
The Survival of Haloferax mediterranei under Stressful Conditions

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Media
2.2. Stress Conditions
2.3. Specific Growth Rate and Doubling Time
2.4. Statistical Analysis
2.5. Sample Treatment for Inductively Coupled Plasma Mass Spectrometry (ICP-MS)
2.6. Quantification of Metal Content by ICP-MS
3. Results and Discussion
3.1. Salinity Stress
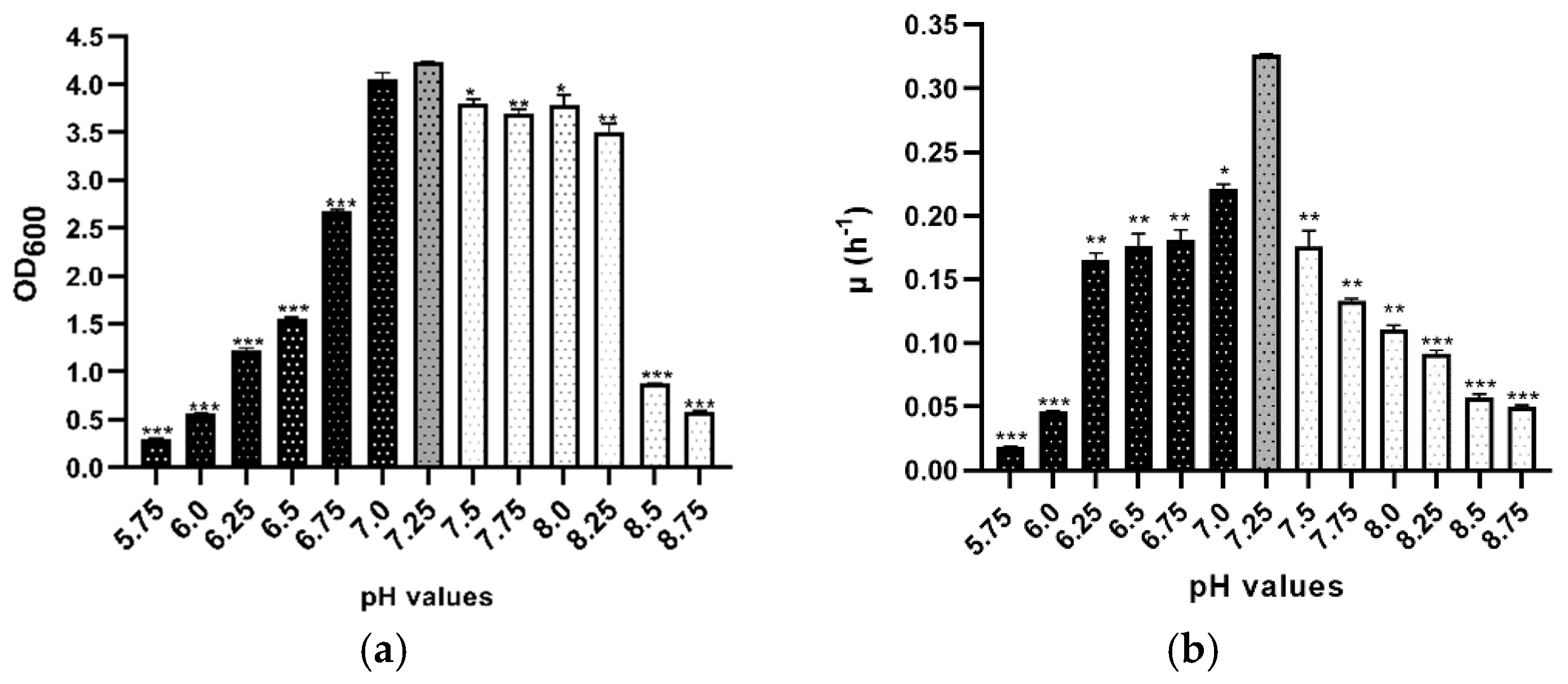
3.2. pH Stress
3.3. Temperature Stress
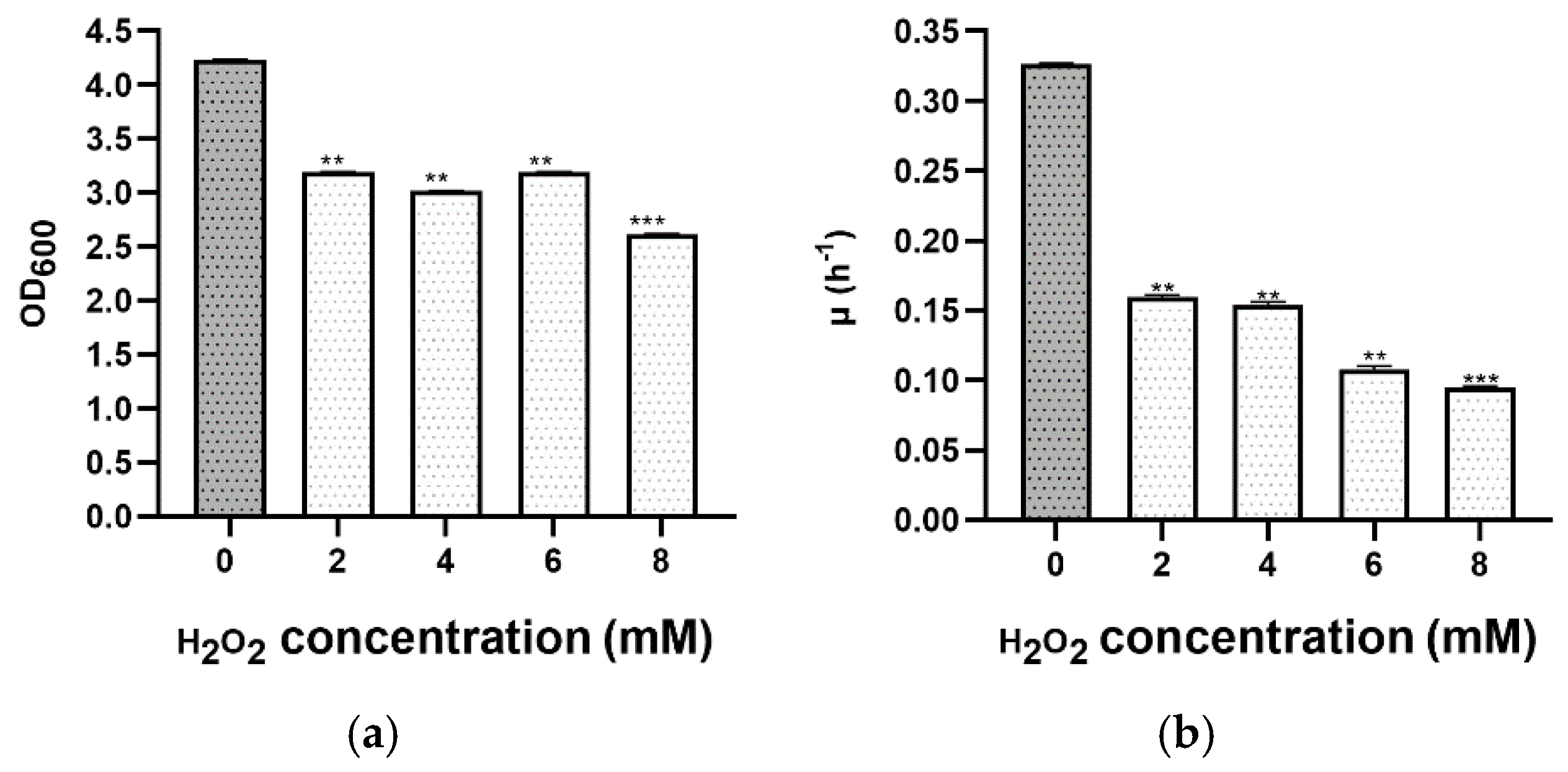
3.4. Oxidative Stress
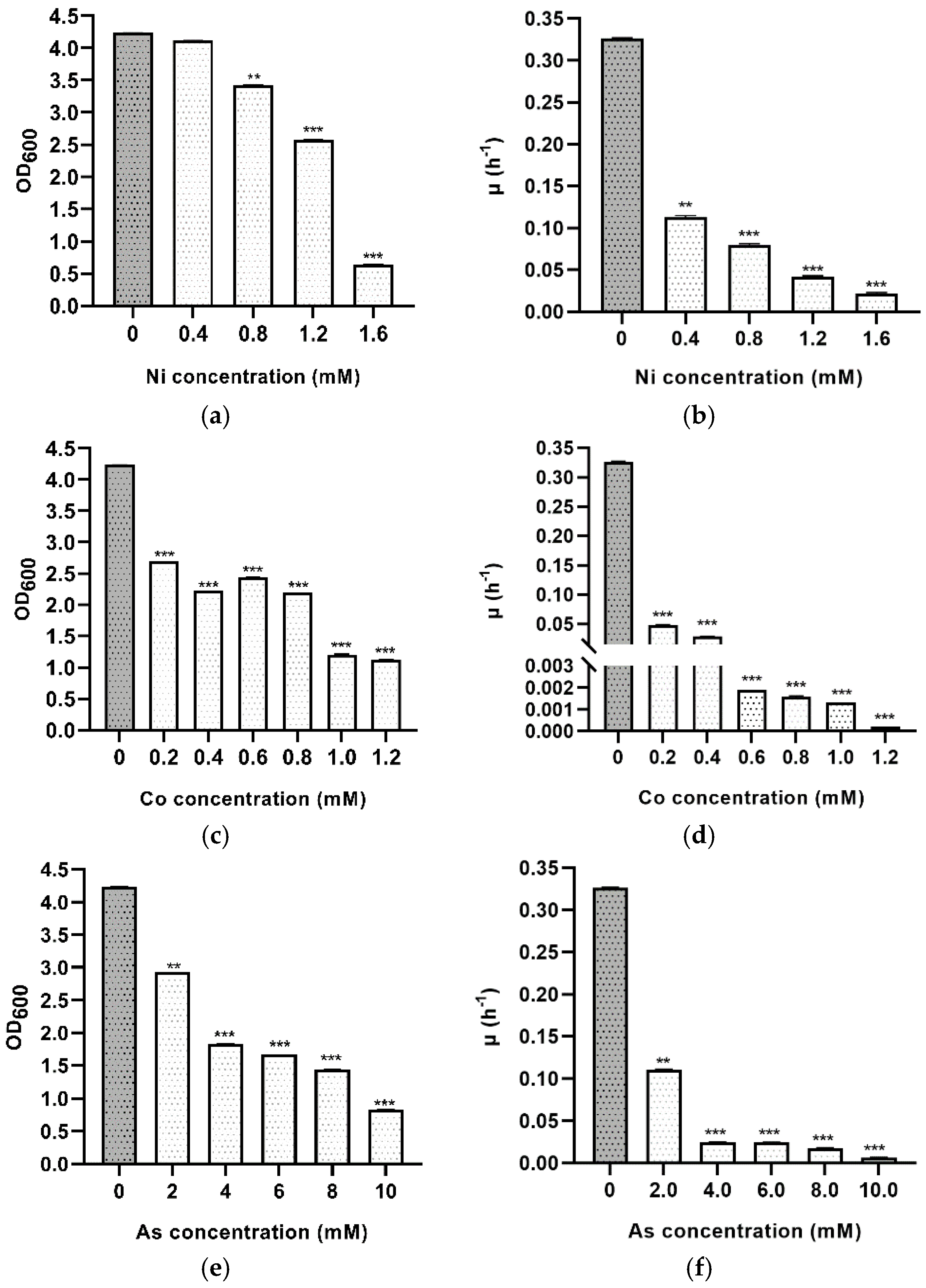
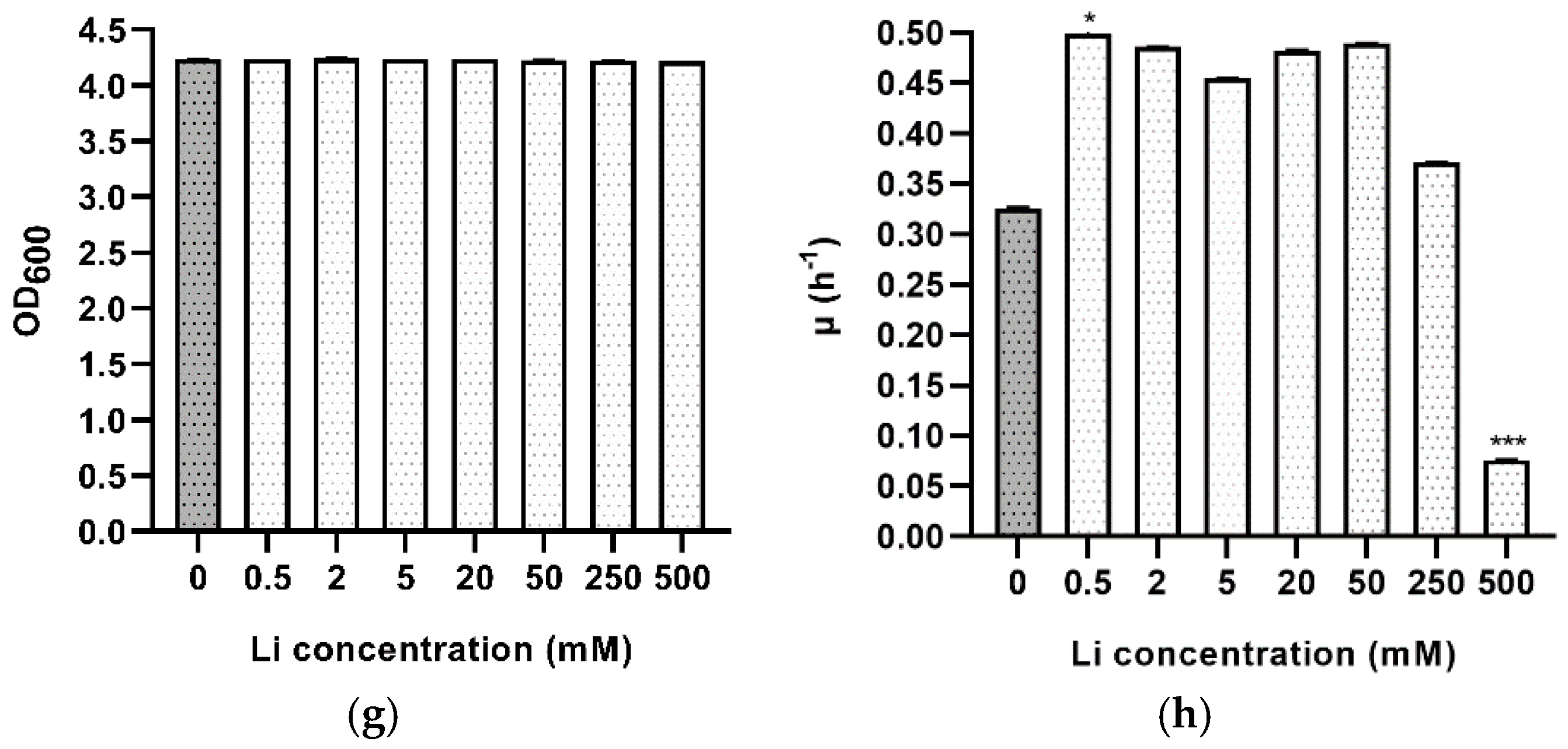
3.5. Metal Stress
3.6. Detection of Intracellular Metal Ion Concentration Using ICP-MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- de Macario, E.C.; Macario, A.J.L. Molecular biology of stress genes in methanogens: Potential for bioreactor technology. Adv. Biochem. Eng. Biotechnol. 2003, 81, 95–150. [Google Scholar] [CrossRef]
- de Macario, E.C.; Macario, A.J. Stressors, stress and survival: Overview. Front. Biosci. 2000, 5, d780–d786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quehenberger, J.; Shen, L.; Albers, S.V.; Siebers, B.; Spadiut, O. Sulfolobus—A potential key organism in future biotechnology. Front. Microbiol. 2017, 8, 2474. [Google Scholar] [CrossRef] [PubMed]
- Gumulya, Y.; Boxall, N.J.; Khaleque, H.N.; Santala, V.; Carlson, R.P.; Kaksonen, A.H. In a quest for engineering acidophiles for biomining applications: Challenges and opportunities. Genes 2018, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, E.R.; Castro, C.; Urbieta, M.S. Thermophilic microorganisms in biomining. World J. Microbiol. Biotechnol. 2016, 32, 179. [Google Scholar] [CrossRef]
- Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lledó, B.; Martínez-Espinosa, R.M.; Marhuenda-Egea, F.C.; Bonete, M.J. Respiratory nitrate reductase from haloarchaeon Haloferax mediterranei: Biochemical and genetic analysis. Biochim. Biophys. Acta Gen. Subj. 2004, 1674, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Andrei, A.Ş.; Banciu, H.L.; Oren, A. Living with salt: Metabolic and phylogenetic diversity of archaea inhabiting saline ecosystems. FEMS Microbiol. Lett. 2012, 330, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Valera, F.; Juez, G.; Kushner, D.J. Halobacterium mediterranei spec, nov., a New Carbohydrate-Utilizing Extreme Halophile. Syst. Appl. Microbiol. 1983, 4, 369–381. [Google Scholar] [CrossRef]
- Robinson, J.L.; Pyzyna, B.; Atrasz, R.G.; Henderson, C.A.; Morrill, K.L.; Burd, A.M.; Desoucy, E.; Fogleman Iii, R.E.; Naylor, J.B.; Steele, S.M.; et al. Growth Kinetics of Extremely Halophilic Archaea (Family Halobacteriaceae) as Revealed by Arrhenius Plots. J. Bacteriol. 2005, 187, 923–929. [Google Scholar] [CrossRef] [Green Version]
- Shand, R.F.; Perez, A.M. Haloarchaeal Growth Physiology. In Enigmatic Microorganisms and Life in Extreme Environments; Springer: Dordrecht, The Netherlands, 1999; pp. 411–424. ISBN 978-94-011-4838-2. [Google Scholar]
- D’Souza, S.E.; Altekar, W.; D’Souza, S.F. Adaptive response of Haloferax mediterranei to low concentrations of NaCl (<20%) in the growth medium. Arch. Microbiol. 1997, 168, 68–71. [Google Scholar] [CrossRef]
- Oren, A.; Hallsworth, J.E. Microbial weeds in hypersaline habitats: The enigma of the weed-like Haloferax mediterranei. FEMS Microbiol. Lett. 2014, 359, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; Martínez-Espinosa, R.M.; Esclapez, J.; Bautista, V.; Pire, C.; Camacho, M.; Richardson, D.J.; Bonete, M.J. Anaerobic Metabolism in Haloferax Genus: Denitrification as Case of Study. Adv. Microb. Physiol. 2016, 68, 41–85. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.J.; Martínez-Espinosa, R.M. Carotenoids from Haloarchaea and their potential in biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [Green Version]
- Mainka, T.; Mahler, N.; Herwig, C.; Pflügl, S. Soft sensor-based monitoring and efficient control strategies of biomass concentration for continuous cultures of Haloferax mediterranei and their application to an industrial production chain. Microorganisms 2019, 7, 648. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; Pire, C.; Martinez-Espinosa, R.M.; Bergaust, L. Denitrifying haloarchaea within the genus Haloferax display divergent respiratory phenotypes, with implications for their release of nitrogenous gases. Environ. Microbiol. 2019, 21, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhuang, X.; Ahmad, S.; Sung, S.; Ni, S.Q. Biotreatment of high-salinity wastewater: Current methods and future directions. World J. Microbiol. Biotechnol. 2020, 36, 37. [Google Scholar] [CrossRef]
- Nájera-Fernández, C.; Zafrilla, B.; Bonete, M.J.; Martínez-Espinosa, R.M. Role of the denitrifying Haloarchaea in the treatment of nitrite-brines. Int. Microbiol. 2012, 15, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thombre, R.S.; Shinde, V.D.; Oke, R.S.; Dhar, S.K.; Shouche, Y.S. Biology and survival of extremely halophilic archaeon Haloarcula marismortui RR12 isolated from Mumbai salterns, India in response to salinity stress. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Amelin, E.; Pindado, P. The challenge of climate change in Spain: Water resources, agriculture and land. J. Hydrol. 2014, 518, 243–249. [Google Scholar] [CrossRef]
- Domínguez, M.; Romera, R.; Sánchez, E.; Fita, L.; Fernández, J.; Jiménez-Guerrero, P.; Montávez, J.P.; Cabos, W.D.; Liguori, G.; Gaertner, M.Á. Present-climate precipitation and temperature extremes over Spain from a set of high resolution RCMs. Clim. Res. 2013, 58, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 8707. [Google Scholar] [CrossRef]
- Estrada, F.; Tol, R.S.J.; Botzen, W.J.W. Global economic impacts of climate variability and change during the 20th century. PLoS ONE 2017, 12, e0172201. [Google Scholar] [CrossRef]
- Crist, E.; Mora, C.; Engelman, R. The interaction of human population, food production, and biodiversity protection. Science 2017, 356, 260–264. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai914. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D.; Stan-Lotter, H. Origins of halophilic microorganisms in ancient salt deposits. Environ. Microbiol. 2000, 2, 243–250. [Google Scholar] [CrossRef]
- Rodríguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Isolation of Extremely Halophilic Bacteria Able to Grow in Defined Inorganic Media with Single Carbon Sources. J. Gen. Microbiol. 1980, 119, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Bowers, K.J.; Wiegel, J. Temperature and pH optima of extremely halophilic archaea: A mini-review. Extremophiles 2011, 15, 119–128. [Google Scholar] [CrossRef]
- Coker, J.A.; Moran-Reyna, A. The effects of extremes of pH on the growth and transcriptomic profiles of three haloarchaea. F1000Research 2014, 3, 168. [Google Scholar] [CrossRef] [Green Version]
- Coker, J.A.; DasSarma, P.; Kumar, J.; Müller, J.A.; DasSarma, S. Transcriptional profiling of the model Archaeon Halobacterium sp. NRC-1: Responses to changes in salinity and temperature. Saline Syst. 2007, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Gillum, N.; Boyd, J.L.; Schmid, A. The RosR transcription factor is required for gene expression dynamics in response to extreme oxidative stress in a hypersaline-adapted archaeon. BMC Genom. 2012, 13, 351. [Google Scholar] [CrossRef] [Green Version]
- Voica, D.M.; Bartha, L.; Banciu, H.L.; Oren, A. Heavy metal resistance in halophilic Bacteria and Archaea. FEMS Microbiol. Lett. 2016, 363, fnw146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Völkel, S.; Fröls, S.; Pfeifer, F. Heavy metal ion stress on Halobacterium salinarum R1 planktonic cells and biofilms. Front. Microbiol. 2018, 9, 3157. [Google Scholar] [CrossRef] [Green Version]
- Baati, H.; Siala, M.; Azri, C.; Ammar, E.; Dunlap, C.; Trigui, M. Resistance of a Halobacterium salinarum isolate from a solar saltern to cadmium, lead, nickel, zinc, and copper. Antonie Van Leeuwenhoek 2020, 113, 1699–1711. [Google Scholar] [CrossRef]
- Nieto, J.J.; Ventosa, A.; Ruiz-Berraquero, F. Susceptibility of Halobacteria to Heavy Metals. Appl. Environ. Microbiol. 1987, 53, 1199–1202. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, P.; Kowshik, M. Mechanisms of metal resistance and homeostasis in Haloarchaea. Archaea 2013, 2013, 732864. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jin, Y.; Zhao, J.; Liu, N.; Cui, Y. Nickel-hydrogen batteries for large-scale energy storage. Proc. Natl. Acad. Sci. USA 2018, 115, 11694–11699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippers, A.; Hedrich, S.; Vasters, J.; Drobe, M.; Sand, W.; Willscher, S. Biomining: Metal recovery from ores with microorganisms. Adv. Biochem. Eng. Biotechnol. 2014, 141, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Tsuruta, T. Removal and recovery of lithium using various microorganisms. J. Biosci. Bioeng. 2005, 100, 562–566. [Google Scholar] [CrossRef]
- Owen, J.R. Rechargeable lithium batteries. Chem. Soc. Rev. 1997, 26, 259–267. [Google Scholar] [CrossRef]
- Wakihara, M. Recent developments in lithium ion batteries. Mater. Sci. Eng. R Rep. 2001, 33, 109–134. [Google Scholar] [CrossRef]
- Belfiore, C.; Curia, M.V.; Farías, M.E. Characterization of Rhodococcus sp. A5wh isolated from a high altitude Andean lake to unravel the survival strategy under lithium stress. Rev. Argent Microbiol. 2018, 50, 311–322. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| %SW | d.t. (h) * | p-Value | Summary |
|---|---|---|---|
| 10 | 14.8 ± 0.5 | <0.001 | *** |
| 12.5 | 9.1 ± 0.2 | <0.001 | *** |
| 15 | 3.4 ± 0.2 | 0.037 | * |
| 17.5 | 2.7 ± 0.2 | 0.148 | ns |
| 20 | 2.15 ± 0.03 | Control | |
| 22.5 | 3.0 ± 0.3 | 0.138 | ns |
| 25 | 3.3 ± 0.2 | 0.037 | * |
| 27.5 | 7.2 ± 0.1 | <0.001 | *** |
| 30 | 6.91 ± 0.09 | <0.001 | *** |
| 32.5 | 7.75 ± 0.05 | <0.001 | *** |
| pH | d.t. (h) * | p-Value | Summary |
|---|---|---|---|
| 5.75 | 36.9 ± 0.3 | <0.001 | *** |
| 6.0 | 15.0 ± 0.3 | <0.001 | *** |
| 6.25 | 3.9 ± 0.1 | 0.006 | ** |
| 6.5 | 4.2 ± 0.1 | 0.003 | ** |
| 6.75 | 3.13 ± 0.05 | 0.03 | ** |
| 7.0 | 3.8 ± 0.2 | 0.004 | * |
| 7.25 | 2.15 ± 0.03 | - | Control |
| 7.5 | 5.22 ± 0.09 | 0.001 | ** |
| 7.75 | 6.3 ± 0.2 | 0.003 | ** |
| 8.0 | 4.0 ± 0.3 | 0.003 | ** |
| 8.25 | 7.6 ± 0.2 | <0.001 | *** |
| 8.5 | 12.1 ± 0.2 | <0.001 | *** |
| 8.75 | 14.0 ± 0.3 | <0.001 | *** |
| Temperature (°C) | d.t. (h) * | p-Value | Summary |
|---|---|---|---|
| 32 | 31.8 ± 0.4 | <0.001 | *** |
| 37 | 23.0 ± 0.4 | <0.001 | *** |
| 42 | 2.13 ± 0.03 | - | Control |
| 47 | 3.65 ± 0.04 | 0.004 | ** |
| 52 | 3.75 ± 0.03 | 0.003 | ** |
| [H2O2] (mM) | d.t. (h) * | p-Value | Summary |
|---|---|---|---|
| 0 | 2.13 ± 0.03 | - | Control |
| 2 | 4.34 ± 0.04 | 0.003 | ** |
| 4 | 4.51± 0.08 | 0.003 | ** |
| 6 | 6.4 ± 0.1 | 0.001 | ** |
| 8 | 7.3 ± 0.1 | <0.001 | *** |
| Heavy Metal | Concentration (mM) | d.t. (h) * | p-Value | Summary |
|---|---|---|---|---|
| - | - | 2.15 ± 0.03 | - | Control |
| Nickel | 0.5 | 6.14 ± 0.09 | 0.001 | ** |
| 0.8 | 8.7 ± 0.2 | <0.001 | *** | |
| 1.2 | 16.6 ± 0.4 | <0.001 | *** | |
| 1.6 | 31.9 ± 1.6 | <0.001 | *** | |
| Cobalt | 0.2 | 14.24 ± 0.06 | <0.001 | *** |
| 0.4 | 24.1 ± 0.2 | <0.001 | *** | |
| 0.6 | 364.81 ± 0.01 | <0.001 | *** | |
| 0.8 | 442.84 ± 0.02 | <0.001 | *** | |
| 1 | 533.19 ± 0.01 | <0.001 | *** | |
| 1.2 | 3465.73 ± 0.02 | <0.001 | *** | |
| Arsenic | 2 | 6.28 ± 0.03 | 0.001 | ** |
| 4 | 28.1 ± 0.2 | <0.001 | *** | |
| 6 | 28.5 ± 0.2 | <0.001 | *** | |
| 8 | 40 ± 1 | <0.001 | *** | |
| 10 | 105 ± 2 | <0.001 | *** | |
| Lithium | 0.5 | 1.390 ± 0.004 | 0.13 | * |
| 2 | 1.426 ± 0.003 | 0.263 | ns | |
| 5 | 1.523 ± 0.001 | >0.999 | ns | |
| 20 | 1.437 ± 0.004 | 0.832 | ns | |
| 50 | 1.416 ± 0.002 | 0.065 | ns | |
| 250 | 1.865 ± 0.002 | >0.999 | ns | |
| 500 | 9.11 ± 0.09 | <0.001 | *** |
| Medium | Cobalt (mg/Kg) | Nickel (mg/Kg) | Lithium (mg/Kg) | Arsenic (mg/Kg) |
|---|---|---|---|---|
| Control | 0.00 ± 0.00 | 0.17 ± 0.00 | 0.50 ± 0.00 | 0.00 ± 0.00 |
| Co 0.2 mM | 1.48 ± 0.01 | - | - | - |
| Co 1.2 mM | 2.41 ± 0.09 | - | - | - |
| As 4 mM | - | - | - | 0.17 ± 0.09 |
| Ni 0.5 mM | - | 0.32 ± 0.01 | - | - |
| Ni 1.2 mM | - | 1.37 ± 0.15 | - | - |
| Li 12 mM | - | - | 0.64 ± 0.01 | - |
| Li 50 mM | - | - | 0.66 ± 0.01 | - |
| Li 250 mM | - | - | 1.56 ± 0.02 | - |
| Li 500 mM | - | - | 1.55 ± 0.02 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matarredona, L.; Camacho, M.; Zafrilla, B.; Bravo-Barrales, G.; Esclapez, J.; Bonete, M.-J. The Survival of Haloferax mediterranei under Stressful Conditions. Microorganisms 2021, 9, 336. https://doi.org/10.3390/microorganisms9020336
Matarredona L, Camacho M, Zafrilla B, Bravo-Barrales G, Esclapez J, Bonete M-J. The Survival of Haloferax mediterranei under Stressful Conditions. Microorganisms. 2021; 9(2):336. https://doi.org/10.3390/microorganisms9020336
Chicago/Turabian StyleMatarredona, Laura, Mónica Camacho, Basilio Zafrilla, Gloria Bravo-Barrales, Julia Esclapez, and María-José Bonete. 2021. "The Survival of Haloferax mediterranei under Stressful Conditions" Microorganisms 9, no. 2: 336. https://doi.org/10.3390/microorganisms9020336