A.M. The role of complement inhibitors beyond controlling inflammation. J. Intern. Med. 2017, 282, 116–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffman, M.; Glass, A.G.; Wentzensen, N.; Rush, B.B.; Castle, P.E.; Scott, D.R.; Buckland, J.; Sherman, M.E.; Rydzak, G.; Kirk, P.; et al. A long-term prospective study of type-specific human papillomavirus infection and risk of cervical neoplasia among 20,000 women in the Portland Kaiser Cohort Study. Cancer Epidemiol. Biomarkers Prev. 2011, 20, 1398–1409. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, H.; Zhang, L.; Qiao, Y. Cervical cancer: Epidemiology, risk factors and screening. Chin. J. Cancer Res. 2020, 32, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Sekine, M.; Hanley, S.J.B.; Kudo, R.; Hara, M.; Adachi, S.; Ueda, Y.; Miyagi, E.; Enomoto, T. Risk factors for HPV infection and high-grade cervical disease in sexually active Japanese women. Sci. Rep. 2021, 11, 2898. [Google Scholar] [CrossRef] [PubMed]
- Leo, P.J.; Madeleine, M.M.; Wang, S.; Schwartz, S.M.; Newell, F.; Pettersson-Kymmer, U.; Hemminki, K.; Hallmans, G.; Tiews, S.; Steinberg, W.; et al. Defining the genetic susceptibility to cervical neoplasia-A genome-wide association study. PLoS Genet. 2017, 13, e1006866. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.H.; Yan, W.H.; Lin, A. The Role of HLA-G in Human Papillomavirus Infections and Cervical Carcinogenesis. Front Immunol. 2020, 11, 1349. [Google Scholar] [CrossRef]
- Fahim, N.M.; Shehata, I.H.; Taha, S.E.; Fahmy, R.A.; Elsayed, M.S. Human Leukocyte Antigen-G (HLA-G) Expression in Precancerous and Cancerous Cervical Lesions: Association with Human Papilloma Virus Infection and Host Immune Response. Egypt J. Immunol. 2018, 25, 125–134. [Google Scholar]
- Alifu, M.; Fan, P.; Kuerban, G.; Yao, X.; Peng, Y.; Dong, T.; Wang, R. Frequency distribution of HLA alleles and haplotypes in Uyghur women with advanced squamous cell cervical cancer and relation to HPV status and clinical outcome. Arch. Gynecol. Obstet. 2018, 297, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Kamiza, A.B.; Kamiza, S.; Mathew, C.G. HLA-DRB1 alleles and cervical cancer: A meta-analysis of 36 case-control studies. Cancer Epidemiol. 2020, 67, 101748. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Guo, Y.; Zhan, S.; Xia, P. Association between HLA-DP Gene Polymorphisms and Cervical Cancer Risk: A Meta-Analysis. Biomed. Res. Int. 2018, 2018, 7301595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.M.; Mooij, M.; Branković, I.; Ouburg, S.; Morré, S.A.; Jordanova, E.S. Cervical Carcinogenesis and Immune Response Gene Polymorphisms: A Review. J. Immunol. Res. 2017, 2017, 8913860. [Google Scholar] [CrossRef]
- Sobti, R.C.; Kordi Tamandani, D.M.; Shekari, M.; Kaur, P.; Malekzadeh, K.; Suri, V. Interleukin 1 beta gene polymorphism and risk of cervical cancer. Int. J. Gynaecol. Obstet. 2008, 101, 47–52. [Google Scholar] [CrossRef]
- Chagas, B.S.; Lima, R.C.P.; Paiva Júnior, S.S.L.; Silva, R.C.O.; Cordeiro, M.N.; Silva Neto, J.D.C.; Batista, M.V.A.; Silva, A.J.D.; Gurgel, A.P.A.D.; Freitas, A.C. Significant association between IL10-1082/-819 and TNF-308 haplotypes and the susceptibility to cervical carcinogenesis in women infected by Human papillomavirus. Cytokine 2019, 113, 99–104. [Google Scholar] [CrossRef]
- Quan, Y.; Zhou, B.; Wang, Y.; Duan, R.; Wang, K.; Gao, Q.; Shi, S.; Song, Y.; Zhang, L.; Xi, M. Association between IL17 polymorphisms and risk of cervical cancer in Chinese women. Clin. Dev. Immunol. 2012, 2012, 258293. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.B.; Louvanto, K.; Ramanakumar, A.V.; Franco, E.L.; Villa, L.L.; For The Ludwig-McGill Cohort Study. Polymorphism in the promoter region of the Toll-like receptor 9 gene and cervical human papillomavirus infection. J. Gen. Virol. 2013, 94, 1858–1864. [Google Scholar] [CrossRef]
- Liu, N.; Song, Y.; Shi, W. IFN-γ +874 T/A polymorphisms contributes to cervical cancer susceptibility: A meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 4008–4015. [Google Scholar] [PubMed]
- Rotar, I.C.; Muresan, D.; Radu, P.; Petrisor, F.; Apostol, S.; Mariana, T.; Butuza, C.; Stamatian, F. TNF-α 308 G/A polymorphism and cervical intraepithelial neoplasia. Anticancer Res. 2014, 34, 373–378. [Google Scholar]
- Tavares, M.C.; de Lima Júnior, S.F.; Coelho, A.V.; Marques, T.R.; de Araújo, D.H.; Heráclio, S.e.A.; Amorim, M.M.; de Souza, P.R.; Crovella, S. Tumor necrosis factor (TNF) alpha and interleukin (IL) 18 genes polymorphisms are correlated with susceptibility to HPV infection in patients with and without cervical intraepithelial lesion. Ann. Hum. Biol. 2016, 43, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.G.; Schwartz, S.M.; Malkki, M.; Du, Q.; Petersdorf, E.W.; Galloway, D.A.; Madeleine, M.M. Risk of cervical cancer associated with allergies and polymorphisms in genes in the chromosome 5 cytokine cluster. Cancer Epidemiol. Biomarkers Prev. 2011, 20, 199–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Sun, H.; Jia, Y.; Tang, F.; Zhou, H.; Li, X.; Zhou, J.; Huang, K.; Zhang, Q.; Hu, T.; et al. Association of 42 SNPs with genetic risk for cervical cancer: An extensive meta-analysis. BMC Med. Genet. 2015, 16, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.S.; Bratti, M.C.; Rodríguez, A.C.; Herrero, R.; Burk, R.D.; Porras, C.; González, P.; Sherman, M.E.; Wacholder, S.; Lan, Z.E.; et al. Common variants in immune and DNA repair genes and risk for human papillomavirus persistence and progression to cervical cancer. J. Infect. Dis. 2009, 199, 20–30. [Google Scholar] [CrossRef]
- Burmeister, A.R.; Marriott, I. The Interleukin-10 Family of Cytokines and Their Role in the CNS. Front. Cell. Neurosci. 2018, 12, 458. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Eskdale, J.; Gallagher, G.; Verweij, C.L.; Keijsers, V.; Westendorp, R.G.; Huizinga, T.W. Interleukin 10 secretion in relation to human IL-10 locus haplotypes. Proc. Natl. Acad. Sci. USA 1998, 95, 9465–9470. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Oki, A.; Satoh, T.; Okada, S.; Minaguchi, T.; Onuki, M.; Ochi, H.; Nakao, S.; Sakurai, M.; Abe, A.; et al. Interleukin-10 -1082 gene polymorphism and susceptibility to cervical cancer among Japanese women. Jpn. J. Clin. Oncol. 2010, 40, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Chagas, B.S.; Gurgel, A.P.; da Cruz, H.L.; Amaral, C.M.; Cardoso, M.V.; Silva Neto, J.a.C.; da Silva, L.A.; de Albuquerque, E.M.; Muniz, M.T.; de Freitas, A.C. An interleukin-10 gene polymorphism associated with the development of cervical lesions in women infected with Human Papillomavirus and using oral contraceptives. Infect. Genet. Evol. 2013, 19, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.P.L.; Trugilo, K.P.; Okuyama, N.C.M.; Sena, M.M.; Couto-Filho, J.D.; Watanabe, M.A.E.; de Oliveira, K.B. IL-10 c.-592C>A (rs1800872) polymorphism is associated with cervical cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 1971–1978. [Google Scholar] [CrossRef]
- Torres-Poveda, K.; Burguete-García, A.I.; Cruz, M.; Martínez-Nava, G.A.; Bahena-Román, M.; Ortíz-Flores, E.; Ramírez-González, A.; López-Estrada, G.; Delgado-Romero, K.; Madrid-Marina, V. The SNP at -592 of human IL-10 gene is associated with serum IL-10 levels and increased risk for human papillomavirus cervical lesion development. Infect. Agent Cancer 2012, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Singhal, P.; Kumar, A.; Bharadwaj, S.; Hussain, S.; Bharadwaj, M. Association of IL-10 GTC haplotype with serum level and HPV infection in the development of cervical carcinoma. Tumour Biol. 2015, 36, 2287–2298. [Google Scholar] [CrossRef]
- Zidi, S.; Gazouani, E.; Stayoussef, M.; Mezlini, A.; Ahmed, S.K.; Yacoubi-Loueslati, B.; Almawi, W.Y. IL-10 gene promoter and intron polymorphisms as genetic biomarkers of cervical cancer susceptibility among Tunisians. Cytokine 2015, 76, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Wen, L.; Song, J.K.; Zeng, W.J.; Dan, C.; Niu, Y.M.; Shen, M. Significant association between interleukin-10 gene polymorphisms and cervical cancer risk: A meta-analysis. Oncotarget 2018, 9, 12365–12375. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.M.; Du, X.Y.; Cai, H.X.; Zhang, C.; Yuan, R.X.; Zeng, X.T.; Luo, J. Increased risks between Interleukin-10 gene polymorphisms and haplotype and head and neck cancer: A meta-analysis. Sci. Rep. 2015, 5, 17149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Omariba, G.; Xu, F.; Wang, M.; Li, K.; Zhou, Y.; Xiao, J. Genome-Wide Analysis of MicroRNA-related Single Nucleotide Polymorphisms (SNPs) in Mouse Genome. Sci. Rep. 2020, 10, 5789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, N.; Singhal, P.; Mehrotra, R.; Bharadwaj, M. Impacts of single nucleotide polymorphisms in three microRNAs (miR-146a, miR-196a2 and miR-499) on the susceptibility to cervical cancer among Indian women. Biosci. Rep. 2019, 39, BSR20180723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, D.; Zhou, W.; Jia, M.; Wang, L.; Zhou, J.; Yin, Y.; Ma, H.; Hu, Z.; Li, N.; Shen, H. Genetic variants within microRNA-binding site of RAD51B are associated with risk of cervical cancer in Chinese women. Cancer Med. 2016, 5, 2596–2601. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, H.; Ding, B.; Feng, X.; Zhao, W.; Cui, M.; Xu, Y.; Shi, M.; Chen, J.; Jin, H. Genetic variants in microRNAs are associated with cervical cancer risk. Mutagenesis 2019, 34, 127–133. [Google Scholar] [CrossRef]
- Mi, Y.; Wang, L.; Zong, L.; Pei, M.; Lu, Q.; Huang, P. Genetic variants in microRNA target sites of 37 selected cancer-related genes and the risk of cervical cancer. PLoS ONE 2014, 9, e86061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Dong, P.; Zhou, L.; Wang, S. The Association between Five Genetic Variants in MicroRNAs (rs2910164, rs11614913, rs3746444, rs11134527, and rs531564) and Cervical Cancer Risk: A Meta-Analysis. Biomed. Res. Int. 2021, 2021, 9180874. [Google Scholar] [CrossRef]
- Moszyńska, A.; Gebert, M.; Collawn, J.F.; Bartoszewski, R. SNPs in microRNA target sites and their potential role in human disease. Open Biol. 2017, 7, 170019. [Google Scholar] [CrossRef]
- Espinoza, J.L.; Minami, M. Sensing Bacterial-Induced DNA Damaging Effects. Front. Immunol. 2018, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Enroth, S.; Liu, H.; Sun, Y.; Wang, H.; Yu, M.; Deng, L.; Xu, S.; Gyllensten, U. Pooled analysis of genome-wide association studies of cervical intraepithelial neoplasia 3 (CIN3) identifies a new susceptibility locus. Oncotarget 2016, 7, 42216–42224. [Google Scholar] [CrossRef]
- Shi, Y.; Li, L.; Hu, Z.; Li, S.; Wang, S.; Liu, J.; Wu, C.; He, L.; Zhou, J.; Li, Z.; et al. A genome-wide association study identifies two new cervical cancer susceptibility loci at 4q12 and 17q12. Nat. Genet. 2013, 45, 918–922. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.; Kasamatsu, A.; Ishige, S.; Koike, K.; Saito, K.; Kouzu, Y.; Koike, H.; Sakamoto, Y.; Ogawara, K.; Shiiba, M.; et al. Exocyst complex component Sec8: A presumed component in the progression of human oral squamous-cell carcinoma by secretion of matrix metalloproteinases. J. Cancer Res. Clin. Oncol. 2013, 139, 533–542. [Google Scholar] [CrossRef]
- Łaźniak, S.; Roszak, A.; Balcerek, A.; Wareńczak-Florczak, Ż.; Prokop, E.; Sowińska, A.; Sotiri, E.; Tsibulski, A.; Sajdak, S.; Jagodziński, P.P. Role of rs13117307 single nuclear polymorphism in the risk of uterine cervical cancer from Polish population and its impact on exocyst complex component 1 expression. Gene 2017, 627, 524–529. [Google Scholar] [CrossRef]
- Liu, X.; Xia, S.; Zhang, Z.; Wu, H.; Lieberman, J. Channelling inflammation: Gasdermins in physiology and disease. Nat Rev Drug Discov. 2021, 20, 384–405. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Mishima, H.; Kinoshita, A.; Hayashida, C.; Abe, S.; Tokunaga, K.; Masuzaki, H.; Yoshiura, K. Genome-wide association study of HPV-associated cervical cancer in Japanese women. J. Med. Virol. 2014, 86, 1153–1158. [Google Scholar] [CrossRef]
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef]
- Giuliano, A.R.; Tortolero-Luna, G.; Ferrer, E.; Burchell, A.N.; de Sanjose, S.; Kjaer, S.K.; Muñoz, N.; Schiffman, M.; Bosch, F.X. Epidemiology of human papillomavirus infection in men, cancers other than cervical and benign conditions. Vaccine 2008, 26 (Suppl. 10), K17–K28. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Gu, Y.; Hou, W.; Wu, S.; Lou, Y.; Yang, W.; Zhu, L.; Hu, Y.; Sun, M.; Xue, H. P53 codon 72 polymorphism, human papillomavirus infection, and their interaction to oral carcinoma susceptibility. BMC Genet. 2015, 16, 72. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Sturgis, E.M.; Etzel, C.J.; Wei, Q.; Li, G. p73 G4C14-to-A4T14 polymorphism and risk of human papillomavirus-associated squamous cell carcinoma of the oropharynx in never smokers and never drinkers. Cancer 2008, 113, 3307–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Sturgis, E.M.; Zhao, C.; Etzel, C.J.; Wei, Q.; Li, G. Association of p73 G4C14-to-A4T14 polymorphism with human papillomavirus type 16 status in squamous cell carcinoma of the head and neck in non-Hispanic whites. Cancer 2009, 115, 1660–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sturgis, E.M.; Guo, W.; Song, X.; Zhang, F.; Xu, L.; Wei, Q.; Li, G. Association of combined p73 and p53 genetic variants with tumor HPV16-positive oropharyngeal cancer. PLoS ONE 2012, 7, e35522. [Google Scholar] [CrossRef]
- Lu, Z.; Zhang, H.; Tao, Y.; Li, X.; Li, G. genetic variants predict HPV16-positive tumors of patients with squamous cell carcinoma of the oropharynx. Oncotarget 2017, 8, 86710–86717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Sturgis, E.M.; Lei, D.; Liu, Z.; Dahlstrom, K.R.; Wei, Q.; Li, G. Association of TGF-beta1 genetic variants with HPV16-positive oropharyngeal cancer. Clin. Cancer Res. 2010, 16, 1416–1422. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Sturgis, E.M.; Huang, Z.; Wang, Y.; Wei, P.; Wang, J.R.; Wei, Q.; Li, G. Genetic Variants Predict Clinical Outcomes of HPV-Positive Oropharyngeal Cancer Patients after Definitive Radiotherapy. Clin. Cancer Res. 2018, 24, 2225–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Wang, X.; Cai, C.; Yang, S.; Huo, N.; Liu, H. Association between TGF-β1 Polymorphisms and Head and Neck Cancer Risk: A Meta-Analysis. Front. Genet. 2017, 8, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, J.D.; Truong, T.; Gaborieau, V.; Chabrier, A.; Chuang, S.C.; Byrnes, G.; Zaridze, D.; Shangina, O.; Szeszenia-Dabrowska, N.; Lissowska, J.; et al. A genome-wide association study of upper aerodigestive tract cancers conducted within the INHANCE consortium. PLoS Genet. 2011, 7, e1001333. [Google Scholar] [CrossRef]
- Liang, C.; Marsit, C.J.; Houseman, E.A.; Butler, R.; Nelson, H.H.; McClean, M.D.; Kelsey, K.T. Gene-environment interactions of novel variants associated with head and neck cancer. Head Neck 2012, 34, 1111–1118. [Google Scholar] [CrossRef]
- Levovitz, C.; Chen, D.; Ivansson, E.; Gyllensten, U.; Finnigan, J.P.; Alshawish, S.; Zhang, W.; Schadt, E.E.; Posner, M.R.; Genden, E.M.; et al. TGFβ receptor 1: An immune susceptibility gene in HPV-associated cancer. Cancer Res. 2014, 74, 6833–6844. [Google Scholar] [CrossRef] [Green Version]
- Lesseur, C.; Diergaarde, B.; Olshan, A.F.; Wünsch-Filho, V.; Ness, A.R.; Liu, G.; Lacko, M.; Eluf-Neto, J.; Franceschi, S.; Lagiou, P.; et al. Genome-wide association analyses identify new susceptibility loci for oral cavity and pharyngeal cancer. Nat. Genet. 2016, 48, 1544–1550. [Google Scholar] [CrossRef]
- Shete, S.; Liu, H.; Wang, J.; Yu, R.; Sturgis, E.M.; Li, G.; Dahlstrom, K.R.; Liu, Z.; Amos, C.I.; Wei, Q. A Genome-Wide Association Study Identifies Two Novel Susceptible Regions for Squamous Cell Carcinoma of the Head and Neck. Cancer Res. 2020, 80, 2451–2460. [Google Scholar] [CrossRef] [Green Version]
- Plant, K.; Fairfax, B.P.; Makino, S.; Vandiedonck, C.; Radhakrishnan, J.; Knight, J.C. Fine mapping genetic determinants of the highly variably expressed MHC gene ZFP57. Eur. J. Hum. Genet. 2014, 22, 568–571. [Google Scholar] [CrossRef] [Green Version]
- Viet, N.H.; Trung, N.Q.; Dong, L.T.; Trung, L.Q.; Espinoza, J.L. Genetic variants in NKG2D axis and susceptibility to Epstein-Barr virus-induced nasopharyngeal carcinoma. J. Cancer Res. Clin. Oncol. 2021, 147, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.V.; Toan, N.L.; Song, L.H.; Bock, C.T.; Kremsner, P.G.; Velavan, T.P. Hepatitis B virus-induced hepatocellular carcinoma: Functional roles of MICA variants. J. Viral. Hepat. 2013, 20, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Lesseur, C.; Ferreiro-Iglesias, A.; McKay, J.D.; Bossé, Y.; Johansson, M.; Gaborieau, V.; Landi, M.T.; Christiani, D.C.; Caporaso, N.C.; Bojesen, S.E.; et al. Genome-wide association meta-analysis identifies pleiotropic risk loci for aerodigestive squamous cell cancers. PLoS Genet. 2021, 17, e1009254. [Google Scholar] [CrossRef] [PubMed]
- Alemany, L.; Saunier, M.; Tinoco, L.; Quirós, B.; Alvarado-Cabrero, I.; Alejo, M.; Joura, E.A.; Maldonado, P.; Klaustermeier, J.; Salmerón, J.; et al. Large contribution of human papillomavirus in vaginal neoplastic lesions: A worldwide study in 597 samples. Eur. J. Cancer 2014, 50, 2846–2854. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Alemany, L.; Ordi, J.; Tous, S.; Alejo, M.; Bigby, S.M.; Joura, E.A.; Maldonado, P.; Laco, J.; Bravo, I.G.; et al. Worldwide human papillomavirus genotype attribution in over 2000 cases of intraepithelial and invasive lesions of the vulva. Eur. J. Cancer 2013, 49, 3450–3461. [Google Scholar] [CrossRef]
- Nicolás-Párraga, S.; Gandini, C.; Pimenoff, V.N.; Alemany, L.; de Sanjosé, S.; Xavier Bosch, F.; Bravo, I.G. HPV16 variants distribution in invasive cancers of the cervix, vulva, vagina, penis, and anus. Cancer Med. 2016, 5, 2909–2919. [Google Scholar] [CrossRef]
- Bertoli, H.K.; Thomsen, L.T.; Iftner, T.; Dehlendorff, C.; Kjær, S.K. Risk of vulvar, vaginal and anal high-grade intraepithelial neoplasia and cancer according to cervical human papillomavirus (HPV) status: A population-based prospective cohort study. Gynecol. Oncol. 2020, 157, 456–462. [Google Scholar] [CrossRef]
- Hardikar, S.; Johnson, L.G.; Malkki, M.; Petersdorf, E.W.; Galloway, D.A.; Schwartz, S.M.; Madeleine, M.M. A population-based case-control study of genetic variation in cytokine genes associated with risk of cervical and vulvar cancers. Gynecol. Oncol. 2015, 139, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Bodelon, C.; Madeleine, M.M.; Johnson, L.G.; Du, Q.; Malkki, M.; Petersdorf, E.W.; Schwartz, S.M. Genetic variation in CD83 and risks of cervical and vulvar cancers: A population-based case-control study. Gynecol. Oncol. 2012, 124, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Borecki, I.; Nguyen, L.; Ma, D.; Smith, K.; Huettner, P.C.; Mutch, D.G.; Herzog, T.J.; Gibb, R.K.; Powell, M.A.; et al. CD83 gene polymorphisms increase susceptibility to human invasive cervical cancer. Cancer Res. 2007, 67, 11202–11208. [Google Scholar] [CrossRef] [Green Version]
- Bodelon, C.; Madeleine, M.M.; Johnson, L.G.; Du, Q.; Galloway, D.A.; Malkki, M.; Petersdorf, E.W.; Schwartz, S.M. Genetic variation in the TLR and NF-κB pathways and cervical and vulvar cancer risk: A population-based case-control study. Int. J. Cancer 2014, 134, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.K.; Madeleine, M.M.; Johnson, L.G.; Du, Q.; Galloway, D.A.; Daling, J.R.; Malkki, M.; Petersdorf, E.W.; Schwartz, S.M. Nucleotide variation in IL-10 and IL-12 and their receptors and cervical and vulvar cancer risk: A hybrid case-parent triad and case-control study. Int. J. Cancer 2013, 133, 201–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chahoud, J.; Pickering, C.R.; Pettaway, C.A. Genetics and penile cancer: Recent developments and implications. Curr. Opin. Urol. 2019, 29, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Whibley, C.; Pharoah, P.D.; Hollstein, M. p53 polymorphisms: Cancer implications. Nat. Rev. Cancer 2009, 9, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Si, D.; Yao, Y.; Chen, X.; Qiu, J. Ethnicity-stratified analysis of the association between P53 rs1042522 polymorphism and women HPV infection: A meta-analysis. Microb. Pathog. 2021, 105099. [Google Scholar] [CrossRef] [PubMed]
- Klug, S.J.; Ressing, M.; Koenig, J.; Abba, M.C.; Agorastos, T.; Brenna, S.M.; Ciotti, M.; Das, B.R.; Del Mistro, A.; Dybikowska, A.; et al. TP53 codon 72 polymorphism and cervical cancer: A pooled analysis of individual data from 49 studies. Lancet Oncol. 2009, 10, 772–784. [Google Scholar] [CrossRef]
- Habbous, S.; Pang, V.; Eng, L.; Xu, W.; Kurtz, G.; Liu, F.F.; Mackay, H.; Amir, E.; Liu, G. p53 Arg72Pro polymorphism, HPV status and initiation, progression, and development of cervical cancer: A systematic review and meta-analysis. Clin. Cancer Res. 2012, 18, 6407–6415. [Google Scholar] [CrossRef] [Green Version]
- Stoehr, R.; Weisser, R.; Wendler, O.; Giedl, J.; Daifalla, K.; Gaisa, N.T.; Richter, G.; Campean, V.; Burger, M.; Wullich, B.; et al. Codon 72 Polymorphism and Risk for Squamous Cell Carcinoma of the Penis: A Caucasian Case-Control Study. J. Cancer 2018, 9, 4234–4241. [Google Scholar] [CrossRef] [Green Version]
- Humbey, O.; Cairey-Remonnay, S.; Guérrini, J.S.; Algros, M.P.; Mougin, C.; Bittard, H.; Aubin, F. Detection of the human papillomavirus and analysis of the TP53 polymorphism of exon 4 at codon 72 in penile squamous cell carcinomas. Eur. J. Cancer 2003, 39, 684–690. [Google Scholar] [CrossRef]
- Tornesello, M.L.; Duraturo, M.L.; Guida, V.; Losito, S.; Botti, G.; Pilotti, S.; Stefanon, B.; De Palo, G.; Buonaguro, L.; Buonaguro, F.M. Analysis of TP53 codon 72 polymorphism in HPV-positive and HPV-negative penile carcinoma. Cancer Lett. 2008, 269, 159–164. [Google Scholar] [CrossRef]
- Rosenthal, A.N.; Ryan, A.; Hopster, D.; Jacobs, I.J. p53 codon 72 polymorphism in vulval cancer and vulval intraepithelial neoplasia. Br. J. Cancer 2000, 83, 1287–1290. [Google Scholar] [CrossRef] [Green Version]
- Shim, T.N.; Harwood, C.A.; Marsh, S.G.; Gotch, F.M.; Quint, W.; de Koning, M.N.; Francis, N.; Jameson, C.; Freeman, A.; Minhas, S.; et al. The prevalence of human leucocyte antigen and human papillomavirus DNA in penile intraepithelial neoplasia in England 2011–2012. Int. J. STD AIDS 2021, 32, 388–395. [Google Scholar] [CrossRef]
- Espinoza, J.L.; Nguyen, V.H.; Ichimura, H.; Pham, T.T.; Nguyen, C.H.; Pham, T.V.; Elbadry, M.I.; Yoshioka, K.; Tanaka, J.; Trung, L.Q.; et al. A functional polymorphism in the NKG2D gene modulates NK-cell cytotoxicity and is associated with susceptibility to Human Papilloma Virus-related cancers. Sci. Rep. 2016, 6, 39231. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, J.L.; Takami, A.; Yoshioka, K.; Nakata, K.; Sato, T.; Kasahara, Y.; Nakao, S. Human microRNA-1245 downregulates the NKG2D receptor in NK cells and impairs NKG2D-mediated functions. Haematologica 2012, 97, 1295–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, N.; Tan, Y.X.; Joncker, N.T.; Choy, A.; Gallardo, F.; Xiong, N.; Knoblaugh, S.; Cado, D.; Greenberg, N.M.; Greenberg, N.R.; et al. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy. Immunity 2008, 28, 571–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, H.; Matsuo, K.; Kumimoto, H.; Hiraki, A.; Suzuki, T.; Yatabe, Y.; Komori, K.; Kanemitsu, Y.; Hirai, T.; Kato, T.; et al. Decreased risk of colorectal cancer with the high natural killer cell activity NKG2D genotype in Japanese. Carcinogenesis 2008, 29, 316–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Imai, K.; Morishita, Y.; Hayashi, I.; Kusunoki, Y.; Nakachi, K. Identification of the NKG2D haplotypes associated with natural cytotoxic activity of peripheral blood lymphocytes and cancer immunosurveillance. Cancer Res. 2006, 66, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.M.O.; Peixoto da Silva, S.; Costa, N.R.; Gil da Costa, R.M.; Medeiros, R. The Role of MicroRNAs in the Metastatic Process of High-Risk HPV-Induced Cancers. Cancers 2018, 10, 493. [Google Scholar] [CrossRef] [Green Version]
- Cicchini, L.; Blumhagen, R.Z.; Westrich, J.A.; Myers, M.E.; Warren, C.J.; Siska, C.; Raben, D.; Kechris, K.J.; Pyeon, D. High-Risk Human Papillomavirus E7 Alters Host DNA Methylome and Represses HLA-E Expression in Human Keratinocytes. Sci. Rep. 2017, 7, 3633. [Google Scholar] [CrossRef] [Green Version]
- Dehlendorff, C.; Baandrup, L.; Kjaer, S.K. Real-World Effectiveness of Human Papillomavirus Vaccination Against Vulvovaginal High-Grade Precancerous Lesions and Cancers. J. Natl. Cancer Inst. 2021, 113, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Selk, A.; Garland, S.M.; Bogliatto, F.; Kyrgiou, M.; Weyers, S.; Arbyn, M. Prophylactic vaccination against human papillomaviruses to prevent vulval and vaginal cancer and their precursors. Expert Rev. Vaccines 2019, 18, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Arbyn, M.; Xu, L. Efficacy and safety of prophylactic HPV vaccines. A Cochrane review of randomized trials. Expert Rev. Vaccines 2018, 17, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Haralambieva, I.H.; Ovsyannikova, I.G.; Pankratz, V.S.; Kennedy, R.B.; Jacobson, R.M.; Poland, G.A. The genetic basis for interindividual immune response variation to measles vaccine: New understanding and new vaccine approaches. Expert Rev. Vaccines 2013, 12, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambuso, R.S.; Rebello, G.; Kaambo, E. Personalized Human Papillomavirus Vaccination for Persistence of Immunity for Cervical Cancer Prevention: A Critical Review with Experts’ Opinions. Front. Oncol. 2020, 10, 548. [Google Scholar] [CrossRef]


{kind=link}
| 1st Author, Year, Reference | Discovery Cohort | Confirmatory Cohort | Ethnicity | Number of SNPs Tested | Main Findings |
|---|---|---|---|---|---|
| Bowden S. Lancet Oncol. 2021 [14]. | 273,377 women (4769 CIN3 and invasive cervical cancer case samples and 145,545 control) | 128,123 women (1648 invasive cervical cancer cases) from a Finnish dataset | Discovery cohort: UK (UK Biobank) Confirmatory cohort: Finland (FinnGen). | 9 million SNPs | Six loci strongly associated with CIN3 and invasive cervical cancer. Two novel SNPs, rs10175462 (PAX8) and rs27069 (CLPTM1L) Verified previously reported SNPs: rs9272050 (HLA-DQA1), rs6938453 (MICA), rs55986091 (HLA-DQB1) and rs9266183 (HLA-B). Three SNPs, rs10175462 (PAX8 gene), rs27069 CLPTM1L) and rs9272050 (HLA-DQA1) were replicated in the confirmatory Finnish dataset. |
| Takeuchi F. Hum Mol Genet. 2019 [79] | 2690 cases and 4712 controls (1289 cases and 2718 controls from previous Chinese study) | 1320 cases and 1994 controls from newly reported Japanese study | Japanese GWAS study: East Asian populations | 4,602,429 SNPs |
|
| Chen D. J. Natl. Cancer Inst. 2013 [18] | GWAS in women of European-ancestry with 1075 cervical cancer cases, and 4014 controls | replicated in 1140 case subjects and 1058 control subjects | European descendants | 731,422 SNPs |
|
| Leo PJ. PLoS Genet. 2017 [41] | 2866 cervical cancer cases and 6481 controls | Nd | European descendants | 10,863,230 SNPs |
|
| Chen D. Oncotarget. 2016 [78] | analysis of data from GWAS of women of European ancestry (1075 cervical cancer cases and 4014 controls + replicated in 1140 case subjects and 1058 control subjects) | 961 patients (827 with CIN3 and 123 with cervical cancer) and 1725 controls | European | 5,471,179 SNPs |
|
| Shi Y. Nat. Genet. 2013 [79] | 1364 women with cervical cancer and 3028 female controls | Dataset 1: 1824 cases and 3808 controls. Dataset 2: 2343 cases and 3388 controls | Han Chinese population | 563,339 SNPs |
|
| Miura K. J. Med Virology. 2014 [84] | 226 women with cervical cancer and 186 controls | Not included | Japanese | 556,045 SNPs |
|
| 1st Author, Year, Reference | No. of Cases | No. of Controls | Ethnicity | No. of SNPs Tested | Gene Variants | Main Findings |
|---|---|---|---|---|---|---|
| Mckay JD, Plos Genet. 2011 [95] | 2091 (oral cavity, pharynx, larynx and esophagus) | 3513 | European | nd | rs1494961, rs4767364 |
|
| Liang C. Head Neck, 2012 [96] | 545 (HNSCC) | 676 | none specified, conducted in Greater Boston, Massachusetts | nd | rs4767364, rs1494961 |
|
| Levovitz C. Cancer Res. 2014 [97] | 2091 UADT cancer patients (HNSCC) | 8334 | European | 296,728 | variants in 14 immune-related genes, TGFβR1 |
|
| Lesseur C. Nat Genet. 2016 [98] | 6034 (oral cavity, pharyngeal cancer) | 6585 | Europe, North America and South America | nd | rs3828805, rs201982221, rs1453414, rs6547741, rs928674, rs8181047, rs10462706 |
|
| Shete S. Cancer Res. 2020 [99] | 2171 GWAS, 5205 cohort | 4493 GWAS, 3232 cohort | European | nd | rs1265081, rs3135001, rs13211972, rs259919, rs142021700, |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza, H.; Ha, K.T.; Pham, T.T.; Espinoza, J.L. Genetic Predisposition to Persistent Human Papillomavirus-Infection and Virus-Induced Cancers. Microorganisms 2021, 9, 2092. https://doi.org/10.3390/microorganisms9102092
Espinoza H, Ha KT, Pham TT, Espinoza JL. Genetic Predisposition to Persistent Human Papillomavirus-Infection and Virus-Induced Cancers. Microorganisms. 2021; 9(10):2092. https://doi.org/10.3390/microorganisms9102092
Chicago/Turabian StyleEspinoza, Helen, Kim T. Ha, Trang T. Pham, and J. Luis Espinoza. 2021. "Genetic Predisposition to Persistent Human Papillomavirus-Infection and Virus-Induced Cancers" Microorganisms 9, no. 10: 2092. https://doi.org/10.3390/microorganisms9102092
APA StyleEspinoza, H., Ha, K. T., Pham, T. T., & Espinoza, J. L. (2021). Genetic Predisposition to Persistent Human Papillomavirus-Infection and Virus-Induced Cancers. Microorganisms, 9(10), 2092. https://doi.org/10.3390/microorganisms9102092