
Genomic Analysis of Prophages from Klebsiella pneumoniae Clinical Isolates

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. K. pneumoniae Isolates Genomes
2.2. Prophage Identification
2.3. Prophage Classification
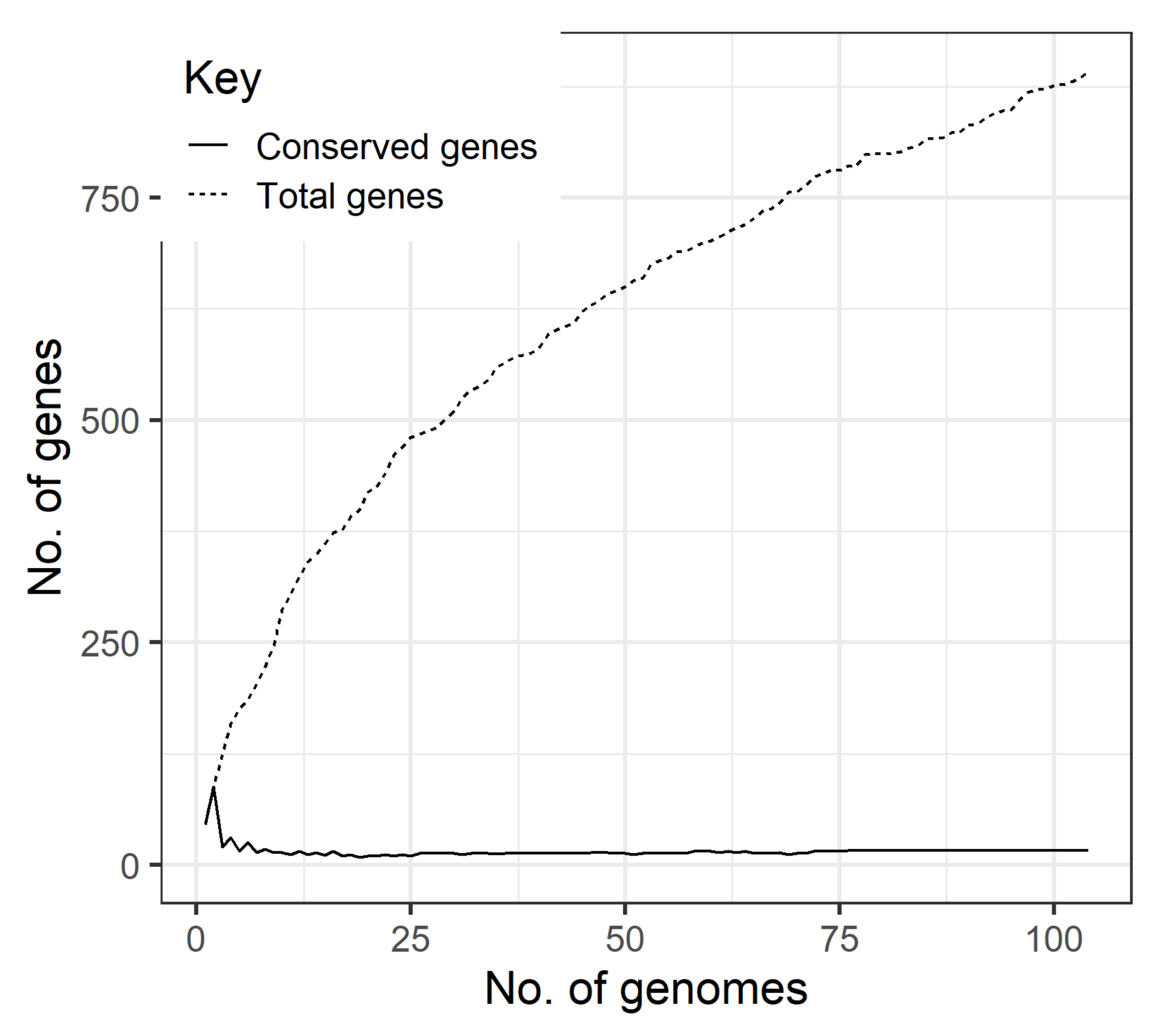
2.4. Prophage Pan-Genome
2.5. Prophage Phylogenetic Analysis
2.6. Prophage-Associated Virulence Factors and Antibiotic Resistance Genes
2.7. Endolysins Identification, Gene Ontology Analysis and Functional Annotation
2.8. Endolysin Phylogenetic Analysis
3. Results
3.1. Identification and Prevalence of Prophages in K. pneumoniae Strains
3.2. Genome Characteristics of K. pneumoniae Prophages
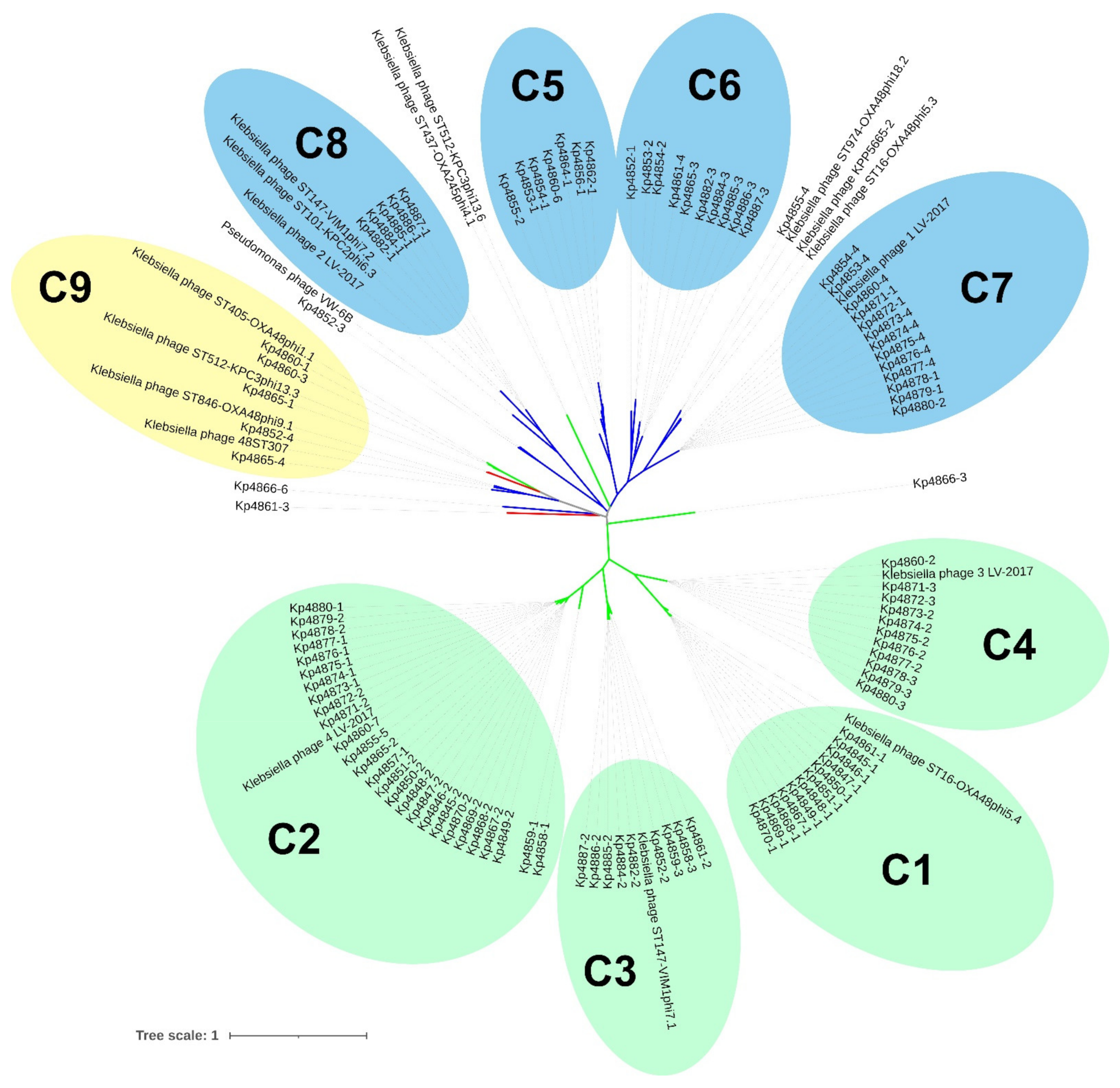
3.3. Classification of K. pneumoniae Prophages
3.4. Genomic and Proteomic Phylogenetic Relationships between K. pneumoniae Prophages
3.5. Presence of Virulence Factors and Antibiotic Resistance Genes within K. pneumoniae Prophages
3.6. Identification of Putative Endolysins within K. pneumoniae Prophages Genomes
3.7. Classification of Endolysins within K. pneumoniae Prophages Genomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae population genomics and antimicrobial-resistant clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [Green Version]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Herridge, W.P.; Shibu, P.; O’Shea, J.; Brook, T.C.; Hoyles, L. Bacteriophages of Klebsiella spp., their diversity and potential therapeutic uses. J. Med. Microbiol. 2020, 69, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Coovadia, Y.M.; Johnson, A.P.; Bhana, R.H.; Hutchinson, G.R.; George, R.C.; Hafferjee, I.E. Multiresistant Klebsiella pneumoniae in a neonatal nursery: The importance of maintenance of infection control policies and procedures in the prevention of outbreaks. J. Hosp. Infect. 1992, 22, 197–205. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti. Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Moolchandani, K. Antimicrobial resistance surveillance among intensive care units of a tertiary care hospital in south India. J. Clin. Diagn. Res. 2017, 11, DC01–DC07. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.; Eberlein, E.; Holley, M.; Schweighart, S.; Rosenthal, E. Spread of Klebsiella pneumoniae producing SHV-5 beta-lactamase among hospitalized patients. Infection 1993, 21, 18–22. [Google Scholar] [CrossRef] [PubMed]
- De Champs, C.; Rouby, D.; Guelon, D.; Sirot, J.; Sirot, D.; Beytout, D.; Gourgand, J.M. A case-control study of an outbreak of infections caused by Klebsiella pneumoniae strains producing CTX-1 (TEM-3) beta-lactamase. J. Hosp. Infect. 1991, 18, 5–13. [Google Scholar] [CrossRef]
- Grundmann, H.; Glasner, C.; Albiger, B.; Aanensen, D.M.; Tomlinson, C.T.; Andrasević, A.T.; Cantón, R.; Carmeli, Y.; Friedrich, A.W.; Giske, C.G.; et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): A prospective, multinational study. Lancet Infect. Dis. 2017, 17, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Bradford, P.A.; Urban, C.; Mariano, N.; Projan, S.J.; Rahal, J.J.; Bush, K. Imipenem resistance in Klebsiella pneumoniae is associated with the combination of ACT-1, a plasmid-mediated AmpC β-lactamase, and the loss of an outer membrane protein. Antimicrob. Agents Chemother. 1997, 41, 563–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twort, F.W. An investigation on the nature of ultra-microscopic viruses. Lancet 1915, 186, 1241–1243. [Google Scholar] [CrossRef] [Green Version]
- D’Hérelle, F. On an invisible microbe antagonistic to dysentery bacilli. Note by M. F. d’Herelle, presented by M. Roux. Comptes Rendus Academie des Sciences 1917; 165:373–5. Bacteriophage 2011, 1, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Debattista, J. Phage therapy: Where east meets west. Expert Rev. Anti. Infect. Ther. 2004, 2, 815–819. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Chen, L.; Porcella, S.F.; Martens, C.A.; Kobayashi, S.D.; Porter, A.R.; Chavda, K.D.; Jacobs, M.R.; Mathema, B.; Olsen, R.J.; et al. Molecular dissection of the evolution of carbapenem-resistant multilocus sequence type 258 Klebsiella pneumoniae. Proc. Natl. Acad. Sci. USA 2014, 111, 4988–4993. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chavda, K.D.; DeLeo, F.R.; Bryant, K.A.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Genome sequence of a Klebsiella pneumoniae sequence type 258 isolate with prophage-encoded K. pneumoniae Carbapenemase. Genome Announc. 2015, 3, e00659-15. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.; Jung, Y.-H.; Lee, S.; Yun, M.; Kim, W.; Kim, D.-W. Comparative genomic analysis of Klebsiella pneumoniae subsp. pneumoniae KP617 and PittNDM01, NUHL24835, and ATCC BAA-2146 reveals unique evolutionary history of this strain. Gut Pathog. 2016, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Fokine, A.; Rossmann, M.G. Molecular architecture of tailed double-stranded DNA phages. Bacteriophage 2014, 4, e28281. [Google Scholar] [CrossRef] [Green Version]
- Fortier, L.-C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Dennehy, J.J.; Abedon, S.T. Bacteriophage ecology. In Bacteriophages; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–42. [Google Scholar] [CrossRef]
- Feiner, R.; Argov, T.; Rabinovich, L.; Sigal, N.; Borovok, I.; Herskovits, A.A. A new perspective on lysogeny: Prophages as active regulatory switches of bacteria. Nat. Rev. Microbiol. 2015, 13, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; Latka, A.; Roszniowski, B.; Valvano, M.A.; Drulis-Kawa, Z. Phage life cycles behind bacterial biodiversity. Curr. Med. Chem. 2017, 24, 3987–4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kim, Y.; Ma, Q.; Hong, S.H.; Pokusaeva, K.; Sturino, J.M.; Wood, T.K. Cryptic prophages help bacteria cope with adverse environments. Nat. Commun. 2010, 1, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R. Bacteriophage lysis: Mechanism and regulation. Microbiol. Rev. 1992, 56, 430–481. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, B.; Olszak, T.; Drulis-Kawa, Z. Applications of bacteriophages versus phage enzymes to combat and cure bacterial infections: An ambitious and also a realistic application? Appl. Microbiol. Biotechnol. 2018, 102, 2563–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borysowski, J.; Weber-Dąbrowska, B.; Górski, A. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as alternatives to antibiotics in clinical care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- De Miguel, T.; Rama, J.L.R.; Sieiro, C.; Sánchez, S.; Villa, T.G. Bacteriophages and lysins as possible alternatives to treat antibiotic-resistant urinary tract infections. Antibiotics 2020, 9, 466. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.C.; Moutinho, C.G.; Pinto, F.C.; Del Fiol, F.S.; Jozala, A.; Chaud, M.V.; Vila, M.M.D.C.; Teixeira, J.A.; Balcão, V.M. Alternatives to overcoming bacterial resistances: State-of-the-art. Microbiol. Res. 2016, 191, 51–80. [Google Scholar] [CrossRef]
- Maciejewska, B.; Roszniowski, B.; Espaillat, A.; Kęsik-Szeloch, A.; Majkowska-Skrobek, G.; Kropinski, A.M.; Briers, Y.; Cava, F.; Lavigne, R.; Drulis-Kawa, Z. Klebsiella phages representing a novel clade of viruses with an unknown DNA modification and biotechnologically interesting enzymes. Appl. Microbiol. Biotechnol. 2017, 101, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Walmagh, M.; Boczkowska, B.; Grymonprez, B.; Briers, Y.; Drulis-Kawa, Z.; Lavigne, R. Characterization of five novel endolysins from Gram-negative infecting bacteriophages. Appl. Microbiol. Biotechnol. 2013, 97, 4369–4375. [Google Scholar] [CrossRef]
- Blasco, L.; Ambroa, A.; Trastoy, R.; Bleriot, I.; Moscoso, M.; Fernández-Garcia, L.; Perez-Nadales, E.; Fernández-Cuenca, F.; Torre-Cisneros, J.; Oteo-Iglesias, J.; et al. In vitro and in vivo efficacy of combinations of colistin and different endolysins against clinical strains of multi-drug resistant pathogens. Sci. Rep. 2020, 10, 7163. [Google Scholar] [CrossRef] [PubMed]
- Perdigão, J.; Modesto, A.; Pereira, A.L.; Neto, O.; Matos, V.; Godinho, A.; Phelan, J.; Charleston, J.; Spadar, A.; de Sessions, P.F.; et al. Whole-genome sequencing resolves a polyclonal outbreak by extended-spectrum beta-lactam and carbapenem-resistant Klebsiella pneumoniae in a Portuguese tertiary-care hospital. Microb. Genom. 2019, 7, mgen000349. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Sun, H.X.; Zhang, C.; Cheng, L.; Peng, Y.; Deng, Z.; Wang, D.; Wang, Y.; Hu, M.; Liu, W.; et al. Prophage hunter: An integrative hunting tool for active prophages. Nucleic Acids Res. 2019, 47, W74–W80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vale, F.; Lehours, P. Relating phage genomes to Helicobacter pylori population structure: General steps using whole-genome sequencing data. Int. J. Mol. Sci. 2018, 19, 1831. [Google Scholar] [CrossRef] [Green Version]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, 29–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modelling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, A.; Tavares, P.; Petit, M.-A.; Guérois, R.; Zinn-Justin, S. Automated classification of tailed bacteriophages according to their neck organization. BMC Genom. 2014, 15, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Yahara, K.; Lehours, P.; Vale, F.F. Analysis of genetic recombination and the pan-genome of a highly recombinogenic bacteriophage species. Microb. Genom. 2019, 5, e000282. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC bioinformatics resource center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, J.G.; Hendrix, R.W.; Casjens, S. Where are the pseudogenes in bacterial genomes? Trends Microbiol. 2001, 9, 535–540. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Ochman, H. The extinction dynamics of bacterial pseudogenes. PLoS Genet. 2010, 6, e1001050. [Google Scholar] [CrossRef]
- Baliga, P.; Shekar, M.; Kallappa, G.S. Genome-wide identification and analysis of chromosomally integrated putative prophages associated with clinical Klebsiella pneumoniae Strains. Curr. Microbiol. 2021, 78, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Bleriot, I.; Trastoy, R.; Blasco, L.; Fernández-Cuenca, F.; Ambroa, A.; Fernández-García, L.; Pacios, O.; Perez-Nadales, E.; Torre-Cisneros, J.; Oteo-Iglesias, J.; et al. Genomic analysis of 40 prophages located in the genomes of 16 carbapenemase-producing clinical strains of Klebsiella pneumoniae. Microb. Genom. 2020, 6, e000369. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.R.; Monteiro, R.; Azeredo, J. Genomic analysis of Acinetobacter baumannii prophages reveals remarkable diversity and suggests profound impact on bacterial virulence and fitness. Sci. Rep. 2018, 8, 15346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, M.E.; Gopalkrishnan, S.; Grace, E.D.; Halliday, J.A.; Gourse, R.L.; Herman, C. DksA and ppGpp regulate the σ S stress response by activating promoters for the small RNA DsrA and the anti-adapter protein IraP. J. Bacteriol. 2018, 200, e00463-17. [Google Scholar] [CrossRef] [Green Version]
- Blankschien, M.D.; Potrykus, K.; Grace, E.; Choudhary, A.; Vinella, D.; Cashel, M.; Herman, C. TraR, a homolog of a RNAP secondary channel interactor, modulates transcription. PLoS Genet. 2009, 5, e1000345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs-Simon, A.; Titball, R.W.; Michell, S.L. Lipoproteins of bacterial pathogens. Infect. Immun. 2011, 79, 548–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Ma, C.; Ye, G.; Shi, Y.; Xu, W.; Zhong, L.; Wang, J.; Yin, Y.; Zhang, X.; Wang, H. DnaJ (hsp40) of Streptococcus pneumoniae is involved in bacterial virulence and elicits a strong natural immune reaction via PI3K/JNK. Mol. Immunol. 2017, 83, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Wang, C.; Zhang, P.; Guo, Z.; Chen, L.; Duan, K. Heat shock protein DnaJ in Pseudomonas aeruginosa affects biofilm formation via pyocyanin production. Microorganisms 2020, 8, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Liu, L.; Vázquez-Torres, A. The DnaK/DnaJ chaperone system enables RNA polymerase-DksA complex formation in Salmonella experiencing oxidative stress. MBio 2021, 12, e03443-20. [Google Scholar] [CrossRef]
- Gancz, A.; Kondratyeva, K.; Cohen-Eli, D.; Navon-Venezia, S. Genomics and virulence of Klebsiella pneumoniae Kpnu95 ST1412 harboring a novel Incf plasmid encoding Blactx-M-15 and Qnrs1 causing community urinary tract infection. Microorganisms 2021, 9, 1022. [Google Scholar] [CrossRef] [PubMed]
- Canova, M.J.; Molle, V. Bacterial serine/threonine protein kinases in host-pathogen interactions. J. Biol. Chem. 2014, 289, 9473–9479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Sang, Y.; Lu, J.; Yao, Y.-F. Protein acetylation and its role in bacterial virulence. Trends Microbiol. 2017, 25, 768–779. [Google Scholar] [CrossRef]
- Shrivastava, S.; Mande, S.S. Identification and functional characterization of gene components of type VI secretion system in bacterial genomes. PLoS ONE 2008, 3, e2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyntum, D.; Venter, S.; Moleleki, L.; Toth, I.; Coutinho, T. Comparative genomics of type VI secretion systems in strains of Pantoea ananatis from different environments. BMC Genom. 2014, 15, 163. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.A.; Kreychman, J.; Gerstein, M. Assessing annotation transfer for genomics: Quantifying the relations between protein sequence, structure and function through traditional and probabilistic scores. J. Mol. Biol. 2000, 297, 233–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, S.; Ahmad, M.; Ahmad, M.; Abdin, M.; Hamid, R.; Khan, M.; Musarrat, J. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Zhou, J.; Xu, Y.; Xiu, Z. Prophages contribute to genome plasticity of Klebsiella pneumoniae and may involve the chromosomal integration of ARGs in CG258. Genomics 2020, 112, 998–1010. [Google Scholar] [CrossRef]
- De Sousa, J.A.M.; Buffet, A.; Haudiquet, M.; Rocha, E.P.C.; Rendueles, O. Modular prophage interactions driven by capsule serotype select for capsule loss under phage predation. ISME J. 2020, 14, 2980–2996. [Google Scholar] [CrossRef] [PubMed]
- Kopotsa, K.; Mbelle, N.M.; Osei Sekyere, J. Epigenomics, genomics, resistome, mobilome, virulome and evolutionary phylogenomics of carbapenem-resistant Klebsiella pneumoniae clinical strains. Microb. Genom. 2020, 6, mgen000474. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, D.; Hou, W.; Jin, Q.; Feng, J.; Zhou, D. Evolutionary diversity of prophage DNA in Klebsiella pneumoniae chromosomes. Front. Microbiol. 2019, 10, 2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobay, L.-M.; Touchon, M.; Rocha, E.P.C. Pervasive domestication of defective prophages by bacteria. Proc. Natl. Acad. Sci. USA 2014, 111, 12127–12132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T. Complete genome sequence of enterohemorrhagic Eschelichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, S.; Rashel, M.; Uchiyama, J.; Sakurai, S.; Ujihara, T.; Kuroda, M.; Imai, S.; Ikeuchi, M.; Tani, T.; Fujieda, M.; et al. Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11, 211–219. [Google Scholar] [CrossRef]
- Ghose, C.; Euler, C.W. Gram-negative bacterial lysins. Antibiotics 2020, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Fischetti, V.A. Bacteriophage lysins as effective antibacterials. Curr. Opin. Microbiol. 2008, 11, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Fenton, M.; McAuliffe, O.; O’Mahony, J.; Coffey, A. Recombinant bacteriophage lysins as antibacterials. Bioeng. Bugs 2010, 1, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Struck, D.K.; Deaton, J.; Wang, I.-N.; Young, R. A signal-arrest-release sequence mediates export and control of the phage P1 endolysin. Proc. Natl. Acad. Sci. USA 2004, 101, 6415–6420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Kuty, G.F.; Arockiasamy, A.; Xu, M.; Young, R.; Sacchettini, J.C. Regulation of a muralytic enzyme by dynamic membrane topology. Nat. Struct. Mol. Biol. 2009, 16, 1192–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prophages | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patients | Strains | Prophages | GC% | Length (kbp) | CDS | Cluster | Family | Related Phages | Accession Number | Query Cover | E Value | Per. Ident |
| Patient 1 | Kp4845 | PKp4845-1 | 50,4 | 33,4 | 44 | C1 | Myoviridae | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% |
| PKp4845-2 | 50,4 | 33,4 | 44 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4846 | PKp4846-1 | 50,4 | 33,4 | 44 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4846-2 | 50,4 | 33,4 | 44 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4847 | PKp4847-1 | 50,4 | 33,4 | 44 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4847-2 | 50,4 | 33,4 | 44 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4848 | PKp4848-1 | 50,4 | 33,4 | 44 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4848-2 | 50,4 | 33,4 | 44 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4850 | PKp4850-1 | 50,4 | 33,4 | 44 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4850-2 | 51,2 | 32,3 | 43 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4851 | PKp4851-1 | 48,3 | 46,7 | 55 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4851-2 | 50,1 | 35,6 | 47 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Patient 2 | Kp4852 | PKp4852-1 | 50,1 | 35,6 | 47 | C6 | Myoviridae | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 35% | 0.0 | 95.09% |
| PKp4852-2 | 50,1 | 35,6 | 47 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 56% | 0.0 | 96.97% | |||
| PKp4852-3 | 50,1 | 35,6 | 47 | N/D | Pseudomonas phage VW-6B | MF975721.1 | 31% | 0.0 | 77.91% | |||
| PKp4852-4 | 50,1 | 35,6 | 47 | C9 | Klebsiella phage ST846-OXA48phi9.1 | MK416021.1 | 77% | 0.0 | 98.39% | |||
| Patient 3 | Kp4853 | PKp4853-1 | 50,1 | 35,6 | 47 | C5 | Myoviridae | Klebsiella phage 2 LV-2017 | KY271396.1 | 54% | 0.0 | 94.42% |
| PKp4853-2 | 50,1 | 35,6 | 47 | C6 | ||||||||
| PKp4853-4 | 50,1 | 35,6 | 47 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Kp4854 | PKp4854-1 | 50,1 | 35,6 | 47 | C5 | Klebsiella phage 2 LV-2017 | KY271396.1 | 54% | 0.0 | 94.42% | ||
| PKp4854-2 | 50,1 | 35,6 | 47 | C6 | ||||||||
| PKp4854-4 | 50,1 | 35,6 | 47 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Patient 4 | Kp4855 | PKp4855-2 | 50,1 | 35,6 | 47 | C5 | Myoviridae | Klebsiella phage ST16-OXA48phi5.3 | MK416014.1 | 28% | 0.0 | 96.13% |
| PKp4855-4 | 51,1 | 31,9 | 43 | N/D | Klebsiella phage ST974-OXA48phi18.2 | MK448237.1 | 68% | 0.0 | 96.79% | |||
| PKp4855-5 | 51,1 | 31,9 | 43 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 82% | 0.0 | 96.43% | |||
| Patient 5 | Kp4856 | PKp4856-1 | 51,1 | 35,1 | 44 | C5 | Myoviridae | Klebsiella phage 2 LV-2017 | KY271396.1 | 46% | 0.0 | 97.25% |
| Patient 6 | Kp4857 | PKp4857-1 | 51,1 | 35,1 | 44 | C2 | Myoviridae | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% |
| Patient 7 | Kp4858 | PKp4858-1 | 51,1 | 35,1 | 44 | C2 | Myoviridae | Klebsiella phage ST512-KPC3phi13.6/Klebsiella phage ST437-OXA245phi4.1 | MK433577.1/MK416011.1 | 86% | 0.0 | 97.25% |
| PKp4858-3 | 51,1 | 35,1 | 44 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 64% | 0.0 | 95.28% | |||
| Patient 8 | Kp4859 | PKp4859-1 | 51,1 | 35,1 | 44 | C2 | Myoviridae | Klebsiella phage ST512-KPC3phi13.6/Klebsiella phage ST437-OXA245phi4.1 | MK433577.1/MK416011.1 | 86% | 0.0 | 97.25% |
| PKp4859-3 | 51 | 34,1 | 40 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 64% | 0.0 | 95.28% | |||
| Patient 9 | Kp4860 | PKp4860-1 | 52,2 | 29,7 | 41 | C9 | Myoviridae | Klebsiella phage ST405-OXA48phi1.1 | MK388859.1 | 88% | 0.0 | 85.05% |
| PKp4860-2 | 52,2 | 29,7 | 41 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4860-3 | 51,3 | 33,5 | 43 | C9 | Klebsiella phage ST405-OXA48phi1.1 | MK388859.1 | 80% | 0.0 | 86.57% | |||
| PKp4860-4 | 50,2 | 36,7 | 43 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| PKp4860-6 | 50,2 | 36,7 | 43 | C5 | Klebsiella phage 2 LV-2017 | KY271396.1 | 36% | 0.0 | 93.97% | |||
| PKp4860-7 | 50,2 | 36,7 | 43 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |||
| Patient 10 | Kp4861 | PKp4861-1 | 50,2 | 36,7 | 43 | C1 | Myoviridae | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 74% | 0.0 | 96.13% |
| PKp4861-2 | 50,2 | 36,7 | 43 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 55% | 0.0 | 97.6% | |||
| PKp4861-3 | 50,2 | 36,7 | 43 | N/D | ||||||||
| PKp4861-4 | 50,2 | 36,7 | 43 | C6 | Klebsiella phage 1 LV-2017 | KY271401.1 | 31% | 0.0 | 96.90% | |||
| Patient 11 | Kp4862 | PKp4862-1 | 50,2 | 36,7 | 43 | C5 | Myoviridae | Klebsiella phage 2 LV-2017 | KY271396.1 | 46% | 0.0 | 97.25% |
| Patient 13 | Kp4864 | PKp4864-1 | 50,2 | 36,7 | 43 | C5 | Myoviridae | Klebsiella phage 2 LV-2017 | KY271396.1 | 35% | 0.0 | 85.93% |
| Patient 14 | Kp4865 | PKp4865-1 | 50,2 | 36,7 | 43 | C9 | Myoviridae | Klebsiella phage ST512-KPC3phi13.3 | MK422448.1 | 73% | 0.0 | 97.23% |
| PKp4865-2 | 50,2 | 36,7 | 43 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 53% | 0.0 | 98.48% | |||
| PKp4865-3 | 49,9 | 37 | 41 | C6 | Klebsiella phage KPP5665-2 | MF695815.1 | 25% | 0.0 | 93.71% | |||
| PKp4865-4 | 54,4 | 35 | 48 | C9 | Klebsiella phage 48ST307 | KY271402.1 | 28% | 0.0 | 94.55% | |||
| Patient 15 | Kp4866 | PKp4866-3 | 54,4 | 35 | 48 | N/D | Myoviridae | |||||
| PKp4866-6 | 54,4 | 35 | 48 | N/D | ||||||||
| Kp4867 | PKp4867-1 | 54,4 | 35 | 48 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4867-2 | 54,4 | 35 | 48 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4868 | PKp4868-1 | 54,4 | 35 | 48 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4868-2 | 54,4 | 35 | 48 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4870 | PKp4870-1 | 54,4 | 35 | 48 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4870-2 | 54,4 | 35 | 48 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Patient 16 | Kp4871 | PKp4871-1 | 50,6 | 39,1 | 63 | C7 | Siphoviridae | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% |
| PKp4871-2 | 50,6 | 39,1 | 63 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |||
| PKp4871-3 | 50,6 | 39,1 | 63 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| Patient 17 | Kp4872 | PKp4872-1 | 50,6 | 39,1 | 63 | C7 | Siphoviridae | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% |
| PKp4872-2 | 50,6 | 39,1 | 63 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |||
| PKp4872-3 | 52 | 40,9 | 57 | C4 | Podoviridae | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | ||
| Kp4873 | PKp4873-1 | 50,2 | 45,9 | 51 | C2 | Siphoviridae | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |
| PKp4873-2 | 53,2 | 34,9 | 32 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4873-4 | 55 | 33,1 | 45 | C7 | Myoviridae | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | ||
| Kp4874 | PKp4874-1 | 50,4 | 39,4 | 49 | C2 | Siphoviridae | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |
| PKp4874-2 | 50,5 | 39,5 | 49 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4874-4 | 50,5 | 39,5 | 49 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Kp4875 | PKp4875-1 | 50,5 | 39,5 | 49 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | ||
| PKp4875-2 | 50,5 | 39,5 | 49 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4875-4 | 50,5 | 39,5 | 49 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Kp4876 | PKp4876-1 | 50,5 | 39,5 | 49 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | ||
| PKp4876-2 | 50,5 | 39,5 | 49 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4876-4 | 50,5 | 39,5 | 49 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Patient 18 | Kp4877 | PKp4877-1 | 50,5 | 39,5 | 49 | C2 | Siphoviridae | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% |
| PKp4877-2 | 50,5 | 39,5 | 49 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| PKp4877-4 | 50,5 | 39,5 | 49 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| Patient 19 | Kp4878 | PKp4878-1 | 50,4 | 39,7 | 49 | C7 | Siphoviridae | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% |
| PKp4878-2 | 50,4 | 40,1 | 50 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |||
| PKp4878-3 | 50,4 | 40,1 | 50 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| Kp4879 | PKp4879-1 | 50,4 | 40,1 | 50 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | ||
| PKp4879-2 | 50,4 | 40,1 | 50 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | |||
| PKp4879-3 | 50,4 | 40,1 | 50 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| Kp4880 | PKp4880-1 | 49,7 | 39,3 | 55 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 100% | 0.0 | 100% | ||
| PKp4880-2 | 50,4 | 41,5 | 61 | C7 | Klebsiella phage 1 LV-2017 | KY271401.1 | 74% | 0.0 | 99.95% | |||
| PKp4880-3 | 50,6 | 38,6 | 56 | C4 | Klebsiella phage 3 LV-2017 | KY271397.1 | 100% | 0.0 | 100% | |||
| Patient 21 | Kp4882 | PKp4882-1 | 51,4 | 39,8 | 67 | C8 | Siphoviridae | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 23% | 0.0 | 97.40% |
| PKp4882-2 | 50,9 | 42,2 | 58 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 70% | 0.0 | 100.00% | |||
| PKp4882-3 | 50,9 | 42,2 | 58 | C6 | Klebsiella phage ST147-VIM1phi7.2 | MK448232.1 | 85% | 0.0 | 99.99% | |||
| Patient 23 | Kp4884 | PKp4884-1 | 49,8 | 45,5 | 60 | C8 | Siphoviridae | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 23% | 0.0 | 97.40% |
| PKp4884-2 | 51,2 | 50,6 | 77 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 70% | 0.0 | 100.00% | |||
| PKp4884-3 | 52,1 | 48,6 | 75 | C6 | Klebsiella phage ST147-VIM1phi7.2 | MK448232.1 | 85% | 0.0 | 99.99% | |||
| Kp4885 | PKp4885-1 | 52,1 | 48,6 | 75 | C8 | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 23% | 0.0 | 97.40% | ||
| PKp4885-2 | 51,5 | 46,8 | 78 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 70% | 0.0 | 100.00% | |||
| PKp4885-3 | 53 | 46,9 | 68 | C6 | Klebsiella phage ST147-VIM1phi7.2 | MK448232.1 | 85% | 0.0 | 99.99% | |||
| Patient 24 | Kp4886 | PKp4886-1 | 53 | 47,1 | 68 | C8 | Siphoviridae | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 23% | 0.0 | 97.40% |
| PKp4886-2 | 52,7 | 48,4 | 73 | C3 | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 70% | 0.0 | 100.00% | |||
| PKp4886-3 | 54,9 | 30,5 | 45 | C6 | Myoviridae | Klebsiella phage ST147-VIM1phi7.2 | MK448232.1 | 85% | 0.0 | 99.99% | ||
| Patient 25 | Kp4887 | PKp4887-1 | 54,9 | 30,2 | 45 | C8 | Myoviridae | Klebsiella phage ST101-KPC2phi6.3 | MK416017.1 | 23% | 0.0 | 97.40% |
| PKp4887-2 | 54 | 35,2 | 43 | C3 | Siphoviridae | Klebsiella phage ST147-VIM1phi7.1 | MK416018.1 | 70% | 0.0 | 100.00% | ||
| PKp4887-3 | 50,9 | 40,2 | 53 | C6 | Podoviridae | Klebsiella phage ST147-VIM1phi7.2 | MK448232.1 | 85% | 0.0 | 99.99% | ||
| Patient 26 | Kp4849 | PKp4849-1 | 50,4 | 33,4 | 44 | C1 | Myoviridae | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% |
| PKp4849-2 | 50,4 | 33,4 | 44 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
| Kp4869 | PKp4869-1 | 54,4 | 35 | 48 | C1 | Klebsiella phage ST16-OXA48phi5.4 | MK416015.1 | 75% | 0.0 | 96.19% | ||
| PKp4869-2 | 54,4 | 35 | 48 | C2 | Klebsiella phage 4 LV-2017 | KY271398.1 | 70% | 0.0 | 97.67% | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, A.T.; Tanoeiro, L.; Duarte, A.; Gonçalves, L.; Vítor, J.M.B.; Vale, F.F. Genomic Analysis of Prophages from Klebsiella pneumoniae Clinical Isolates. Microorganisms 2021, 9, 2252. https://doi.org/10.3390/microorganisms9112252
Marques AT, Tanoeiro L, Duarte A, Gonçalves L, Vítor JMB, Vale FF. Genomic Analysis of Prophages from Klebsiella pneumoniae Clinical Isolates. Microorganisms. 2021; 9(11):2252. https://doi.org/10.3390/microorganisms9112252
Chicago/Turabian StyleMarques, Andreia T., Luís Tanoeiro, Aida Duarte, Luisa Gonçalves, Jorge M. B. Vítor, and Filipa F. Vale. 2021. "Genomic Analysis of Prophages from Klebsiella pneumoniae Clinical Isolates" Microorganisms 9, no. 11: 2252. https://doi.org/10.3390/microorganisms9112252
APA StyleMarques, A. T., Tanoeiro, L., Duarte, A., Gonçalves, L., Vítor, J. M. B., & Vale, F. F. (2021). Genomic Analysis of Prophages from Klebsiella pneumoniae Clinical Isolates. Microorganisms, 9(11), 2252. https://doi.org/10.3390/microorganisms9112252