
Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
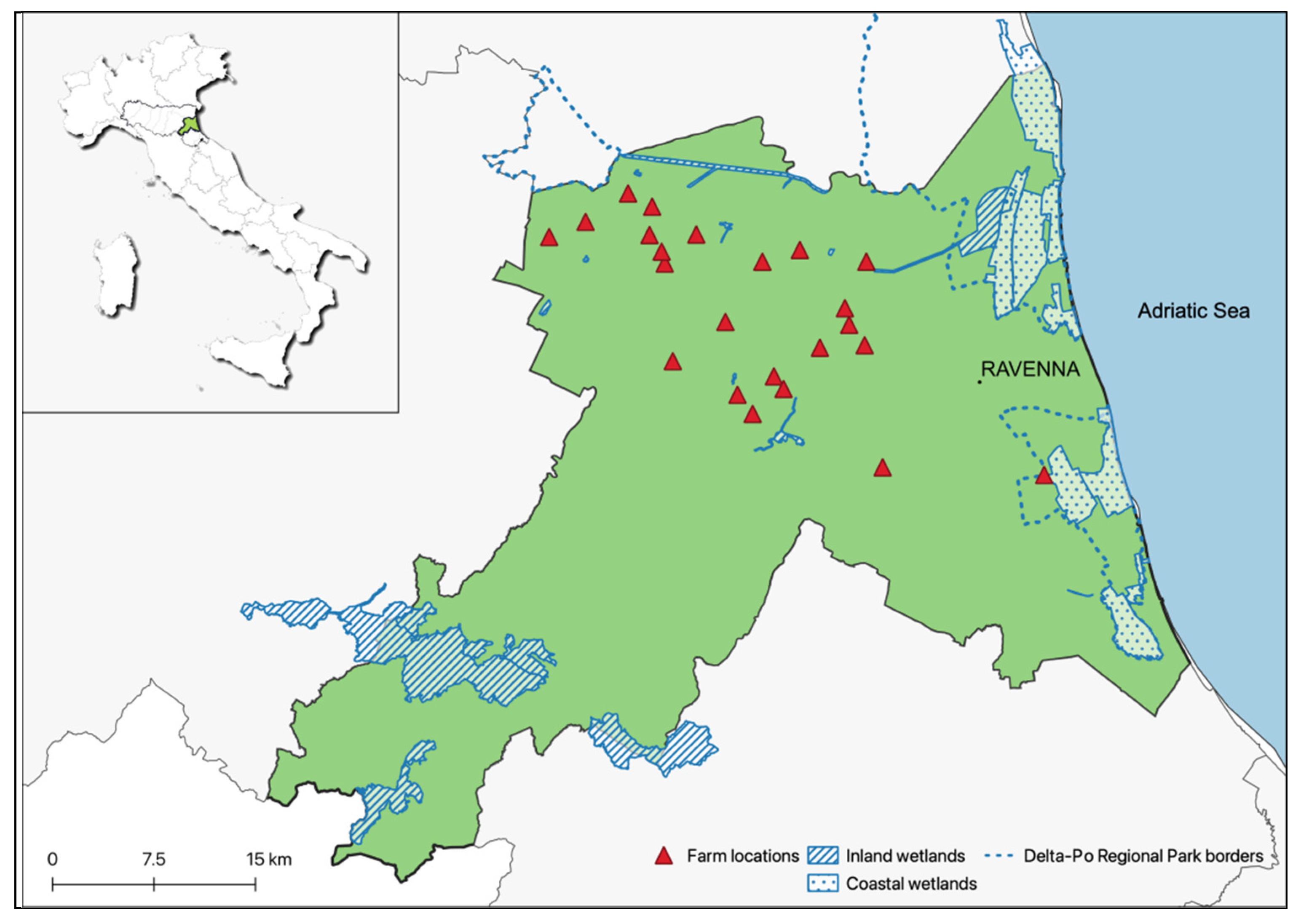
2.1. Study Area and Bird Farming System
2.2. Sample Collection in Birds and Humans
2.3. Animal and Human Rights Statement
2.4. Experimental Design and Laboratory Tests in Birds
2.5. Experimental Design and Laboratory Tests in Humans
2.6. Statistical Methods
3. Results
3.1. Bird and Human Study Populations
3.2. Virological and Serological Results from Birds
3.2.1. Avian Influenza Virus Detection, Isolation, and Characterization
3.2.2. Serological Analyses
3.3. Serological Results from Humans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Webster, R.G.; Govorkova, E.A. Continuing challenges in influenza. Ann. N. Y. Acad. Sci. 2014, 1323, 115–139. [Google Scholar] [CrossRef]
- Neumann, G.; Kawaoka, Y. Transmission of influenza A viruses. Virology 2015, 479–480, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Freidl, G.S.; Meijer, A.; de Bruin, E.; de Nardi, M.; Munoz, O.; Capua, I.; Breed, A.C.; Harris, K.; Hill, A.; Kosmider, R.; et al. FLURISK Consortium. Influenza at the animal-human interface: A review of the literature for virological evidence of human infection with swine or avian influenza viruses other than A(H5N1). Eurosurveillance 2014, 19, 20793. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.C. The Pandemic Threat of Emerging H5 and H7 Avian Influenza Viruses. Viruses 2018, 10, 461. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic Potential of Influenza A Viruses: A Comprehensive Overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control. First Identification of Human Cases of Avian Influenza A (H5N8) Infection; ECDC: Stockholm, Sweden, 2021.
- Wang, Y.; Niu, S.; Zhang, B.; Yang, C.; Zhou, Z. The whole genome analysis for the first human infection with H10N3 influenza virus in China. J. Infect. 2021. [Google Scholar] [CrossRef]
- Sikkema, R.S.; Freidl, G.S.; de Bruin, E.; Koopmans, M. Weighing serological evidence of human exposure to animal influenza viruses—A literature review. Eurosurveillance 2016, 21, 30388. [Google Scholar] [CrossRef] [Green Version]
- Puzelli, S.; Di Trani, L.; Fabiani, C.; Campitelli, L.; De Marco, M.A.; Capua, I.; Aguilera, J.F.; Zambon, M.; Donatelli, I. Serological analysis of serum samples from humans exposed to avian H7 influenza viruses in Italy between 1999 and 2003. J. Infect. Dis. 2005, 192, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Di Trani, L.; Porru, S.; Bonfanti, L.; Cordioli, P.; Cesana, B.M.; Boni, A.; Di Carlo, A.S.; Arici, C.; Donatelli, I.; Tomao, P.; et al. Serosurvey against H5 and H7 avian influenza viruses in Italian poultry workers. Avian Dis. 2012, 56, 1068–1071. [Google Scholar] [CrossRef]
- Puzelli, S.; Rossini, G.; Facchini, M.; Vaccari, G.; Di Trani, L.; Di Martino, A.; Gaibani, P.; Vocale, C.; Cattoli, G.; Bennett, M.; et al. Influenza Task Force. Human infection with highly pathogenic A(H7N7) avian influenza virus, Italy, 2013. Emerg. Infect. Dis. 2014, 20, 1745–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puzelli, S.; Rizzo, C.; Fabiani, C.; Facchini, M.; Gaibani, P.; Landini, M.P.; Gagliotti, C.; Moro, M.L.; Rangoni, R.; Piccolomini, L.L.; et al. Influenza A(H7N7) Virus among Poultry Workers, Italy, 2013. Emerg. Infect. Dis. 2016, 22, 1512–1513. [Google Scholar] [CrossRef] [Green Version]
- Van Kerkhove, M.D.; Mumford, E.; Mounts, A.W.; Bresee, J.; Ly, S.; Bridges, C.B.; Otte, J. Highly pathogenic avian influenza (H5N1): Pathways of exposure at the animal-human interface, a systematic review. PLoS ONE 2011, 6, e14582, Erratum in 2012, 7. [Google Scholar] [CrossRef]
- Bahl, J.; Pham, T.T.; Hill, N.J.; Hussein, I.T.; Ma, E.J.; Easterday, B.C.; Halpin, R.A.; Stockwell, T.B.; Wentworth, D.E.; Kayali, G.; et al. Ecosystem Interactions Underlie the Spread of Avian Influenza A Viruses with Pandemic Potential. PLoS Pathog. 2016, 12, e1005620. [Google Scholar] [CrossRef]
- Halpin, J. Avian flu from an occupational health perspective. Arch. Environ. Occup. Health 2005, 60, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E. Occupational and consumer risks from avian influenza viruses. Dev. Biol. 2006, 124, 85–90. [Google Scholar]
- Shafir, S.C.; Fuller, T.; Smith, T.B.; Rimoin, A.W. A national study of individuals who handle migratory birds for evidence of avian and swine-origin influenza virus infections. J. Clin. Virol. 2012, 54, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Centro Emiliano Romagnolo di Epidemiologia Veterinaria—IZSLER “Bruno Ubertini”. Regional Plan for Permanent Monitoring of Avian Influenza. Results of Controls Year. 2006. Available online: https://www.izsler.it/izs_bs/ftp//doc/Influenza%20Aviare%20x%20ftp/Piano%20monitoraggio%20dati2006.pdf (accessed on 28 July 2021).
- Council of European Union. Council Directive 92/40/EEC of 19 May 1992 Introducing Community Measures for the Control of Avian Influenza. Off. J. L 1992, 167, 1–16. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A31992L0040 (accessed on 26 July 2021).
- World Health Organization. WHO Manual on Animal Influenza Diagnosis and Surveillance. 2002. Available online: https://apps.who.int/iris/bitstream/handle/10665/68026/WHO_CDS_CSR_NCS_2002.5.pdf?sequence=1&isAllowed=y (accessed on 26 July 2021).
- Siebinga, J.T.; de Boer, G.F. Influenza A viral nucleoprotein detection in isolates from human and various animal species. Arch. Virol. 1988, 100, 75–87. [Google Scholar] [CrossRef]
- Delogu, M.; Piredda, I.; Pintore, A.; Cabras, P.; Cotti, C.; Ghetti, G.; Raffini, E.; De Marco, M.A. Avian influenza and animal health risk: Conservation of endemic threatened wild birds in Sardinia Island. Avian Dis. 2012, 56 (Suppl. 4), 1034–1039. [Google Scholar] [CrossRef]
- Stephenson, I.; Wood, J.M.; Nicholson, K.G.; Charlett, A.; Zambon, M.C. Detection of anti-H5 responses in human sera by HI using horse erythrocytes following MF59-adjuvanted influenza A/Duck/Singapore/97 vaccine. Virus Res. 2004, 103, 91–95. [Google Scholar] [CrossRef]
- Meijer, A.; Bosman, A.; van de Kamp, E.E.; Wilbrink, B.; van Beest Holle, M.D.R.; Koopmans, M. Measurement of antibodies to avian influenza virus A(H7N7) in humans by hemagglutination inhibition test. J. Virol. Methods 2006, 132, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Kayali, G.; Setterquist, S.F.; Capuano, A.W.; Myers, K.P.; Gill, J.S.; Gray, G.C. Testing human sera for antibodies against avian influenza viruses: Horse RBC hemagglutination inhibition vs. microneutralization assays. J. Clin. Virol. 2008, 43, 73–78. [Google Scholar] [CrossRef] [Green Version]
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2021. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.04_AI.pdf (accessed on 26 July 2021).
- World Health Organization. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization: Geneva, Switzerland, 2011; Available online: https://apps.who.int/iris/handle/10665/44518 (accessed on 28 July 2021).
- Rowe, T.; Abernathy, R.A.; Hu-Primmer, J.; Thompson, W.W.; Lu, X.; Lim, W.; Fukuda, K.; Cox, N.J.; Katz, J.M. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J. Clin. Microbiol. 1999, 37, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Myers, K.P.; Setterquist, S.F.; Capuano, A.W.; Gray, G.C. Infection due to 3 avian influenza subtypes in United States veterinarians. Clin. Infect. Dis. 2007, 45, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.C.; McCarthy, T.; Capuano, A.W.; Setterquist, S.F.; Alavanja, M.C.; Lynch, C.F. Evidence for avian influenza A infections among Iowa’s agricultural workers. Influenza Other Respir. Viruses 2008, 2, 61–69. [Google Scholar] [CrossRef]
- Kayali, G.; Ortiz, E.J.; Chorazy, M.L.; Gray, G.C. Evidence of previous avian influenza infection among US turkey workers. Zoonoses Public Health 2010, 57, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Kayali, G.; Barbour, E.; Dbaibo, G.; Tabet, C.; Saade, M.; Shaib, H.A.; Debeauchamp, J.; Webby, R.J. Evidence of infection with H4 and H11 avian influenza viruses among Lebanese chicken growers. PLoS ONE 2011, 6, e26818. [Google Scholar] [CrossRef]
- Brown, I.; Kuiken, T.; Mulatti, P.; Smietanka, K.; Staubach, C.; Stroud, D.; Therkildsen, O.R.; Willeberg, P.; Baldinelli, F.; Verdonck, F.; et al. Scientific report: Avian influenza overview September–November 2017. EFSA J. 2017, 15, e05141. [Google Scholar]
- Terregino, C.; De Nardi, R.; Guberti, V.; Scremin, M.; Raffini, E.; Martin, A.M.; Cattoli, G.; Bonfanti, L.; Capua, I. Active surveillance for avian influenza viruses in wild birds and backyard flocks in Northern Italy during 2004 to 2006. Avian Pathol. 2007, 36, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, M.A.; Foni, E.; Campitelli, L.; Delogu, M.; Raffini, E.; Chiapponi, C.; Barigazzi, G.; Cordioli, P.; Di Tran, L.; Donatelli, I. Influenza virus circulation in wild aquatic birds in Italy during H5N2 and H7N1 poultry epidemic periods (1998 to 2000). Avian Pathol. 2005, 34, 480–485. [Google Scholar] [CrossRef]
- Capua, I.; Kajaste-Rudnitski, A.; Bertoli, E.; Vicenzi, E. Pandemic vaccine preparedness--have we left something behind? PLoS Pathog. 2009, 5, e1000482. [Google Scholar] [CrossRef] [Green Version]
- Babu, T.M.; Perera, R.A.P.M.; Wu, J.T.; Fitzgerald, T.; Nolan, C.; Cowling, B.J.; Krauss, S.; Treanor, J.J.; Peiris, M. Population Serologic Immunity to Human and Avian H2N2 Viruses in the United States and Hong Kong for Pandemic Risk Assessment. J. Infect. Dis. 2018, 218, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Beare, A.S.; Webster, R.G. Replication of avian influenza viruses in humans. Arch. Virol. 1991, 119, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.H.; Yang, J.R.; Wu, H.S.; Chang, M.C.; Lin, J.S.; Lin, C.Y.; Liu, Y.L.; Lo, Y.C.; Yang, C.H.; Chuang, J.H.; et al. Human infection with avian influenza A H6N1 virus: An epidemiological analysis. Lancet Respir. Med. 2013, 1, 771–778. [Google Scholar] [CrossRef]
- Okoye, J.O.; Eze, D.C.; Krueger, W.S.; Heil, G.L.; White, S.K.; Merrill, H.R.; Gray, G.C. Evidence for subclinical H5N1 avian influenza infections among Nigerian poultry workers. J. Med. Virol. 2014, 86, 2070–2075. [Google Scholar] [CrossRef]
- Khuntirat, B.P.; Yoon, I.K.; Blair, P.J.; Krueger, W.S.; Chittaganpitch, M.; Putnam, S.D.; Supawat, K.; Gibbons, R.V.; Pattamadilok, S.; Sawanpanyalert, P.; et al. Evidence for subclinical avian influenza virus infections among rural Thai villagers. Clin. Infect. Dis. 2011, 53, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Khurelbaatar, N.; Krueger, W.S.; Heil, G.L.; Darmaa, B.; Ulziimaa, D.; Tserennorov, D.; Baterdene, A.; Anderson, B.D.; Gray, G.C. Sparse evidence for equine or avian influenza virus infections among Mongolian adults with animal exposures. Influenza Other Respir. Viruses 2013, 7, 1246–1250. [Google Scholar] [CrossRef] [Green Version]
- Khurelbaatar, N.; Krueger, W.S.; Heil, G.L.; Darmaa, B.; Ulziimaa, D.; Tserennorov, D.; Baterdene, A.; Anderson, B.D.; Gray, G.C. Little evidence of avian or equine influenza virus infection among a cohort of Mongolian adults with animal exposures, 2010–2011. PLoS ONE 2014, 21, e85616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.Y.; Yang, J.R.; Lin, Y.J.; Yang, C.H.; Cheng, M.C.; Liu, M.T.; Wu, H.S.; Chang, F.Y. Serological comparison of antibodies to avian influenza viruses, subtypes H5N2, H6N1, H7N3 and H7N9 between poultry workers and non-poultry workers in Taiwan in 2012. Epidemiol. Infect. 2015, 143, 2965–2974. [Google Scholar] [CrossRef]
- Coman, A.; Maftei, D.N.; Krueger, W.S.; Heil, G.L.; Chereches, R.M.; Sirlincan, E.; Bria, P.; Dragnea, C.; Kasler, I.; Valentine, M.A.; et al. A prospective study of Romanian agriculture workers for zoonotic influenza infections. PLoS ONE 2014, 9, e98248. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Qin, K. Human-infecting influenza A (H9N2) virus: A forgotten potential pandemic strain? Zoonoses Public Health 2020, 67, 203–212. [Google Scholar] [CrossRef]
- Barbour, E.K.; Sagherian, V.K.; Sagherian, N.K.; Dankar, S.K.; Jaber, L.S.; Usayran, N.N.; Farran, M.T. Avian influenza outbreak in poultry in the Lebanon and transmission to neighbouring farmers and swine. Vet. Ital. 2006, 42, 77–85. [Google Scholar]
- Heidari, A.; Mancin, M.; Nili, H.; Pourghanbari, G.H.; Lankarani, K.B.; Leardini, S.; Cattoli, G.; Monne, I.; Piccirillo, A. Serological evidence of H9N2 avian influenza virus exposure among poultry workers from Fars province of Iran. Virol. J. 2016, 27, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawar, S.D.; Tandale, B.V.; Raut, C.G.; Parkhi, S.S.; Barde, T.D.; Gurav, Y.K.; Kode, S.S.; Mishra, A.C. Avian influenza H9N2 seroprevalence among poultry workers in Pune, India, 2010. PLoS ONE 2012, 7, e36374. [Google Scholar] [CrossRef] [PubMed]
- Ahad, A.; Thornton, R.N.; Rabbani, M.; Yaqub, T.; Younus, M.; Muhammad, K.; Mahmood, A.; Shabbir, M.Z.; Kashem, M.A.; Islam, M.Z.; et al. Risk factors for H7 and H9 infection in commercial poultry farm workers in provinces within Pakistan. Prev. Vet. Med. 2014, 117, 610–614. [Google Scholar] [CrossRef]
- Chaudhry, M.; Webby, R.; Swayne, D.; Rashid, H.B.; DeBeauchamp, J.; Killmaster, L.; Criado, M.F.; Lee, D.H.; Webb, A.; Yousaf, S.; et al. Avian influenza at animal-human interface: One-health challenge in live poultry retail stalls of Chakwal, Pakistan. Influenza Other Respir. Viruses 2020, 14, 257–265. [Google Scholar] [CrossRef]
- Lu, C.Y.; Lu, J.H.; Chen, W.Q.; Jiang, L.F.; Tan, B.Y.; Ling, W.H.; Zheng, B.J.; Sui, H.Y. Potential infections of H5N1 and H9N2 avian influenza do exist in Guangdong populations of China. Chin. Med. J. 2008, 121, 2050–2053. [Google Scholar] [CrossRef]
- Jia, N.; de Vlas, S.J.; Liu, Y.X.; Zhang, J.S.; Zhan, L.; Dang, R.L.; Ma, Y.H.; Wang, X.J.; Liu, T.; Yang, G.P.; et al. Serological reports of human infections of H7 and H9 avian influenza viruses in northern China. J. Clin. Virol. 2009, 44, 225–229. [Google Scholar] [CrossRef]
- Wang, Q.; Ju, L.; Liu, P.; Zhou, J.; Lv, X.; Li, L.; Shen, H.; Su, H.; Jiang, L.; Jiang, Q. Serological and virological surveillance of avian influenza A virus H9N2 subtype in humans and poultry in Shanghai, China, between 2008 and 2010. Zoonoses Public Health 2015, 62, 131–140. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, L.; Pu, J.; Zhao, J.; Sun, Y.; Shen, G.; Wei, H.; Zhu, J.; Zheng, R.; Xiong, D.; et al. Risk perceptions for avian influenza virus infection among poultry workers, China. Emerg. Infect. Dis. 2013, 19, 313–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Tian, B.; Jianfang, Z.; Yongkun, C.; Xiaodan, L.; Wenfei, Z.; Yan, L.; Jing, T.; Junfeng, G.; Tao, C.; et al. A comprehensive retrospective study of the seroprevalence of H9N2 avian influenza viruses in occupationally exposed populations in China. PLoS ONE 2017, 12, e0178328. [Google Scholar] [CrossRef]
- Yang, P.; Ma, C.; Shi, W.; Cui, S.; Lu, G.; Peng, X.; Zhang, D.; Liu, Y.; Liang, H.; Zhang, Y.; et al. A serological survey of antibodies to H5, H7 and H9 avian influenza viruses amongst the duck-related workers in Beijing, China. PLoS ONE 2012, 7, e50770. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Wang, A.R.; Liu, Z.H.; Liang, W.; Li, X.X.; Tang, Y.J.; Miao, Z.M.; Chai, T.J. Seroprevalence of avian influenza H9N2 among poultry workers in Shandong Province, China. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1347–1351. [Google Scholar] [CrossRef]
- de Bruin, E.; Zhang, X.; Ke, C.; Sikkema, R.; Koopmans, M. Serological evidence for exposure to avian influenza viruses within poultry workers in southern China. Zoonoses Public Health 2017, 64, e51–e59. [Google Scholar] [CrossRef]
- Li, S.; Zhou, Y.; Song, W.; Pang, Q.; Miao, Z. Avian influenza virus H9N2 seroprevalence and risk factors for infection in occupational poultry-exposed workers in Tai’an of China. J. Med. Virol. 2016, 88, 1453–1456. [Google Scholar] [CrossRef] [PubMed]
- Horm, S.V.; Tarantola, A.; Rith, S.; Ly, S.; Gambaretti, J.; Duong, V.Y.P.; Sorn, S.; Holl, D.; Allal, L.; Kalpravidh, W.; et al. Intense circulation of A/H5N1 and other avian influenza viruses in Cambodian live-bird markets with serological evidence of sub-clinical human infections. Emerg. Microbes Infect. 2016, 5, e70. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.; Hung, I.F.; Lui, Y.M.; Mok, F.K.; Chan, A.S.; Li, P.T.; Wong, T.L.; Ho, D.T.; Chan, J.F.; Chan, K.H.; et al. Ongoing transmission of avian influenza A viruses in Hong Kong despite very comprehensive poultry control measures: A prospective seroepidemiology study. J. Infect. 2016, 72, 207–213. [Google Scholar] [CrossRef]
- Li, S.; Zhou, Y.; Zhao, Y.; Li, W.; Song, W.; Miao, Z. Avian influenza H9N2 seroprevalence among pig population and pig farm staff in Shandong, China. Virol. J. 2015, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Cui, S.; Sun, Y.; Zhao, J.; Zhang, D.; Zhang, L.; Zhang, Y.; Pan, Y.; Wu, S.; Duan, W.; et al. Avian influenza A (H9N2) virus infections among poultry workers, swine workers, and the general population in Beijing, China, 2013–2016: A serological cohort study. Influenza Other Respir. Viruses 2019, 13, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.J.; Zhao, T.; Chen, S.H.; Xia, X.; Yang, X.X.; Wang, G.L.; Fang, L.Q.; Ma, G.Y.; Wu, M.N.; Qian, Y.H.; et al. Avian Influenza A Virus Infection among Workers at Live Poultry Markets, China, 2013–2016. Emerg. Infect. Dis. 2018, 24, 1246–1256. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Nguyen, D.C.; Rowe, T.; Lu, X.; Hu-Primmer, J.; Huynh, L.P.; Hang, N.L.; Katz, J.M. Seroprevalence of antibodies to avian influenza A (H5) and A (H9) viruses among market poultry workers, Hanoi, Vietnam, 2001. PLoS ONE 2012, 7, e43948. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, M.R.; Kayed, A.S.; Elabd, M.A.; Zeid, D.A.; Zaki, S.A.; El Rifay, A.S.; Sherif, L.S.; McKenzie, P.P.; Webster, R.G.; Webby, R.J.; et al. Avian influenza A(H5N1) and A(H9N2) seroprevalence and risk factors for infection among Egyptians: A prospective, controlled seroepidemiological study. J. Infect. Dis. 2015, 211, 1399–1407. [Google Scholar] [CrossRef] [Green Version]
- Gray, G.C.; Ferguson, D.D.; Lowther, P.E.; Heil, G.L.; Friary, J.A. A national study of US bird banders for evidence of avian influenza virus infections. J. Clin. Virol. 2011, 51, 132–135. [Google Scholar] [CrossRef] [Green Version]
- Coman, A.; Maftei, D.N.; Krueger, W.S.; Heil, G.L.; Friary, J.A.; Chereches, R.M.; Sirlincan, E.; Bria, P.; Dragnea, C.; Kasler, I.; et al. Serological evidence for avian H9N2 influenza virus infections among Romanian agriculture workers. J. Infect. Public Health 2013, 6, 438–447, Erratum in: J. Infect. Public Health 2018, 11, 902. [Google Scholar] [CrossRef] [Green Version]
- Simulundu, E.; Nao, N.; Yabe, J.; Muto, N.A.; Sithebe, T.; Sawa, H.; Manzoor, R.; Kajihara, M.; Muramatsu, M.; Ishii, A.; et al. The zoonotic potential of avian influenza viruses isolated from wild waterfowl in Zambia. Arch. Virol. 2014, 159, 2633–2640. [Google Scholar] [CrossRef] [Green Version]
- Gill, J.S.; Webby, R.; Gilchrist, M.J.; Gray, G.C. Avian influenza among waterfowl hunters and wildlife professionals. Emerg. Infect. Dis. 2006, 12, 1284–1286. [Google Scholar] [CrossRef]
- De Marco, M.A.; Campitelli, L.; Foni, E.; Raffini, E.; Barigazzi, G.; Delogu, M.; Guberti, V.; Di Trani, L.; Tollis, M.; Donatelli, I. Influenza surveillance in birds in Italian wetlands (1992–1998): Is there a host restricted circulation of influenza viruses in sympatric ducks and coots? Vet. Microbiol. 2004, 98, 197–208. [Google Scholar] [CrossRef]
- Krammer, F. The human antibody response to influenza A virus infection and vaccination. Nat. Rev. Immunol. 2019, 19, 383–397. [Google Scholar] [CrossRef]
- Kitphati, R.; Pooruk, P.; Lerdsamran, H.; Poosuwan, S.; Louisirirotchanakul, S.; Auewarakul, P.; Chokphaibulkit, K.; Noisumdaeng, P.; Sawanpanyalert, P.; Puthavathana, P. Kinetics and longevity of antibody response to influenza A H5N1 virus infection in humans. Clin. Vaccine Immunol. 2009, 16, 978–981. [Google Scholar] [CrossRef] [Green Version]
- Lindskog, C.; Ellström, P.; Olsen, B.; Pontén, F.; van Riel, D.; Munster, V.J.; González-Acuña, D.; Kuiken, T.; Jourdain, E. European H16N3 gull influenza virus attaches to the human respiratory tract and eye. PLoS ONE 2013, 8, e60757. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Tian, J.; Zhang, Y.; Bai, X.; Wang, X.; Shi, J.; Wang, Y.; Ma, L.; Yang, C.; et al. Characteristics of the first H16N3 subtype influenza A viruses isolated in western China. Transbound Emerg. Dis. 2020, 67, 1677–1687, Erratum in 2020, 67, 3075.. [Google Scholar] [CrossRef]
- Qi, L.; Pujanauski, L.M.; Davis, A.S.; Schwartzman, L.M.; Chertow, D.S.; Baxter, D.; Scherler, K.; Hartshorn, K.L.; Slemons, R.D.; Walters, K.A.; et al. Contemporary avian influenza A virus subtype H1, H6, H7, H10, and H15 hemagglutinin genes encode a mammalian virulence factor similar to the 1918 pandemic virus H1 hemagglutinin. mBio 2014, 5, e02116. [Google Scholar] [CrossRef] [Green Version]
- Tzarum, N.; McBride, R.; Nycholat, C.M.; Peng, W.; Paulson, J.C.; Wilson, I.A. Unique Structural Features of Influenza Virus H15 Hemagglutinin. J. Virol. 2017, 91, e00046-17. [Google Scholar] [CrossRef] [Green Version]
- Cannon, R.M.; Roe, R.T. Livestock Disease Surveys: A Field Manual for Veterinarians; Australian Bureau of Animal Health; Australian Government Publishing Service: Canberra, Australia, 1982; pp. 1–35. Available online: https://www.aphis.usda.gov/animal_health/surveillance_toolbox/docs/epi_surv/cannon_roe_1982_livestock_disease_surveys.pdf (accessed on 4 October 2021).
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Farm ID Number | No. of Locations † | Husbrandry/Commercial Area (Type) | Species Reared | No. of Birds |
|---|---|---|---|---|
| 1 | 1 | F-RW (breeding) | Mallards, geese | 2000 |
| 2 | 1 | L-BGB (breeding) | Pheasants | 1600–12,000 |
| 3 | 3 | F-RW (breeding) | Ducks, muscovy ducks, geese, swan geese | 6000 |
| 4 | 1 | L-BGB (breeding) | Pheasants | 850 |
| 5 | 1 | F-RW (breeding) | Ducks, muscovy ducks, geese, swan geese | 4800 |
| 6 | 1 | F-RW (restocking, fattening) | Mallards | 1600–3200 |
| 7 | 1 | F-RW (fattening) | Ducks, geese | 1500 |
| 8 | 1 | D (ornamental species dealer) | Ornamental ducks and chickens, pigeons, geese, peacocks, turkeys | 1000 |
| 9 | 1 | G (growing) | Ducks, chickens | 500–1000 |
| 10 | 1 | L-BGB (hatching, breeding, restocking) | Pheasants, red-legged partridges, quails, grey partridges | 5000–45,000 |
| 11 | 1 | L-BGB (restocking) | Pheasants | 48,000 |
| 12 | 4 | L-BGB (restocking) | Pheasants | 10,000–100,000 |
| 13 | 4 | G (growing) | Ducks, guinea fowls, geese, chickens, turkeys | >10,000 |
| 14 | 1 | D (poultry dealer) | Ducks, guinea fowls, geese, chickens, quails | 2000–3000 |
| 15 | 2 | F-RW (hatching, breeding, restocking) | Mallards | 1000–8000 |
| 16 | 1 | F-RW (breeding) | Geese | 1600 |
| 17 | 1 | L-BGB (breeding) | Pheasants, red-legged partridges, rock partridges, grey partridges | 1000 |
| Farm ID Number † | No. of PWs Examined/Employed (%) | Yearly Average Occupational Exposure (Min–Max) | AIV Isolation in the Farm | HI Seropositivity in Bird Groups (July 2005–June 2006) ‡, § |
|---|---|---|---|---|
| 1 | 1/1 (100) | 23 | - | H1, H2, H6, H9, H10 |
| 2 | 1/4 (25) | 31 | - | H12 |
| 3 | 3/20 (15) | 11 (8–15) | H4N6 | H4, H11, H13 |
| 4 | 1/2 (50) | 30 | - | H14 |
| 5 | 1/4 (25) | 15 | - | H1, H4, H5, H6, H7, H8, H10, H11, H13, H14 |
| 6 | 0/1 | na | H4N6 | H2, H4, H5 |
| 7 | 0/2 | na | - | none * |
| 8 | 1/2 (50) | 15 | - | H1, H4, H5, H12, H13, H14 |
| 9 | 0/1 | na | - | none ¶ |
| 10 | 4/5 (80) | 20.5 (2–48) | - | none ** |
| 11 | 11/20 (55) | 12.8 (4–31) | - | H4 |
| 12 | 2/8 (25) | 14 (8–20) | - | H12 |
| 13 | 11/11 (100) | 13.2 (2–40) | - | H2, H9, H12 |
| 14 | 1/3 (33.3) | 30 | H2N2 | none ¶ |
| 15 | 2/2 (100) | 1 (1–1) | H10N7 | H1, H2, H4, H6, H9, H10, H12, H13 |
| 16 | 1/3 (33.3) | 8 | - | H1, H2, H3, H4, H5, H6, H7, H11, H12, H13 |
| 17 | 6/7 (85.7) | 15.3 (4–25) | - | H10, H12 |
| Variable | 1st Sampling Period † | 2nd Sampling Period ‡ | 3rd Sampling Period § | |||
|---|---|---|---|---|---|---|
| BEWs, n = 54 | Cs, n = 6 | BEWs, n = 46 | Cs, n = 6 | BEWs, n = 34 | Cs, n = 6 | |
| Age group ^ (%) | ||||||
| 20–29 | 4 (7.4) | 2 (33.3) | 4 (8.7) | 2 (33.3) | 0 | 3 (50) |
| 30–39 | 12 (22.2) | 0 | 11 (23.9) | 0 | 8 (23.5) | 0 |
| 40–49 | 16 (29.6) | 2 (33.3) | 12 (26.1) | 2 (33.3) | 13 (38.2) | 2 (33.3) |
| 50–59 | 14 (25.9) | 2 (33.3) | 14 (30.4) | 2 (33.3) | 9 (26.5) | 1 (16.7) |
| ≥60 | 8 (14.8) | 0 | 5 (10.9) | 0 | 4 (11.8) | 0 |
| Gender (%) | ||||||
| Male | 35 (64.8) | 3 (50) | 27 (58.7) | 3 (50) | 23 (67.6) | 4 (66.7) |
| Female | 19 (35.2) | 3 (50) | 19 (41.3) | 3 (50) | 11 (32.4) | 2 (33.3) |
| Nationality (%) | ||||||
| North Africa | 3 (5.6) | 0 | 3 (6.5) | 0 | 2 (5.9) | 0 |
| West Africa | 4 (7.4) | 0 | 3 (6.5) | 0 | 3 (8.8) | 0 |
| Italy | 47 (87) | 6 (100) | 40 (87) | 6 (100) | 29 (85.3) | 6 (100) |
| YR of OE (%) | ||||||
| ≤2 | 5 (9.3) | na | 3 (6.5) | na | 0 | na |
| 2–10 | 17 (31.5) | na | 15 (32.6) | na | 9 (26.5) | na |
| 10–20 | 20 (37) | na | 17 (37) | na | 18 (52.9) | na |
| 20–30 | 8 (14.8) | na | 7 (15.2) | na | 4 (11.8) | na |
| >30 | 4 (7.4) | na | 4 (8.7) | na | 3 (8.8) | na |
| Titer against | 1st Samplig Period ** | 2nd Sampling Period ** | 3rd Sampling Period ** | |||
|---|---|---|---|---|---|---|
| AIV Strain * | BEWs | Cs | BEWs | Cs | BEWs | Cs |
| H1N1/05 | ||||||
| <1:10 | 34 | 3 | 35 | 4 | 30 | 5 |
| 1:10 | 1 | 0 | 1 | 0 | 0 | 0 |
| 1:20 | 11 | 0 | 1 | 2 | 2 | 1 |
| 1:40 | 5 | 2 | 9 | 0 | 1 | 0 |
| 1:80 | 3 | 1 | 0 | 0 | 1 | 0 |
| GMT | 9.5 | 15.9 | 7.9 | 7.9 | 6.3 | 6.3 |
| H2N3/73 | ||||||
| <1:10 | 38 | 5 | 30 | 3 | 23 | 5 |
| 1:10 | 8 | 1 | 9 | 2 | 10 | 1 |
| 1:20 | 7 | 0 | 4 | 1 | 1 | 0 |
| 1:40 | 1 | 0 | 3 | 0 | 0 | 0 |
| GMT | 6.9 | 5.6 | 7.4 | 7.9 | 6.4 | 5.6 |
| H3N8/04 | ||||||
| <1:10 | 24 | 2 | 21 | 2 | 16 | 2 |
| 1:10 | 9 | 2 | 12 | 2 | 6 | 2 |
| 1:20 | 12 | 2 | 8 | 1 | 6 | 2 |
| 1:40 | 7 | 0 | 3 | 1 | 4 | 0 |
| 1:80 | 2 | 0 | 2 | 0 | 2 | 0 |
| GMT | 11.1 | 10.0 | 9.9 | 11.2 | 10.8 | 10.0 |
| H4N6/05 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H5N9/97 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H5N3/00 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H5N1/05 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H6N5/72 | ||||||
| <1:10 | 53 | 6 | 46 | 6 | 33 | 6 |
| 1:10 | 0 | 0 | 0 | 0 | 1 | 0 |
| 1:20 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1:40 | 1 | 0 | 0 | 0 | 0 | 0 |
| GMT | 5.2 | 5 | 5 | 5 | 5.1 | 5 |
| H7N1/99 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H7N3/02 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H8N4/68 | ||||||
| <1:10 | 53 | 6 | 45 | 6 | 33 | 6 |
| 1:10 | 1 | 0 | 1 | 0 | 0 | 0 |
| 1:20 | 0 | 0 | 0 | 0 | 1 | 0 |
| GMT | 5.1 | 5 | 5.1 | 5 | 5.2 | 5 |
| H9N8/03 | ||||||
| <1:10 | 53 | 6 | 45 | 6 | 33 | 6 |
| 1:10 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1:20 | 1 | 0 | 1 | 0 | 0 | 0 |
| 1:40 | 0 | 0 | 0 | 0 | 1 | 0 |
| GMT | 5.1 | 5 | 5.2 | 5 | 5.3 | 5 |
| H10N7/05 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H11N9/05 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 33 | 6 |
| 1:10 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1:20 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1:40 | 0 | 0 | 0 | 0 | 1 | 0 |
| GMT | 5.0 | 5.0 | 5.0 | 5.0 | 5.3 | 5.0 |
| H12N5/76 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| H13N6/77 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 33 | 6 |
| 1:10 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1:20 | 0 | 0 | 0 | 0 | 1 | 0 |
| GMT | 5 | 5 | 5 | 5 | 5.2 | 5 |
| H14N5/82 | ||||||
| <1:10 | 54 | 6 | 46 | 6 | 34 | 6 |
| GMT | 5 | 5 | 5 | 5 | 5 | 5 |
| Sex | Age Group | BEWs Type | BEY | AIVs Used in HI | HI Titer | AIVs Used in MN * | MN Titer | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Period | Sampling Period | ||||||||||||
| 1st ** | 2nd ** | 3rd ** | 1st ** | 2nd ** | 3rd ** | 4th ^ | 5th ^^ | ||||||
| F | 50–59 | L-BGB | 19 | H8N4/68 | 10 | 10 | 20 | H8N4/06 | 93.0 | na | 204.0 | na | na |
| H9N8/03 | 20 | 20 | 40 | H9N8/03 | <10 | <10 | 27.5 | na | na | ||||
| M | >60 | G | 30 | H6N5/72 | 40 | na | 10 | H6N5/72 | 430 | na | 292 | na | na |
| M | >60 | G | 2 | H3N8/04 | 20 | 20 | na | H3N8/04 | 181.0 | 147.0 | na | na | na |
| M | 40–49 | WP | 15 | H11N6/56 | na | na | 80 | H11N6/56 | na | na | 99.0 | 98.7 | 36.5 |
| H11N9/00 | na | na | na | H11N9/00 | na | na | 307.9 | 289.2 | 117.8 | ||||
| H11N9/05 | na | na | 40 | H11N9/05 | na | na | 47.0 | 50.0 | 14.0 | ||||
| H13N6/77 | na | na | 20 | H13N6/77 | na | na | 33.3 | 31.7 | 15.2 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marco, M.A.; Delogu, M.; Facchini, M.; Di Trani, L.; Boni, A.; Cotti, C.; Graziosi, G.; Venturini, D.; Regazzi, D.; Ravaioli, V.; et al. Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface. Microorganisms 2021, 9, 2153. https://doi.org/10.3390/microorganisms9102153
De Marco MA, Delogu M, Facchini M, Di Trani L, Boni A, Cotti C, Graziosi G, Venturini D, Regazzi D, Ravaioli V, et al. Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface. Microorganisms. 2021; 9(10):2153. https://doi.org/10.3390/microorganisms9102153
Chicago/Turabian StyleDe Marco, Maria Alessandra, Mauro Delogu, Marzia Facchini, Livia Di Trani, Arianna Boni, Claudia Cotti, Giulia Graziosi, Diana Venturini, Denise Regazzi, Valentina Ravaioli, and et al. 2021. "Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface" Microorganisms 9, no. 10: 2153. https://doi.org/10.3390/microorganisms9102153
APA StyleDe Marco, M. A., Delogu, M., Facchini, M., Di Trani, L., Boni, A., Cotti, C., Graziosi, G., Venturini, D., Regazzi, D., Ravaioli, V., Marzadori, F., Frasnelli, M., Castrucci, M. R., & Raffini, E. (2021). Serologic Evidence of Occupational Exposure to Avian Influenza Viruses at the Wildfowl/Poultry/Human Interface. Microorganisms, 9(10), 2153. https://doi.org/10.3390/microorganisms9102153