
Flexibility of Microcystis Overwintering Strategy in Response to Winter Temperatures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacteria Cultures
2.2. Experimental Design
2.3. Flow Cytometric Analysis of Microalgal Cells
2.3.1. Cell Division
2.3.2. Growth Measurement
2.3.3. Cell Viability
2.3.4. Cell Size, Complexity, and Autofluorescence
2.4. Analyzing the Photochemical Efficiency of Microcystis
2.5. Calculations and Statistics
3. Results
3.1. Microcystis Growth under Different Temperature and Light Conditions
3.2. Re-Growth of Microcystis Cells after an Overwintering for a Period
3.3. Cell Division of Microcystis under Different Temperature and Light Conditions
3.4. Ratio of Undamaged Microcystis Cells under Different Temperature and Light Conditions
3.5. Cell Size and Complexity of Microcystis Cultured under Different Temperature and Light Conditions
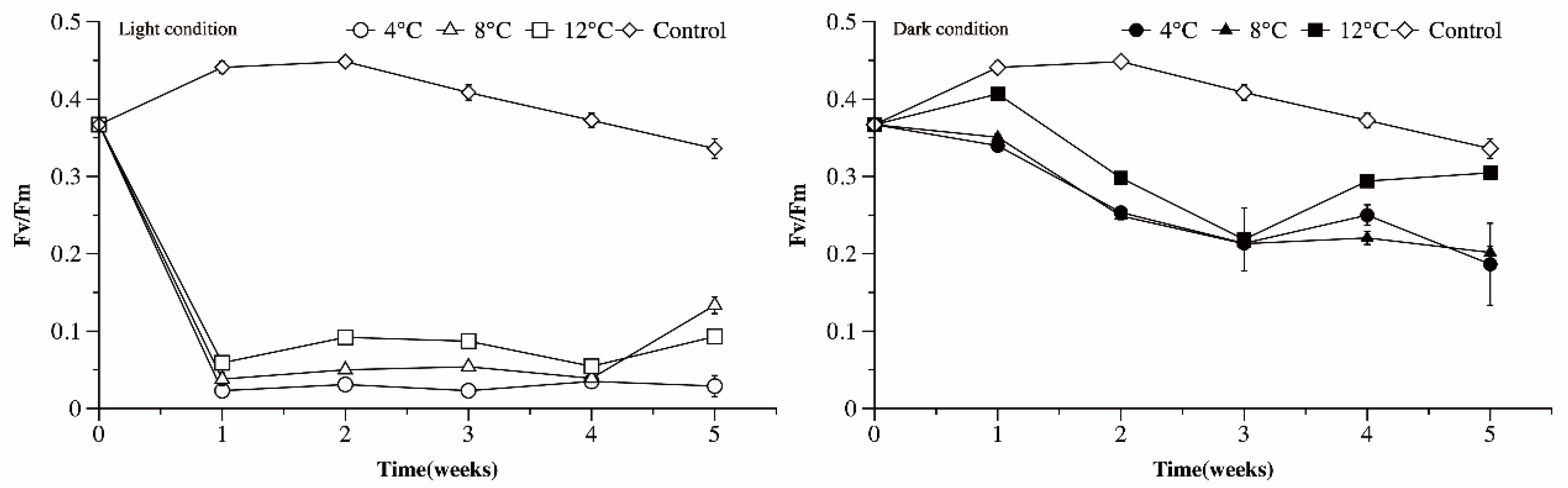
3.6. Photosynthetic Efficiency of Microcystis Cultures under Different Temperature and Light Conditions
4. Discussion
4.1. Response of Microcystis to Different Temperatures and Light Conditions
4.2. Overwintering Strategies of Microcystis Populations in Lakes Taihu, Chaohu, and Dianchi
4.3. Response of Winter Microcystis Blooms to Global Warming
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortiz, D.; Palmer, J.; Wilkinson, G. Detecting changes in statistical indicators of resilience prior to algal blooms in shallow eutrophic lakes. Ecosphere 2020, 11, e03200. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Dittmann, E.; Wiegand, C. Cyanobacterial toxins—Occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.S.; Jaworski, G.H.M.; Cmiech, H.A.; Leedale, G.F. On the annual cycle of the blue-green alga Microcystis Aeruginosa Kütz. Emend. Elenkin. Philos. Trans. R. Soc. B Biol. Sci. 1981, 293, 419–477. [Google Scholar] [CrossRef]
- Ihle, T.; Jähnichen, S.; Benndorf, J. Wax and wane of Microcystis (Cyanophyceae) and microcystins in Lake sediments: A case study in Quitzdorf reservoir (Germany). J. Phycol. 2005, 41, 479–488. [Google Scholar] [CrossRef]
- Kong, F.X.; Gao, G. Hypothesis on cyanobacteria bloom-forming mechanism in large shallow eutrophic lakes. Acta Ecol. Sin. 2005, 25, 589–595. [Google Scholar]
- Tsujimura, S.; Tsukada, H.; Nakahara, H.; Nakajima, T.; Nishino, M. Seasonal variations of Microcystis populations in sediments of Lake Biwa, Japan. Hydrobiologia 2000, 434, 183–192. [Google Scholar] [CrossRef]
- Brunberg, A.K.; Blomqvist, P. Benthic overwintering of Microcystis colonies under different environmental conditions. J. Plankton Res. 2002, 24, 1247–1252. [Google Scholar] [CrossRef]
- Sabart, M.; Misson, B.; Jobard, M.; Bronner, G.; Donnadieu-Bernard, F.; Duffaud, E.; Salençon, M.-J.; Amblard, C.; Latour, D. Genetic diversity along the life cycle of the cyanobacterium Microcystis: Highlight on the complexity of benthic and planktonic interactions. Environ. Microbiol. 2014, 17, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Zhang, Y.; Zhang, Y.; Li, N.; Qin, B.; Zhu, G.; Zhou, Y. Phenology of Phytoplankton Blooms in a Trophic Lake Observed from Long-Term MODIS Data. Environ. Sci. Technol. 2019, 53, 2324–2331. [Google Scholar] [CrossRef]
- Chen, S.L.; Lu, J.W.; Shen, J.Q. Study of the spatio-temporal change of water temperature in Lake Taihu. Jiangshu Water Res. 2009, 3, 38–39. (In Chinese) [Google Scholar]
- Hu, M.Q.; Zhang, Y.C.; Ma, R.H.; Zhang, Y.X. Spatial and Temporal Dynamics of Floating Algal Blooms in Lake Chaohu in 2016 and Their Environmental Drivers. Environ. Sci. 2018, 39, 87–99. (In Chinese) [Google Scholar]
- Sigee, D.C.; Selwyn, A.; Gallois, P.; Dean, A.P. Patterns of cell death in fresh- water colonial cyanobacteria during the late summer bloom. Phycologia 2007, 46, 284–292. [Google Scholar] [CrossRef]
- Ma, J.; Qin, B.; Paerl, H.W.; Brookes, J.D.; Hall, N.S.; Shi, K.; Zhou, Y.; Guo, J.; Li, Z.; Xu, H.; et al. The persistence of cyanobacterial (Microcystis spp.) blooms throughout winter in Lake Taihu, China. Limnol. Oceanogr. 2016, 61, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, L.; Gan, N.; Zheng, L.; Ma, H.; Shan, K.; Liu, J.; Xiao, B.; Song, L. Seasonal dynamics of water bloom-forming Microcystis morphospecies and the associated extracellular microcystin concentrations in large, shallow, eutrophic Dianchi Lake. J. Environ. Sci. 2014, 26, 1921–1929. [Google Scholar] [CrossRef]
- Latour, D.; Salençon, M.J.; Reyss, J.L.; Giraudet, H. Sedimentary imprint of Microcystis aeruginosa (Cyanobacteria) blooms in Grangent reservoir (Loire, France). J. Phycol. 2007, 43, 417–425. [Google Scholar] [CrossRef]
- Wang, C.; Feng, B.; Tian, C.; Tian, Y.; Chen, D.; Wu, X.; Li, G.; Xiao, B. Quantitative study on the survivability of Microcystis colonies in lake sediments. Environ. Boil. Fishes 2017, 30, 495–506. [Google Scholar] [CrossRef]
- Yang, Y.; Yin, C.T.; Li, W.Z.; Xu, X.D. α-Tocopherol is essential for acquired chill-light tolerance in the cyanobacterium Synechocystis sp. Strain PCC 6803. J. Bacteriol. 2008, 190, 1554–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamau, S.W.; Nunez, R.; Grimm, F. Flow cytometry analysis of the effect of allopurinol and the dinitroaniline compound (Chloralin) on the viability and proliferation of Leishmania infantum promastigotes. BMC Pharmacol. 2001, 1, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Rioboo, C.; O’Connor, J.E.; Prado, R.; Herrero, C.; Cid, A. Cell proliferation alterations in Chlorella cells under stress conditions. Aquat. Toxicol. 2009, 94, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Chen, S.; Zeng, J.; Song, W.; Yu, X. Comparing the effects of chlorination on membrane integrity and toxin fate of high- and low-viability cyanobacteria. Water Res. 2020, 177, 115769. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.R. Fluorescent dyes for lymphocyte migration and proliferation studies. Immunol. Cell Biol. 1999, 77, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Lyons, A.B.; Hasbold, J.; Hodgkin, P.D. Flow cytometric analysis of cell division history using dilution of carboxyfluorescein diacetate succinimidyl ester, a stably integrated fluorescent probe. In Methods in Cell Biology: Cytometry, 3rd ed.; Darzynkiewicz, Z., Crissman, H.A., Robinson, J.P., Eds.; Academic Press: London, UK, 2001; Volume 63, pp. 375–398. [Google Scholar]
- Zhou, C.; Place, A.R.; Yan, X.; Xu, J.; Luo, Q.; William, E.; Jiang, Y. Interactions between Karlodinium veneficum and Prorocentrum donghaiense from the East China Sea. Harmful Algae 2015, 49, 50–57. [Google Scholar] [CrossRef]
- Cunningham, A. Analysis of microalgae and cyanobacteria by flow cytometry. In Flow Cytometry in Microbiology; Lloyd, D., Ed.; Springer: London, UK, 1993; pp. 131–142. [Google Scholar]
- Trask, B.J.; Engh, G.J.V.D.; Elgershuizen, J.H.B.W. Analysis of phytoplankton by flow cytometry. Cytometry 1982, 2, 258–264. [Google Scholar] [CrossRef]
- Olson, R.J.; Frankel, S.L.; Chisholm, S.W.; Shapiro, H.M. An inexpensive flow cytometer for the analysis of fluorescence signals in phytoplankton: Chlorophyll and DNA distributions. J. Exp. Mar. Biol. Ecol. 1983, 68, 129–144. [Google Scholar] [CrossRef]
- Brussaard, C.; Marie, D.; Thyrhaug, R.; Bratbak, G. Flow cytometric analysis of phytoplankton viability following viral infection. Aquat. Microb. Ecol. 2001, 26, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.L.; Poot, M.; Yue, S.T.; Millard, P.J. Bacterial viability and antibiotic susceptibility testing with SYTOX green nucleic acid stain. Appl. Environ. Microbiol. 1997, 63, 2421–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermeño, P.; Marañón, E.; Rodríguez, J.; Fernandez, E. Large-sized phytoplankton sustain higher carbon-specific photosynthesis than smaller cells in a coastal eutrophic ecosystem. Mar. Ecol. Prog. Ser. 2005, 297, 51–60. [Google Scholar] [CrossRef]
- Toepel, J.; Wilhelm, C.; Meister, A.; Beckert, A.; Martinez-Ballesta, M.D. Cytometry of freshwater phytoplankton. Methods Cell Biol. 2004, 75, 375–407. [Google Scholar] [PubMed]
- Yang, Y.-P.; Juang, Y.-S.; Hsu, B.-D. A quick method for assessing chloroplastic starch granules by flow cytometry. J. Plant Physiol. 2002, 159, 103–106. [Google Scholar] [CrossRef]
- Ou, H.; Gao, N.; Deng, Y.; Qiao, J.; Wang, H. Immediate and long-term impacts of UV-C irradiation on photosynthetic capacity, survival and microcystin-LR release risk of Microcystis aeruginosa. Water Res. 2012, 46, 1241–1250. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Gan, N.; Liu, J.; Zheng, L.; Li, L.; Song, L. Survival, recovery and microcystin release of Microcystis aeruginosa in cold or dark condition. Chin. J. Oceanol. Limnol. 2016, 35, 313–323. [Google Scholar] [CrossRef]
- Shan, K.; Song, L.; Chen, W.; Li, L.; Liu, L.; Wu, Y.; Jia, Y.; Zhou, Q.; Peng, L. Analysis of environmental drivers influencing interspecific variations and associations among bloom-forming cyanobacteria in large, shallow eutrophic lakes. Harmful Algae 2019, 84, 84–94. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, M.; Yu, Y.; Shi, X. Temperature triggers the annual cycle of Microcystis, comparable results from the laboratory and a large shallow lake. Chemosphere 2020, 260, 127543. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Yang, Z.; Wei, L.; Yang, W.; Chen, C.; Kong, F. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: Patterns and driving factors. Phycol. Res. 2015, 64, 44–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Zhang, M.; Duan, H.; Loiselle, S.; Xu, J. Fourteen-Year Record (2000–2013) of the Spatial and Temporal Dynamics of Floating Algae Blooms in Lake Chaohu, Observed from Time Series of MODIS Images. Remote Sens. 2016, 7, 10523–10542. [Google Scholar] [CrossRef] [Green Version]
- Sellner, K.G. Physiology, ecology, and toxic properties of marine cyanobacteria blooms. Limnol. Oceanogr. 1997, 42, 1089–1104. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A drinking water crisis in Lake Taihu, China: Linkage to climatic variability and lake management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, P.; Cai, Q.; He, F.; Huang, Y.; Tian, C.; Wu, X.; Wang, C.; Xiao, B. Flexibility of Microcystis Overwintering Strategy in Response to Winter Temperatures. Microorganisms 2021, 9, 2278. https://doi.org/10.3390/microorganisms9112278
Cai P, Cai Q, He F, Huang Y, Tian C, Wu X, Wang C, Xiao B. Flexibility of Microcystis Overwintering Strategy in Response to Winter Temperatures. Microorganisms. 2021; 9(11):2278. https://doi.org/10.3390/microorganisms9112278
Chicago/Turabian StyleCai, Pei, Qijia Cai, Feng He, Yuhong Huang, Cuicui Tian, Xingqiang Wu, Chunbo Wang, and Bangding Xiao. 2021. "Flexibility of Microcystis Overwintering Strategy in Response to Winter Temperatures" Microorganisms 9, no. 11: 2278. https://doi.org/10.3390/microorganisms9112278
APA StyleCai, P., Cai, Q., He, F., Huang, Y., Tian, C., Wu, X., Wang, C., & Xiao, B. (2021). Flexibility of Microcystis Overwintering Strategy in Response to Winter Temperatures. Microorganisms, 9(11), 2278. https://doi.org/10.3390/microorganisms9112278