
Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. House Fly Colony
2.2. Virus and Polymerase Chain Reaction
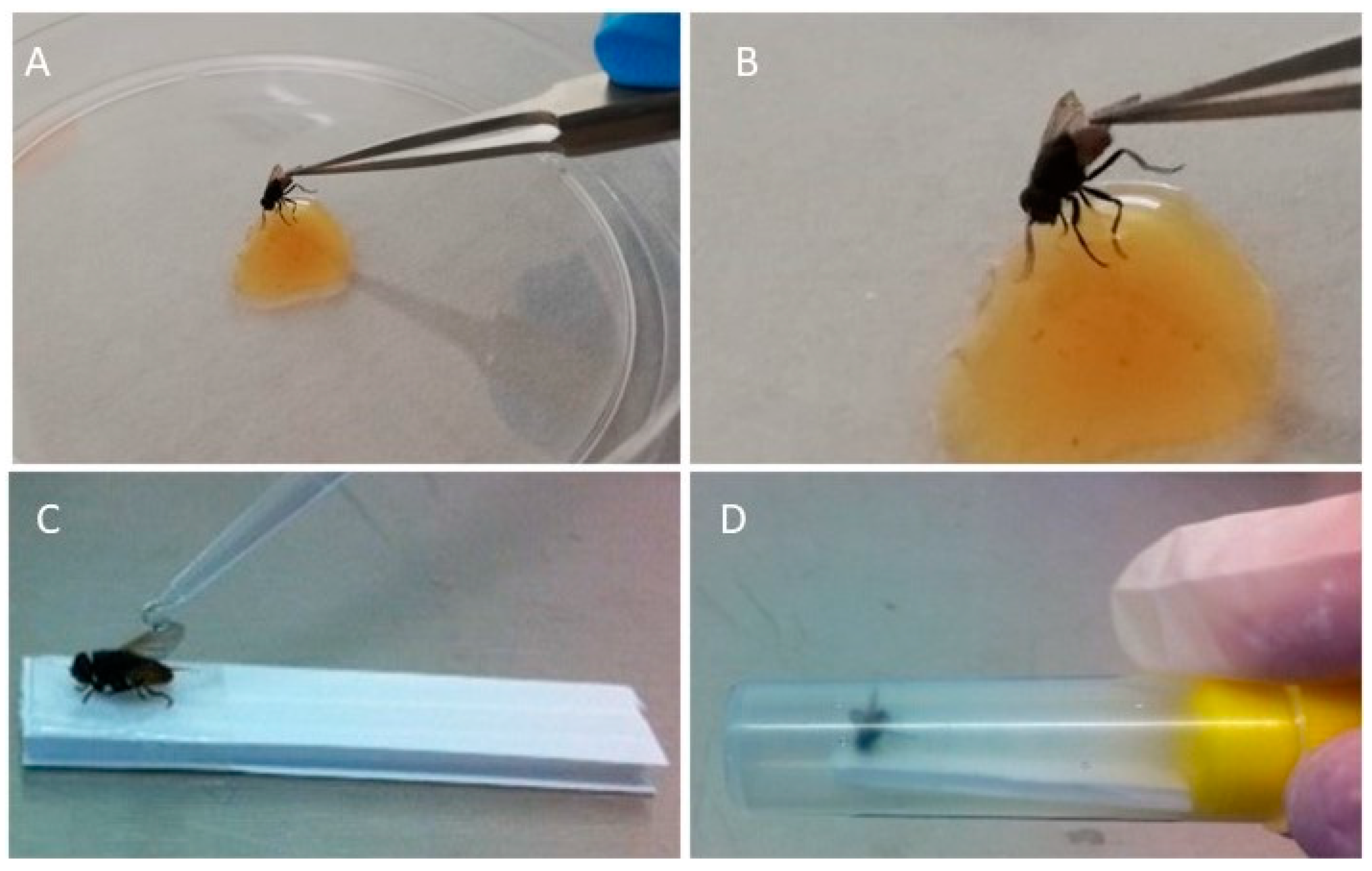
2.3. Infection of Flies with ORF Virus
2.4. Statistical Analysis
3. Results
3.1. Test 1 and Test 2
3.2. Test 3
4. Discussion
Observations and Limitations of the Study
- -
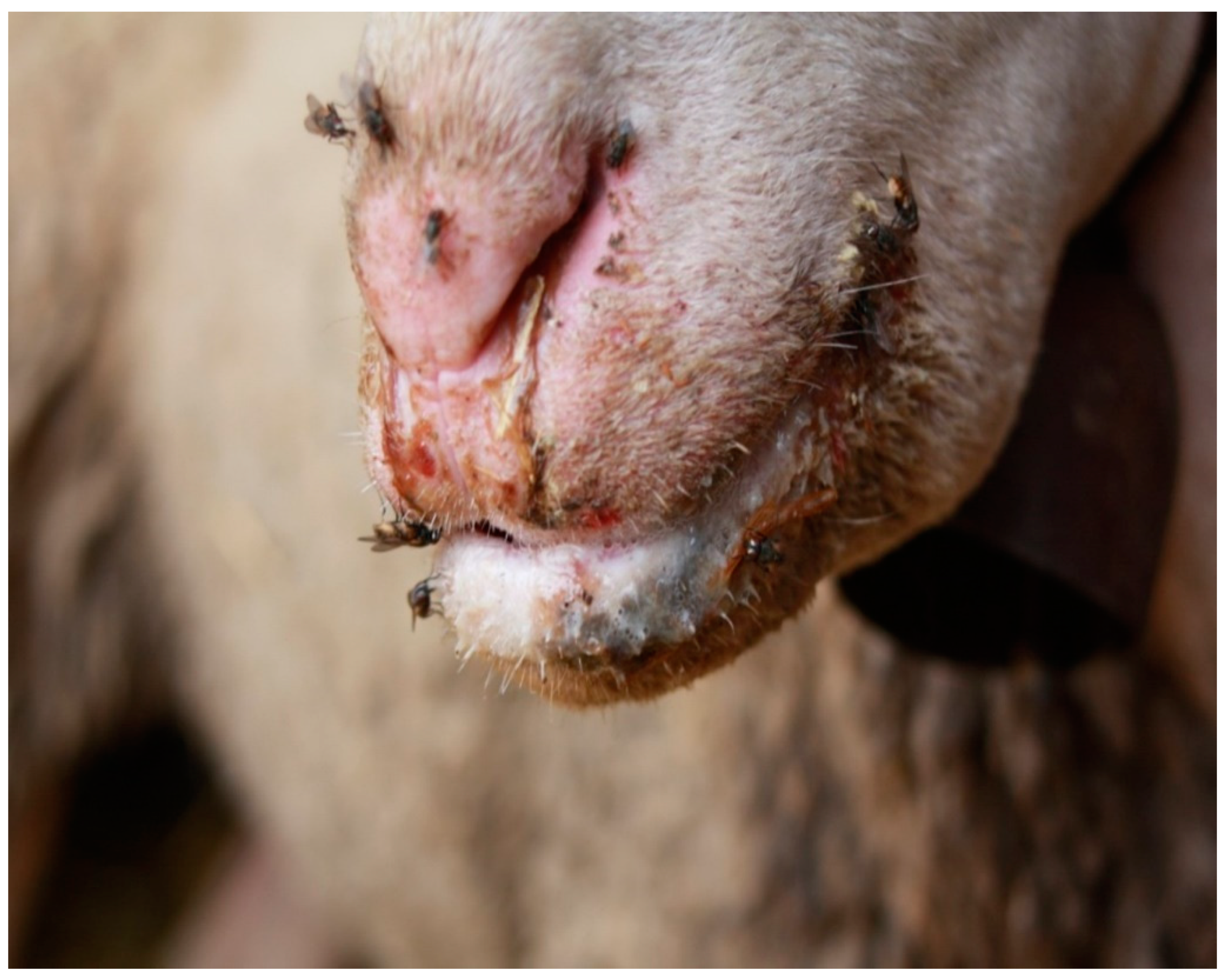
- Particularly, crusty lesions in infected sheep/goats represent the preferential site for nourishment of house flies (Figure 3).
- -
- Fresh ORFV-infected crusts, containing live virus, are the main source of environmental contamination.
- -
- Our study identified the presence of ORF viral genome in flies; no information is available on the presence of live virus in the flies.
- -
- More flies should be involved in future studies to better take into consideration the variability of behavior of flies.
- -
- Long-term studies based on sampling of flies in the field during ORF virus outbreaks would be needed to really understand the role of house flies in the spread of the pathogen.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosamani, M.; Scagliarini, A.; Bhanuprakash, V.; McInnes, C.J.; Singh, R.K. ORF: An update on current research and future perspectives. Expert Rev. Anti. Infect. Ther. 2009, 7, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, Y.; Kong, H.; Shang, Y.; Liu, X. Human infection with ORF virus from goats in China, 2012. Vector Borne Zoonotic. Dis. 2014, 14, 365–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haig, D.M.; Mercer, A. Ovine diseases. ORF. Vet. Res. 1998, 29, 311–326. [Google Scholar] [PubMed]
- Kuhl, J.T.; Huerter, C.J.; Hashish, H. A case of human ORF contracted from a deer. Cutis 2003, 71, 288–290. [Google Scholar] [PubMed]
- Haddock, E.S.; Cheng, C.E.; Bradley, J.S.; Hsu, C.H.; Zhao, H.; Davidson, W.B.; Barrio, V.R. Extensive orf infection in a toddler with associated id reaction. Pediatr. Dermatol. 2017, 34, e337–e340. [Google Scholar] [CrossRef] [PubMed]
- Haig, D.M.; McInnes, C.J.; Hutchison, G.; Seow, H.-F.; Reid, H.W. Cyclosporin A abrogates the acquired immunity to cutaneous reinfection with the parapoxvirus orf virus. Immunology 1996, 89, 524–531. [Google Scholar] [CrossRef]
- Veraldi, S.; Esposito, L.; Pontini, P.; Vaira, F.; Nazzaro, G. Feast of Sacrifice and ORF, Milan, Italy, 2015–2018. Emerg. Infect. Dis. 2019, 25, 1585–1586. [Google Scholar] [CrossRef] [Green Version]
- Kettle, D.S. Medical and Veterinary Entomology: Muscidae (Houseflies, Stableflies); Kettle: Wallingford, UK, 1990. [Google Scholar]
- Greenberg, B. Flies and Disease: Ecology, Classification, and Biotic Associations; Princeton University Press: Princeton, NJ, USA, 1971; Volume I. [Google Scholar]
- Greenberg, B. Flies and Disease: Biology and Disease Transmission; Princeton University Press: Princeton, NJ, USA, 1973; Volume II. [Google Scholar]
- Giangaspero, A. Le Mosche di Interesse Veterinario. I Muscidae. Guida alla Conoscenza e al Riconoscimento; Edagricole: Bologna, Italy, 1997. [Google Scholar]
- Giangaspero, A.; Cafiero, M.A. Contributo alla conoscenza delle specie di mosche presenti negli allevamenti dei bovini dell’Italia meridionale. In Proceedings of the Symposium, Federazione SISVET, Società Italiana Scienze Veterinarie, Altavilla Milicia, Italy, 25–28 September 1991, 45, 1419–1421. [Google Scholar]
- Cafarchia, C.; Lia, R.P.; Romito, D.; Otranto, D. Competence of the housefly, Musca domestica, as a vector of Microsporum canis under experimental conditions. Med. Vet. Entomol. 2009, 23, 21–25. [Google Scholar] [CrossRef]
- Gregorio, S.B.; Nakao, J.C.; Beran, G.W. Human enteroviruses in animals and arthropods in central Philippines, Southeast Asian. J. Trop. Med. Public Health 1972, 3, 45–51. [Google Scholar]
- Graczyk, T.K.; Knight, R.; Gilman, R.H.; Cranfield, M.R. The role of non-biting flies in the epidemiology of human infectious diseases. Microbes Infect. 2001, 3, 231–235. [Google Scholar] [CrossRef]
- Wanaratana, S.; Panyim, S.; Pakpinyo, S. The potential of house flies to act as a vector of avian influenza subtype H5N1 under experimental conditions. Med. Vet. Entomol. 2011, 25, 58–63. [Google Scholar] [CrossRef]
- Sprygin, A.; Pestova, Y.; Wallace, D.; Tuppurainen, E.; Kononov, A. Transmission of lumpy skin disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef]
- Galante, D.; Cafiero, M.A.; Raele, D.A.; Pugliese, N.; Padalino, I.; Cavaliere, N.; Buonavoglia, C. Identification and characterization of ORF viruses isolated from sheep and goats in Southern Italy. Vet. Ital. 2019, 55, 347–353. [Google Scholar]
- Cafiero, M.A.; (Istituto Zooprofilattico Sperimentale della Puglia e della Basilicata, Foggia, Italy). Personal communication, 2014.
- Webbs, G. Sheep and Goat Pox, Transmission of Capripoxviruses by Various Flies Indicated the Need for a Reassessment of the Methods of Controlling this Disease; Institute for Animal Health: Pirbright, UK, 1980. [Google Scholar]
- Kottaridi, C.; Nomikou, K.; Lelli, R.; Markoulatos, P.; Mangana, O. Laboratory diagnosis of contagious ecthyma: Comparison of different PCR protocols with virus isolation in cell culture. J. Virol. Methods. 2006, 134, 119–124. [Google Scholar] [CrossRef]
- Delhon, G.; Tulman, E.R.; Afonso, C.L.; Lu, Z.; de la Concha-Bermejillo, A.; Lehmkuhl, H.D.; Piccone, M.E.; Kutish, G.F.; Rock, D.L. Genomes of the parapoxviruses ORF virus and bovine papular stomatitis virus. J. Virol. 2004, 78, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Parisot, J.; Fernier, L. The Best Methods of Treating Manure Heaps to Prevent the Hatching of Flies. Q. Bull. Health Organisation. Leag. Nations 1934, 3, 1–31. [Google Scholar]
- Yap, K.L.; Kalpana, M.; Lee, H.L. Wings of the common house fly (Musca domestica L.): Importance in mechanical transmission of Vibrio cholerae. Trop. Biomed. 2008, 25, 1–8. [Google Scholar] [PubMed]
- Jacques, B.J.; Bourret, T.J.; Shaffer, J.J. Role of fly clearing behavior on carriage of Escherichia coli and Pseudomonas aeruginosa. J. Med. Entomol. 2017, 54, 1712–1717. [Google Scholar] [PubMed]
- Spyrou, V.; Valiakos, G. Orf virus infection in sheep or goats. Vet. Microbiol. 2015, 181, 178–182. [Google Scholar]
- Tan, S.W.; Yap, K.L.; Lee, H.L. Mechanical transport of rotavirus by the legs and wings of Musca domestica (Diptera: Muscidae). J. Med. Entomol. 1997, 34, 527–531. [Google Scholar] [CrossRef]
- Steinhart, B. Orf in humans: Dramatic but benign. CJEM 2005, 7, 417–419. [Google Scholar]
- Leavell, U.W.; McNamara, M.J.; Muelling, R.; Talbert, W.M.; Rucker, R.C.; Dalton, A.J. ORF. Report of 19 human cases with clinical and pathological observations. JAMA 1968, 204, 657–664. [Google Scholar] [CrossRef]
- Kugler, J. Musicidae of Israel. Isr. J. Entomol. 1969, 4, 322–337. [Google Scholar]
- Crossland-Marr, L.V. Consuming Local, Thinking Global: Building a Halal Industry in a World of Made in Italy; Washington University: St. Louis, MO, USA, 2020. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Human Orf virus infection from household exposures—United States, 2009–2011. MMWR Morb. Mortal. Wkly. Rep. 2012, 61, 245–248. [Google Scholar]
- Andreani, J.; Fongue, J.; Khalil, J.Y.B.; David, L.; Mougari, S.; Le Bideau, M.; Abrahão, J.; Berbis, P.; La Scola, B. Human Infection with Orf Virus and Description of Its Whole Genome, France, 2017. Emerg. Infect. Dis. 2019, 25, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Engel, H.; Marot, L.; Zech, F.; Thys, F. The “Orf” virus: Focus and review of the literature. Rev. Med. Liege. 2010, 65, 691–695. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Flies Infected | Starter (Infected Ovine Crusts) | K−/K+ | 2 h | 4 h | 6 h | After 6 h | Positive Flies/Total |
|---|---|---|---|---|---|---|---|
| ORFV-DNA in Fly Spots (Vomit/Feces) | ORFV-DNA in Fly Spots (Vomit/Feces) | ORFV-DNA in Fly Spots (Vomit/Feces) | ORFV-DNA in Fly Crop | ||||
| Fly 1 | Ct 106,4 | −/+ | - | - | - | - | 60% (9/15) |
| Fly 2 | Ct 106,4 | −/+ | Ct 105 | - | Ct 104 | - | |
| Fly 3 | Ct 106,4 | −/+ | - | - | - | Ct 105 | |
| Fly 4 | Ct 106,4 | −/+ | - | - | - | - | |
| Fly 5 | Ct 106,4 | −/+ | - | - | - | - | |
| Fly 6 | Ct 106,4 | −/+ | Ct 104 | Ct 104 | Ct 104 | Ct 105 | |
| Fly 7 | Ct 106,4 | −/+ | - | Ct 104 | - | - | |
| Fly 8 | Ct 106,4 | −/+ | Ct 103 | - | - | - | |
| Fly 9 | Ct 106,4 | −/+ | - | Ct 104 | Ct 104 | Ct 105 | |
| Fly 10 | Ct 106,4 | −/+ | - | - | Ct 104 | Ct 105 | |
| Fly 11 | Ct 106,4 | −/+ | - | Ct 104 | Ct 104 | Ct 104 | |
| Fly 12 | Ct 106,4 | −/+ | Ct 103 | Ct 103 | Ct 104 | - | |
| Fly 13 | Ct 106,4 | −/+ | - | - | - | - | |
| Fly 14 | Ct 106,4 | −/+ | - | - | - | - | |
| Fly 15 | Ct 106,4 | −/+ | - | - | - | - | |
| Results (%) | 26% (4/15) | 33% (5/15) | 40% (6/15) | 33% (5/15) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raele, D.A.; Stoffolano, J.G., Jr.; Vasco, I.; Pennuzzi, G.; Nardella La Porta, M.C.; Cafiero, M.A. Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions. Microorganisms 2021, 9, 2185. https://doi.org/10.3390/microorganisms9112185
Raele DA, Stoffolano JG Jr., Vasco I, Pennuzzi G, Nardella La Porta MC, Cafiero MA. Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions. Microorganisms. 2021; 9(11):2185. https://doi.org/10.3390/microorganisms9112185
Chicago/Turabian StyleRaele, Donato Antonio, John G. Stoffolano, Jr., Ilaria Vasco, Germana Pennuzzi, Maria Concetta Nardella La Porta, and Maria Assunta Cafiero. 2021. "Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions" Microorganisms 9, no. 11: 2185. https://doi.org/10.3390/microorganisms9112185
APA StyleRaele, D. A., Stoffolano, J. G., Jr., Vasco, I., Pennuzzi, G., Nardella La Porta, M. C., & Cafiero, M. A. (2021). Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions. Microorganisms, 9(11), 2185. https://doi.org/10.3390/microorganisms9112185