
Interactions of Bacillus subtilis Basement Spore Coat Layer Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of Bacillus Subtilis yncD Strain
2.3. Analysis of Protein–Protein Interactions Using a Bacterial Two Hybrid System (BACTH)
2.4. Protein Expression and Purification and Pull-Down Assay
2.5. Spore Preparation
2.6. Germination Assay
2.7. Thermal Resistance Assay
2.8. Fluorescence Microscopy
3. Results
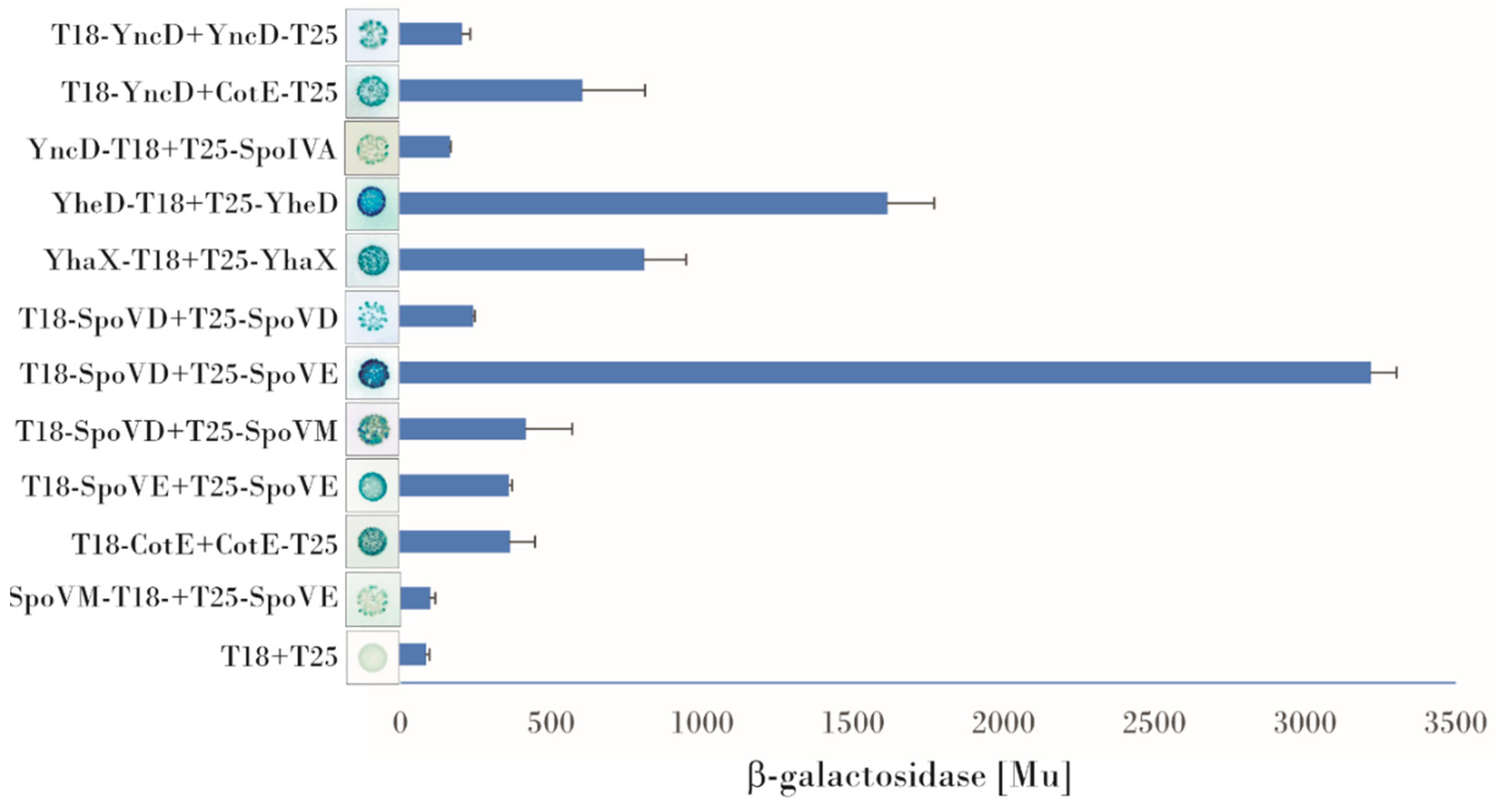
3.1. Examination of Protein-Protein Interactions Using a Bacterial Two Hybrid System
- YncD contacts with morphogenetic coat proteins: YncD, an alanine racemase, is deposited onto a developing forespore under the control of CotE, as reported by McKenney [2]. Using BACTH, we observed that YncD physically contacts CotE. Screening all possible combinations of plasmids with the cotE and yncD genes fused to either the T18 or T25 domains of adenylate cyclase in plate assays showed that the highest expression of the β-galactosidase reporter gene arose from the T18-YncD/CotE-T25 combination (600 Mu). Some signals were also obtained for the combinations T18-YncD/T25-CotE, T25-YncD/CotE-T18, and T25-YncD/T18-CotE. As described previously [31], we compared these signals with the T18-CotE/CotE-T25 self-interaction used as the positive control (360 Mu in these experiments) [18,32]. The strength of the interaction of YncD and CotE observed by BACTH was almost twice that of CotE self-interaction and the observed blue color of the bacterial colonies harboring the YncD/CotE pair after 48 h of incubation was also more intense. A direct interaction between SpoVID or SpoVM and YncD was not detected, but a weak blue color could be observed in those bacterial colonies harboring combinations of plasmids containing the spoIVA and yncD genes. The highest β-galactosidase signal detected on X-gal plates for this pair was obtained from the YncD-T18/T25-SpoIVA and T18-SpoIVA/YncD-T25 combinations (160 Mu), which was approximately twice the negative control. A YncD/YncD self-interaction was also observed (220 Mu), indicating that the protein can form oligomers (Figure 1).
- Protein contacts of YheD and YhaX: Although the deposition of these two envelope proteins on the forespore surface has been shown to be dependent on SpoIVA [20], our bacterial two hybrid assays did not detect any direct binding to SpoIVA or to any other morphogenetic protein, only an YheD self-interaction could be observed. The positive signal was one of the strongest in this screening (1724 Mu) and arose from two plasmid combinations (T25–YheD/YheD–T18 and T25–YheD/T18–YheD). YhaX also formed homooligomers: β-galactosidase reporter gene activity was measured after complementation of the hybrid proteins T25–YhaX/YhaX–T18 and T25-YhaX/T18–YhaX (704 Mu) (Figure 1).
- Bacterial two hybrid assay of interactions of SpoVB, SpoVD and SpoVE with coat morphogenetic proteins: BACTH was previously used to successfully investigate interactions between protein involved in cell wall synthesis [33], many of which are integral membrane proteins or are associated with a membrane. Here we investigated the possible contacts between proteins essential for cortex peptidoglycan synthesis. An analysis of the topology of these proteins predicted that SpoVE and SpoVB have 10–15 transmembrane segments while SpoVD has one transmembrane segment at its N-terminus. Despite some difficulties with cloning, especially of SpoVE and SpoVD which may be toxic to E. coli cells, we managed to prepare the correct plasmid constructs. First, we tested their self-association. No homodimerization of SpoVB was observed, but positive signals were obtained for SpoVE and SpoVD; in both cases, β-galactosidase expression occurred in only one combination of plasmids. For SpoVE, we saw that bacterial colonies turned blue when the SpoVE–T18/T25–SpoVE hybrid proteins were co-expressed and we measured a β-galactosidase activity of up to 360 Mu (Figure 1). For SpoVD, it was important that, after fusion with the adenylate cyclase fragment, the protein be properly incorporated into the membrane such that the T18/T25 domain would be in the cytoplasm (on its N-terminus). Under these restrictions, we detected a positive signal from the combination T18–SpoVD/T25–SpoVD whose β-galactosidase activity reached 240 Mu (Figure 1). We next examined the interactions of these proteins with the coat morphogenetic proteins. Our experiments showed that SpoVD interacts with SpoVM when the hybrid protein T25–SpoVD was combined with T18–SpoVM; very faint blue bacterial colonies were also observed when T25–SpoVD was co-expressed with SpoVM–T18. Quantitative evaluation of the contact strength showed that the interaction of SpoVD with SpoVM was at roughly the same level as the CotE self-interaction (420 Mu vs. 360 Mu), which makes it an intermediate-strength interaction. Screening the interactions of SpoVE with the coat morphogenetic proteins indicated that there may also be an interaction of SpoVE and SpoVM, although the positive signals were very weak and could be identified only after prolonged incubation of the X-gal plate at 4 °C. The signals could be detected only for the T25–SpoVE and SpoVM–T18 and T18–SpoVM combinations (the β-galactosidase activity was approximately twice the negative control). The BACTH system did not detect any contact between these proteins and SpoIVA or between SpoVB and SpoVM or SpoIVA (Figure 1).
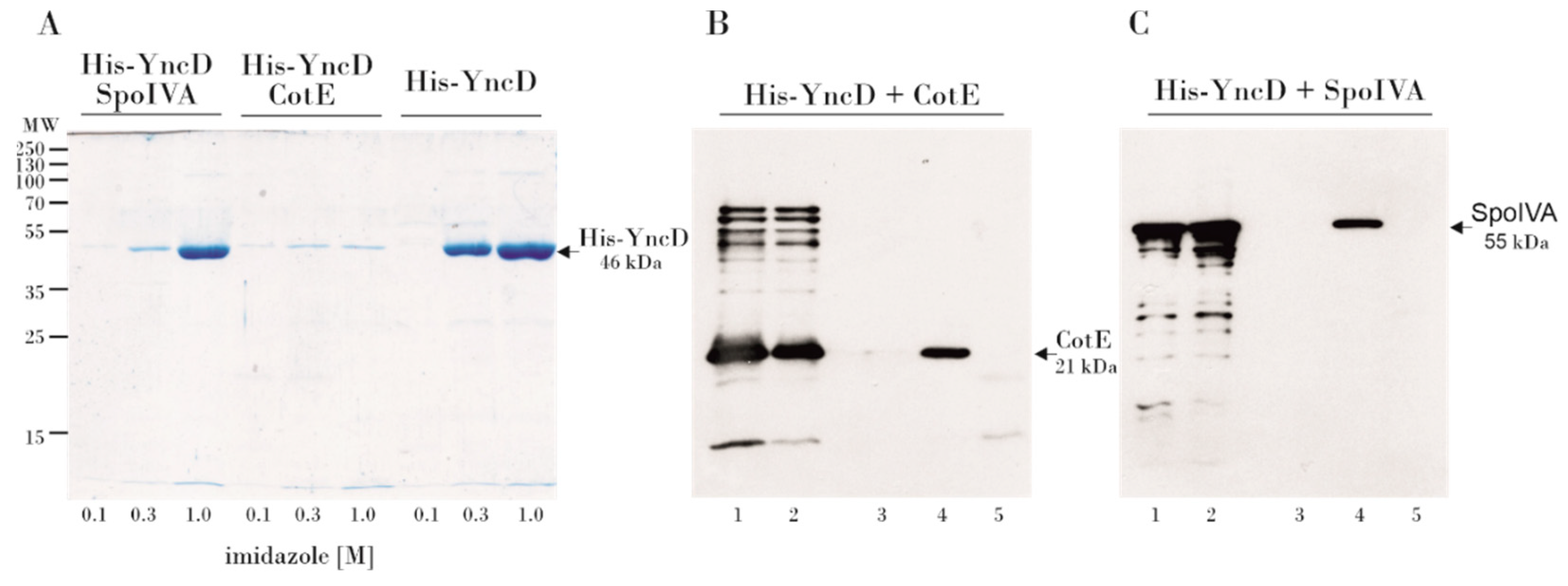
3.2. Examination of the YncD–CotE and YncD–SpoIVA Interactions Using a Pull-Down Assay
3.3. Characterization of the ΔyncD Mutant Spores
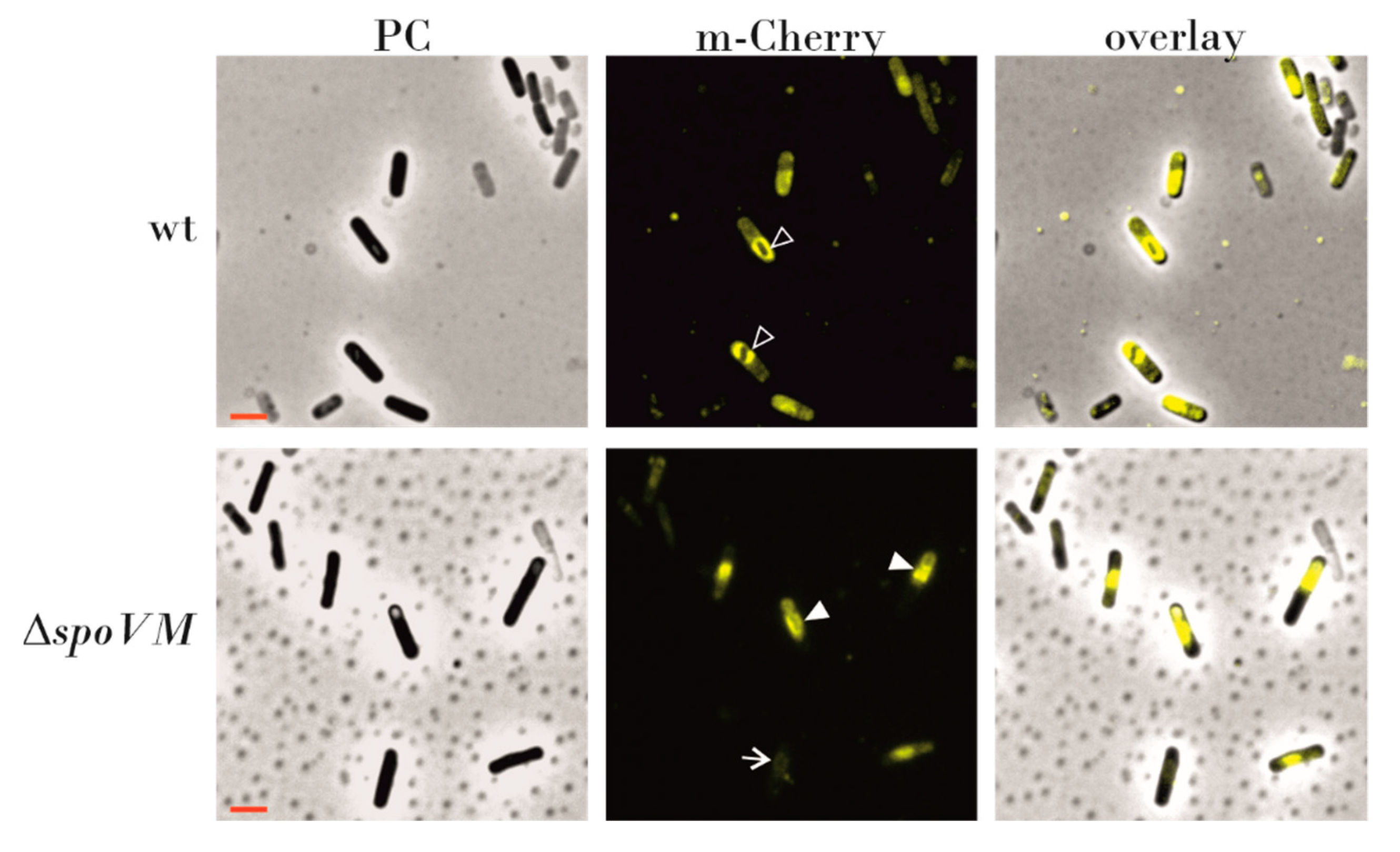
3.4. Localization of SpoVD-mCherry is Dependent on SpoVM
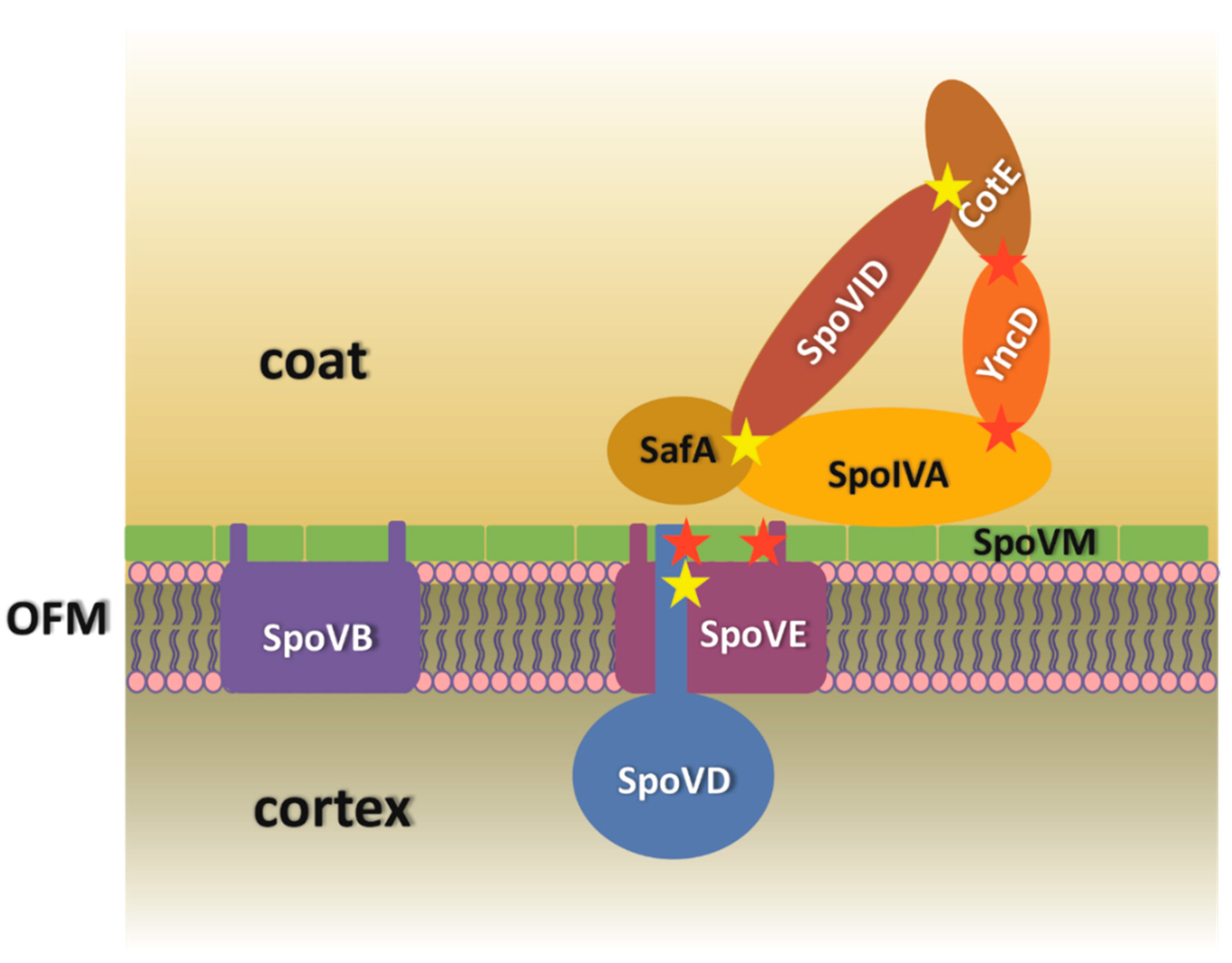
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Driks, A. Bacillus subtilis Spore Coat. Microbiol. Mol. Biol. Rev. 1999, 63, 1–20. [Google Scholar] [CrossRef] [Green Version]
- McKenney, P.T.; Driks, A.; Eskandarian, H.A.; Grabowski, P.; Guberman, J.; Wang, K.H.; Gitai, Z.; Eichenberger, P. A Distance-Weighted Interaction Map Reveals a Previously Uncharacterized Layer of the Bacillus subtilis Spore Coat. Curr. Biol. 2010, 20, 934–938. [Google Scholar] [CrossRef] [Green Version]
- Driks, A.; Eichenberger, P. The Spore Coat. Microbiol Spectr. 2016. [Google Scholar] [CrossRef]
- Levin, P.A.; Fan, N.; Ricca, E.; Driks, A.; Losick, R.; Cutting, S. An unusually small gene required for sporulation by Bacillus subtilis. Mol. Microbiol. 1993, 9, 761–771. [Google Scholar] [CrossRef]
- Roels, S.; Driks, A.; Losick, R. Characterization of spoIVA, a Sporulation Gene Involved in Coat Morphogenesis in Bacillus subtilis. J. Bacteriol. 1992, 174, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Ramamurthi, K.S.; Lecuyer, S.; Stone, H.A.; Losick, R. Geometric Cue for Protein Localization in a Bacterium. Science 2009, 323, 1354–1357. [Google Scholar] [CrossRef] [Green Version]
- Castaing, J.P.; Nagy, A.; Anantharaman, V.; Aravind, L.; Ramamurthi, K.S. ATP hydrolysis by a domain related to translation factor GTPases drives polymerization of a static bacterial morphogenetic protein. Proc. Natl. Acad. Sci. USA 2013, 110, E151–E160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramamurthi, K.S.; Losick, R. ATP-Driven Self-Assembly of a Morphogenetic Protein in Bacillus subtilis. Mol. Cell 2008, 31, 406–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, E.A.; Updegrove, T.B.; Chen, J.; Shroff, H.; Ramamurthi, K.S. A 2-dimensional ratchet model describes assembly initiation of a specialized bacterial cell surface. Proc. Natl. Acad. Sci. USA 2019, 116, 21789–21799. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.H.; Isidro, A.L.; Domingues, L.; Eskandarian, H.A.; McKenney, P.T.; Drew, K.; Grabowski, P.; Chua, M.H.; Barry, S.N.; Guan, M.; et al. The coat morphogenetic protein SpoVID is necessary for spore encasement in Bacillus subtilis. Mol. Microbiol. 2009, 74, 634–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.C.; Nunes, F.; Cruz, F.; Fernandes, C.; Isidro, A.L.; Lousa, D.; Soares, C.M.; Moran, C.P.; Henriques, A.O.; Serrano, M. A LysM Domain intervenes in sequential protein-protein and protein-peptidoglycan interactions important for spore coat assembly in bacillus subtilis. J. Bacteriol. 2019, 201, e00642-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.B.; Donovan, W.P.; Fitz-James, P.C.; Losick, R. Gene encoding a morphogenic protein required in the assembly of the outer coat of the Bacillus subtilis endospore. Genes Dev. 1988, 2, 1047–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamatsu, H.; Chikahiro, Y.; Kodama, T.; Koide, H.; Kozuka, S.; Tochikubo, K.; Watabe, K. A Spore Coat Protein, CotS, of Bacillus subtilis Is Synthesized under the Regulation of K and GerE during Development and Is Located in the Inner Coat Layer of Spores. J. Bacteriol. 1998, 180, 2968–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, T.; Little, S.; Stöver, A.G.; Driks, A. Functional Regions of the Bacillus subtilis Spore Coat Morphogenetic Protein CotE. J. Bacteriol. 1999, 181, 7043–7051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, T.; Isidro, A.L.; Moran, C.P.; Henriques, A.O. Interaction between coat morphogenetic proteins SafA and SpoVID. J. Bacteriol. 2006, 188, 7731–7734. [Google Scholar] [CrossRef] [Green Version]
- Müllerová, D.; Krajčíková, D.; Barák, I. Interactions between Bacillus subtilis early spore coat morphogenetic proteins. FEMS Microbiol. Lett. 2009, 299, 74–85. [Google Scholar] [CrossRef]
- De Francesco, M.D.; Jacobs, J.Z.; Nunes, F.; Serrano, M.; Mckenney, P.T.; Chua, M.H.; Henriques, A.O.; Eichenberger, P. Physical interaction between coat morphogenetic proteins spovid and cote is necessary for spore encasement in Bacillus subtilis. J. Bacteriol. 2012, 194, 4941–4950. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Krajcikova, D.; Xing, C.; Lu, B.; Hao, J.; Ke, X.; Wang, H.; Barak, I.; Tang, J. Study of the interactions between the key spore coat morphogenetic proteins CotE and SpoVID. J. Struct. Biol. 2013, 181, 128–135. [Google Scholar] [CrossRef]
- Liu, H.; Qiao, H.; Krajcikova, D.; Zhang, Z.; Wang, H.; Barak, I.; Tang, J. Physical interaction and assembly of Bacillus subtilis spore coat proteins CotE and CotZ studied by atomic force microscopy. J. Struct. Biol. 2016, 195, 245–251. [Google Scholar] [CrossRef] [Green Version]
- McKenney, P.T.; Eichenberger, P. Dynamics of spore coat morphogenesis in Bacillus subtilis. Mol. Microbiol. 2012, 83, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Pierce, K.J.; Salifu, S.P.; Tangney, M. Gene cloning and characterization of a second alanine racemase from Bacillus subtilis encoded by yncD. FEMS Microbiol. Lett. 2008, 283, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steichen, C.; Chen, P.; Kearney, J.F.; Turnbough, C.L. Identification of the immunodominant protein and other proteins of the Bacillus anthracis exosporium. J. Bacteriol. 2003, 185, 1903–1910. [Google Scholar] [CrossRef] [Green Version]
- Chesnokova, O.N.; McPherson, S.A.; Steichen, C.T.; Turnbough, C.L. The spore-specific alanine racemase of Bacillus anthracis and its role in suppressing germination during spore development. J. Bacteriol. 2009, 191, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebmeier, S.E.; Tan, I.S.; Clapham, K.R.; Ramamurthi, K.S. Small proteins link coat and cortex assembly during sporulation in Bacillus subtilis. Mol. Microbiol. 2012, 84, 682–696. [Google Scholar] [CrossRef] [Green Version]
- Tan, I.S.; Weiss, C.A.; Popham, D.L.; Ramamurthi, K.S. A Quality-Control Mechanism Removes Unfit Cells from a Population of Sporulating Bacteria. Dev. Cell 2015, 34, 682–693. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, P.; Weaver, A.; Reichert, E.D.; Linnstaedt, S.D.; Popham, D.L. Spore cortex formation in Bacillus subtilis is regulated by accumulation of peptidoglycan precursors under the control of sigma K. Mol. Microbiol. 2007, 65, 1582–1594. [Google Scholar] [CrossRef] [PubMed]
- Fay, A.; Meyer, P.; Dworkin, J. Interactions between late-acting proteins required for peptidoglycan synthesis during sporulation. J. Mol. Biol. 2010, 399, 547–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popham, D.L.; Stragier, P. Cloning, Characterization, and Expression of the spoVB Gene of Bacillus subtilis. J. Bacteriol. 1991, 173, 7942–7949. [Google Scholar] [CrossRef] [Green Version]
- Meeske, A.J.; Riley, E.P.; Robins, W.P.; Uehara, T.; Mekalanos, J.J.; Kahne, D.; Walker, S.; Kruse, A.C.; Bernhardt, T.G.; Rudner, D.Z. SEDS proteins are a widespread family of bacterial cell wall polymerases. Nature 2016, 537, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Meeske, A.J.; Sham, L.T.; Kimsey, H.; Koo, B.M.; Gross, C.A.; Bernhardt, T.G.; Rudner, D.Z. MurJ and a novel lipid II flippase are required for cell wall biogenesis in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2015, 112, 6437–6442. [Google Scholar] [CrossRef] [Green Version]
- Krajčíková, D.; Forgáč, V.; Szabo, A.; Barák, I. Exploring the interaction network of the Bacillus subtilis outer coat and crust proteins. Microbiol. Res. 2017, 204, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Krajčíková, D.; Lukáčová, M.; Müllerová, D.; Cutting, S.M.; Barák, I. Searching for protein-protein interactions within the Bacillus subtilis spore coat. J. Bacteriol. 2009, 191, 3212–3219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankute, M.; Byng, C.V.; Alderwick, L.J.; Besra, G.S. Elucidation of a protein-protein interaction network involved in Corynebacterium glutamicum cell wall biosynthesis as determined by bacterial two-hybrid analysis. Glycoconj. J. 2014, 31, 475–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukowska-Faniband, E.; Hederstedt, L. Cortex synthesis during Bacillus subtilis sporulation depends on the transpeptidase activity of SpoVD. FEMS Microbiol. Lett. 2013, 346, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Prágai, Z.; Harwood, C.R. Regulatory interactions between the Pho and σB-dependent general stress regulons of Bacillus subtilis. Microbiology 2002, 148, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Van Ooij, C.; Eichenberger, P.; Losick, R. Dynamic patterns of subcellular protein localization during spore coat morphogenesis in Bacillus subtilis. J. Bacteriol. 2004, 186, 4441–4448. [Google Scholar] [CrossRef] [Green Version]
- Butzin, X.Y.; Troiano, A.J.; Coleman, W.H.; Griffiths, K.K.; Doona, C.J.; Feeherry, F.E.; Wang, G.; Li, Y.Q.; Setlow, P. Analysis of the effects of a gerP mutation on the germination of spores of Bacillus subtilis. J. Bacteriol. 2012, 194, 5749–5758. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.G.; Moran, C.P.; Henriques, A.O. Autoregulation of SafA assembly through recruitment of a protein cross-linking enzyme. J. Bacteriol. 2018, 200, e00066-18. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Lysozyme | H2O2 | ||||
|---|---|---|---|---|---|---|
| Spore count (CFU/mL) | Survival (%) | Spore Count (CFU/mL) | Survival (%) | |||
| − | + | − | + | |||
| PY79 | 1.1 × 108 | 2.6 × 107 | 24 | 4.2 × 107 | 1.5 × 107 | 36 |
| ΔyncD | 6.6 × 107 | 1.3 × 107 | 20 | 7.8 × 107 | 1.3 × 107 | 17 |
| Strain | Temperature | Prep 1 | Prep 2 | Prep 3 | |||
|---|---|---|---|---|---|---|---|
| D-Value (min) | R2 | D-Value (min) | R2 | D-Value (min) | R2 | ||
| PY79 | 100 °C | 5.4 | 0.96 | 5.6 | 0.90 | 12.9 | 0.87 |
| 110 °C | 1.2 | 0.87 | 0.4 | 0.81 | 0.4 | 0.78 | |
| ΔyncD | 100 °C | 2.9 | 0.94 | 2.5 | 0.90 | 4.2 | 0.94 |
| 110 °C | 0.6 | 0.97 | 0.6 | 0.93 | 0.5 | 0.80 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krajčíková, D.; Bugárová, V.; Barák, I. Interactions of Bacillus subtilis Basement Spore Coat Layer Proteins. Microorganisms 2021, 9, 285. https://doi.org/10.3390/microorganisms9020285
Krajčíková D, Bugárová V, Barák I. Interactions of Bacillus subtilis Basement Spore Coat Layer Proteins. Microorganisms. 2021; 9(2):285. https://doi.org/10.3390/microorganisms9020285
Chicago/Turabian StyleKrajčíková, Daniela, Veronika Bugárová, and Imrich Barák. 2021. "Interactions of Bacillus subtilis Basement Spore Coat Layer Proteins" Microorganisms 9, no. 2: 285. https://doi.org/10.3390/microorganisms9020285