
CD4+ and CD8+ Circulating Memory T Cells Are Crucial in the Protection Induced by Vaccination with Salmonella Typhi Porins

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bacteria
2.3. S. Typhi Porins
2.4. Vaccination
2.5. Delayed-Type Hypersensivity (DTH)
2.6. T Cell Extraction and Stimulation
2.7. T Cell Immunophenotyping
2.8. IgG and IgM Antibody Titers
2.9. CD4 and CD8 Depletion
2.10. Challenge with Bacteria and CFU Count
2.11. Survival Test and Clinical Score
3. Results
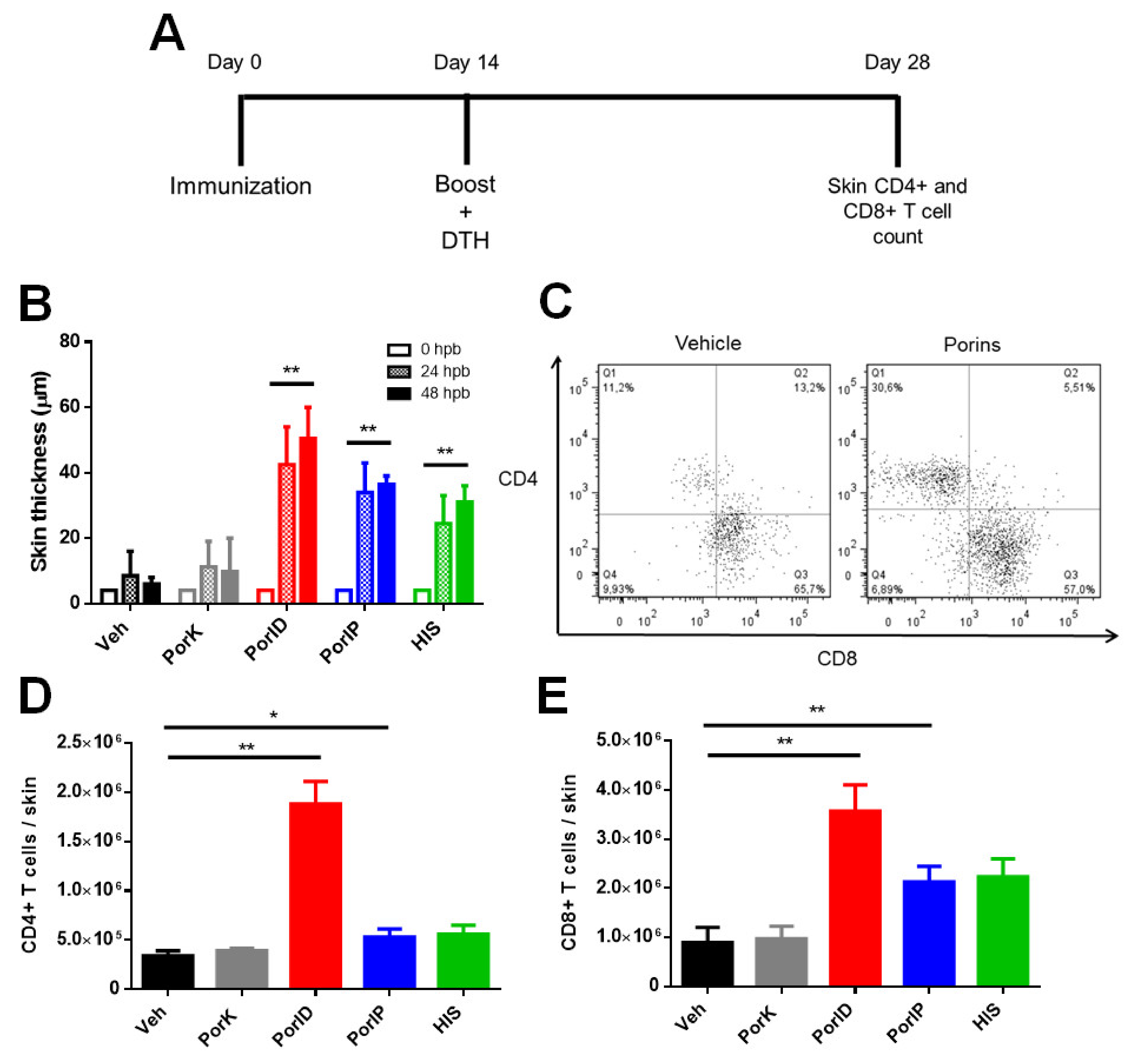
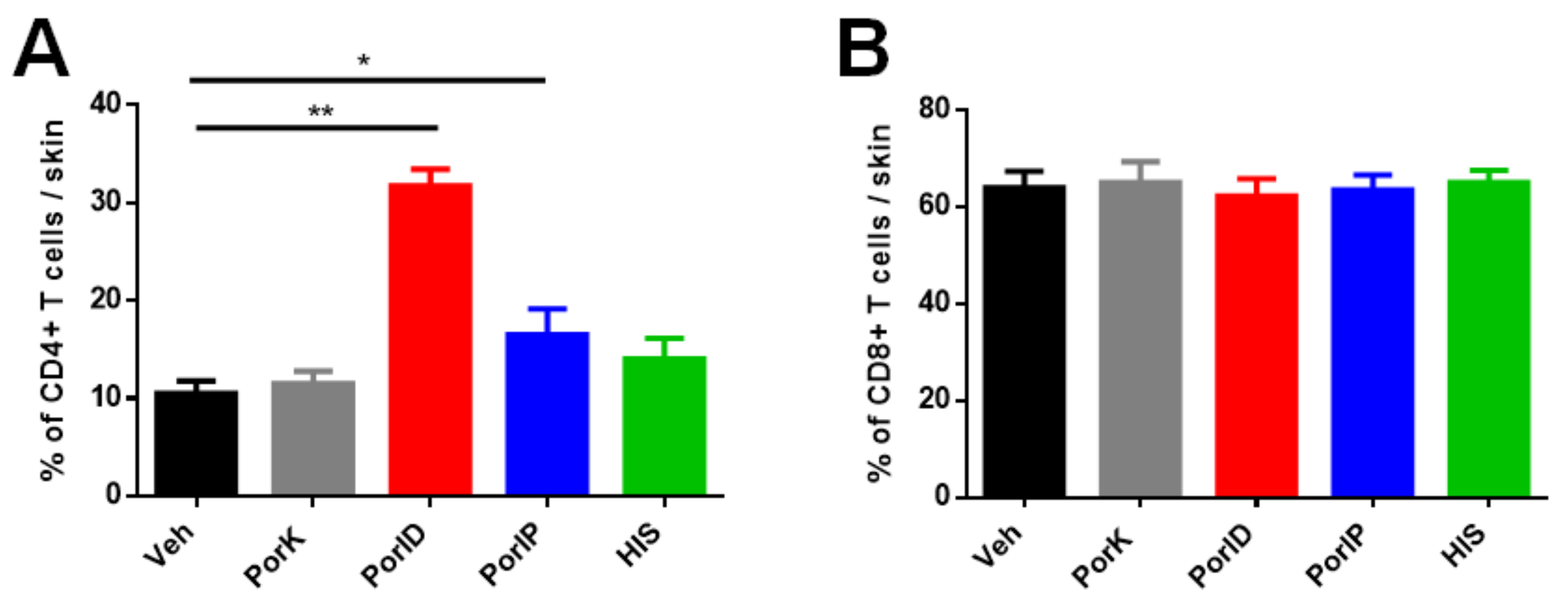
3.1. Vaccination with S. Typhi Porins Induces CD4+ and CD8+ T Cell Responses
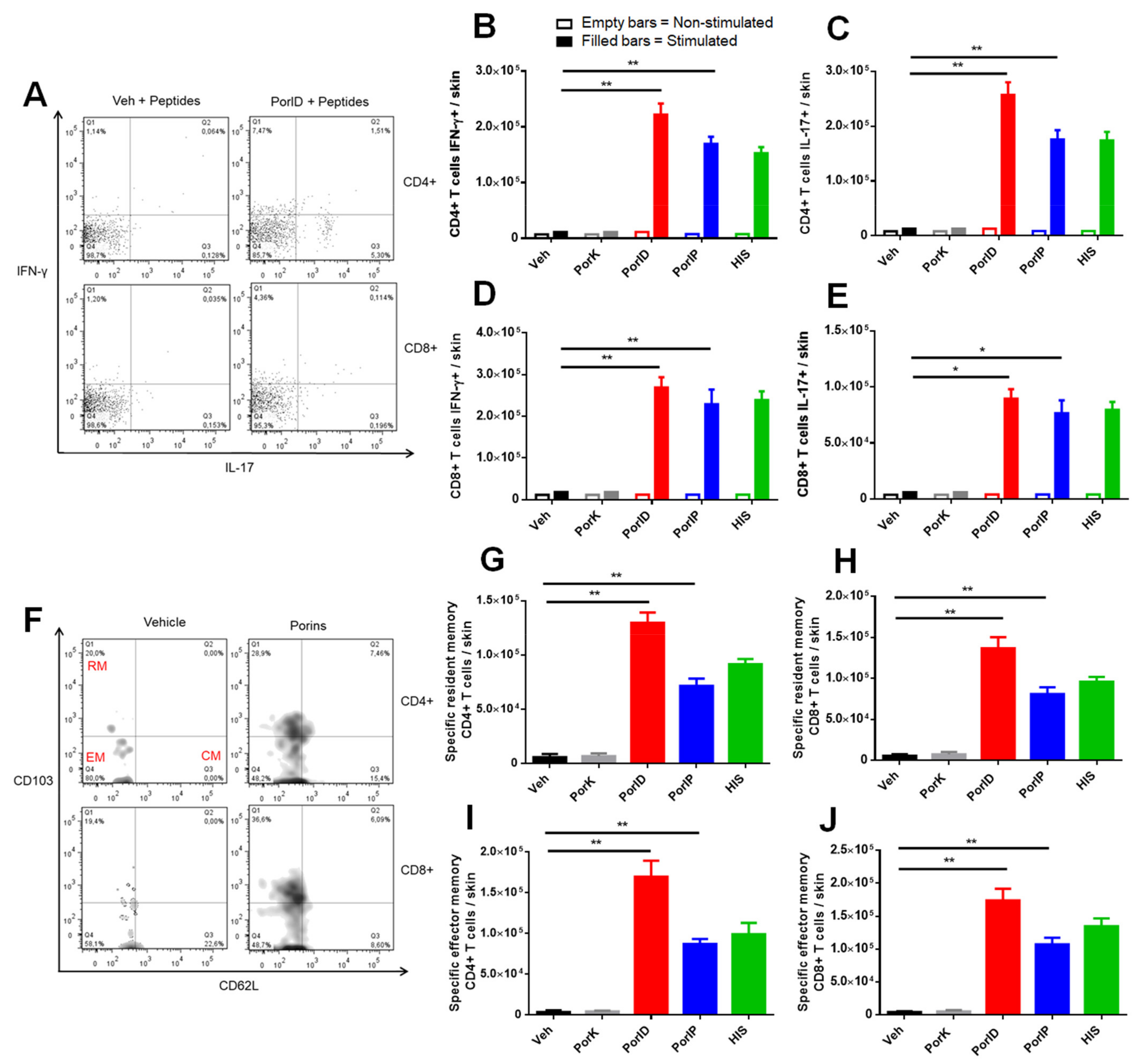
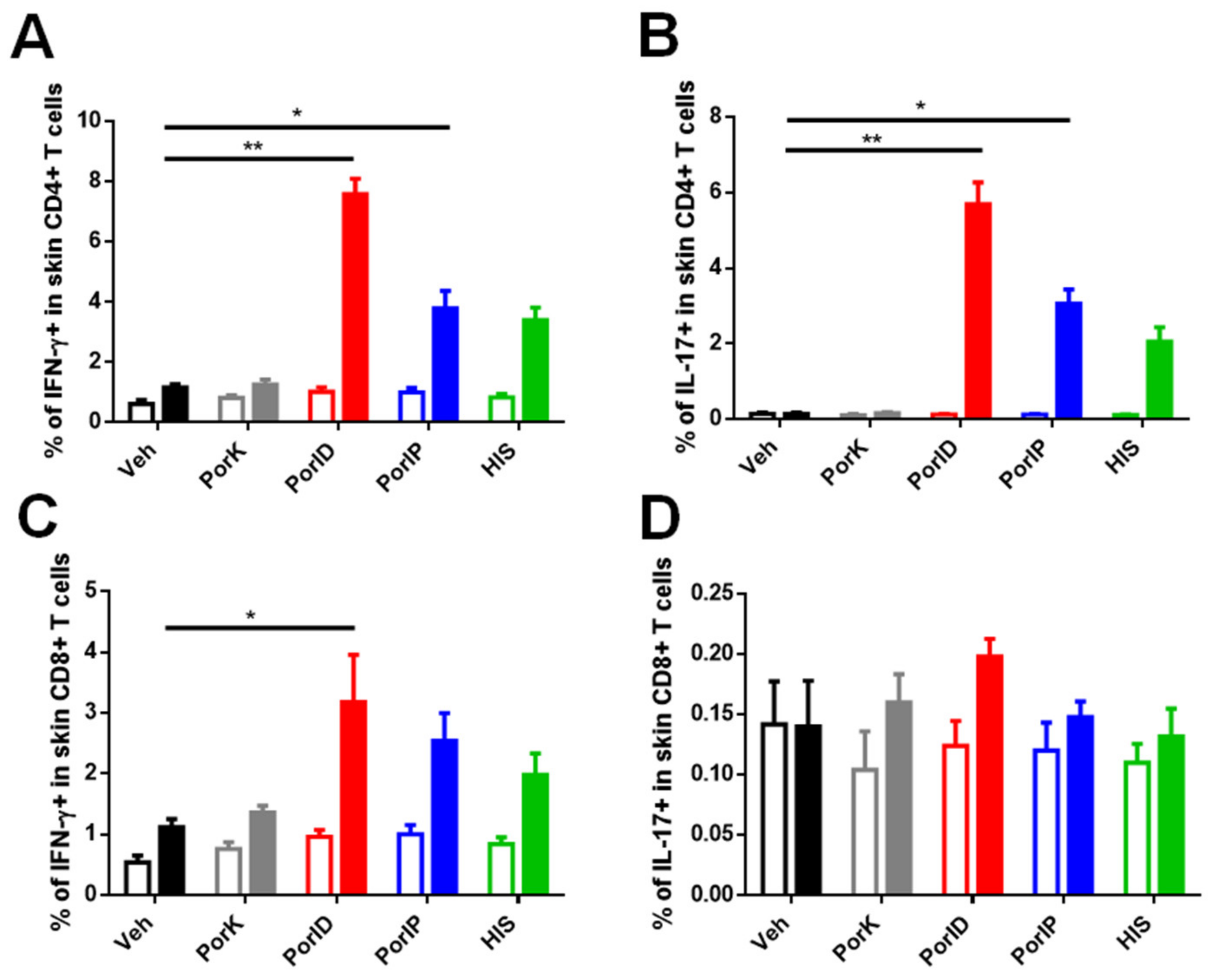
3.2. Vaccination with S. Typhi Porins Induces Functional Resident and Effector Memory CD4+ and CD8+ T Cell Responses
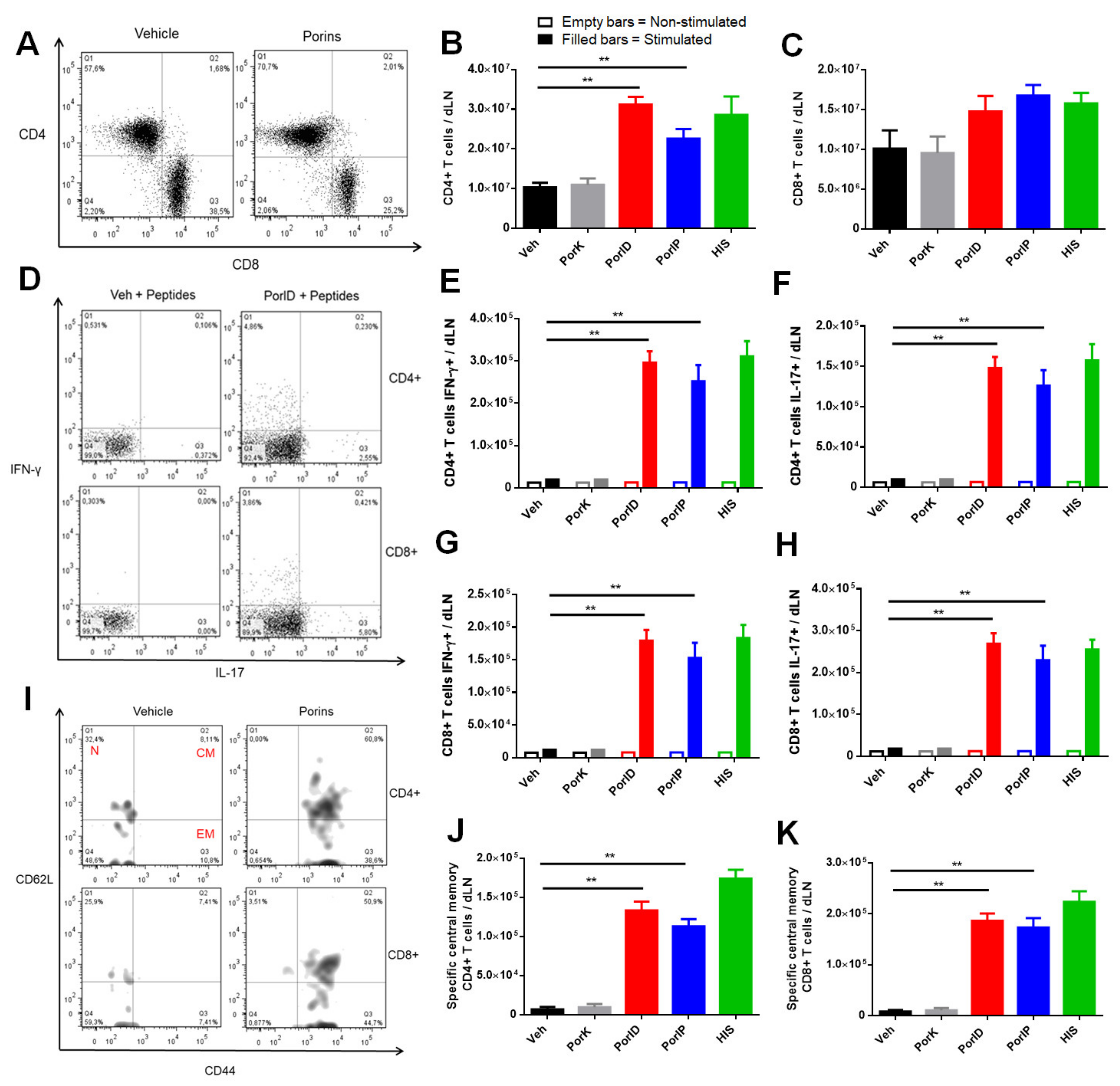
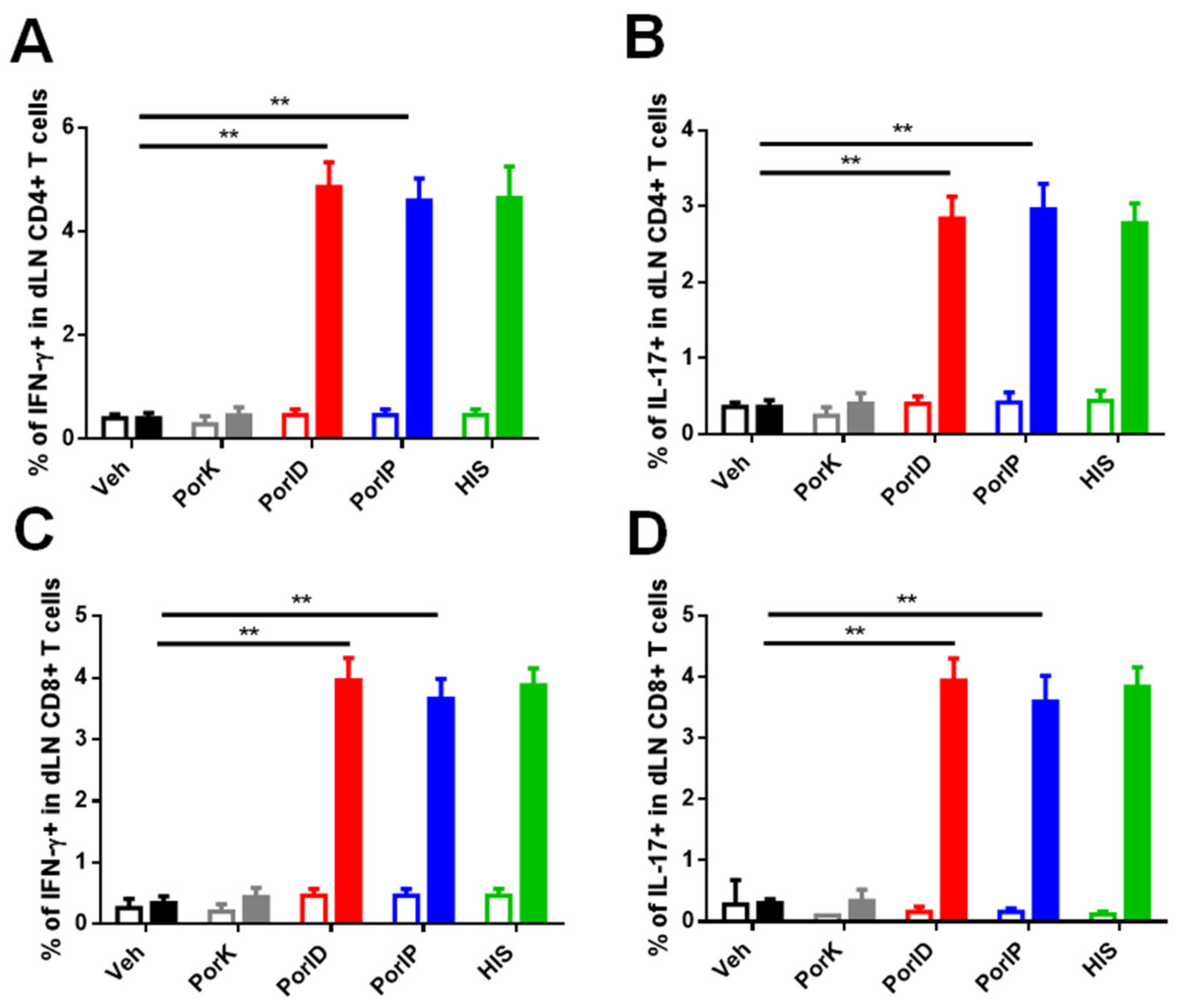
3.3. Vaccination with S. Typhi Porins Induces Functional Central Memory CD4+ and CD8+ T Cell Responses
3.4. Vaccination with S. Typhi Porins Reduces the Bacterial Burden Following Challenge
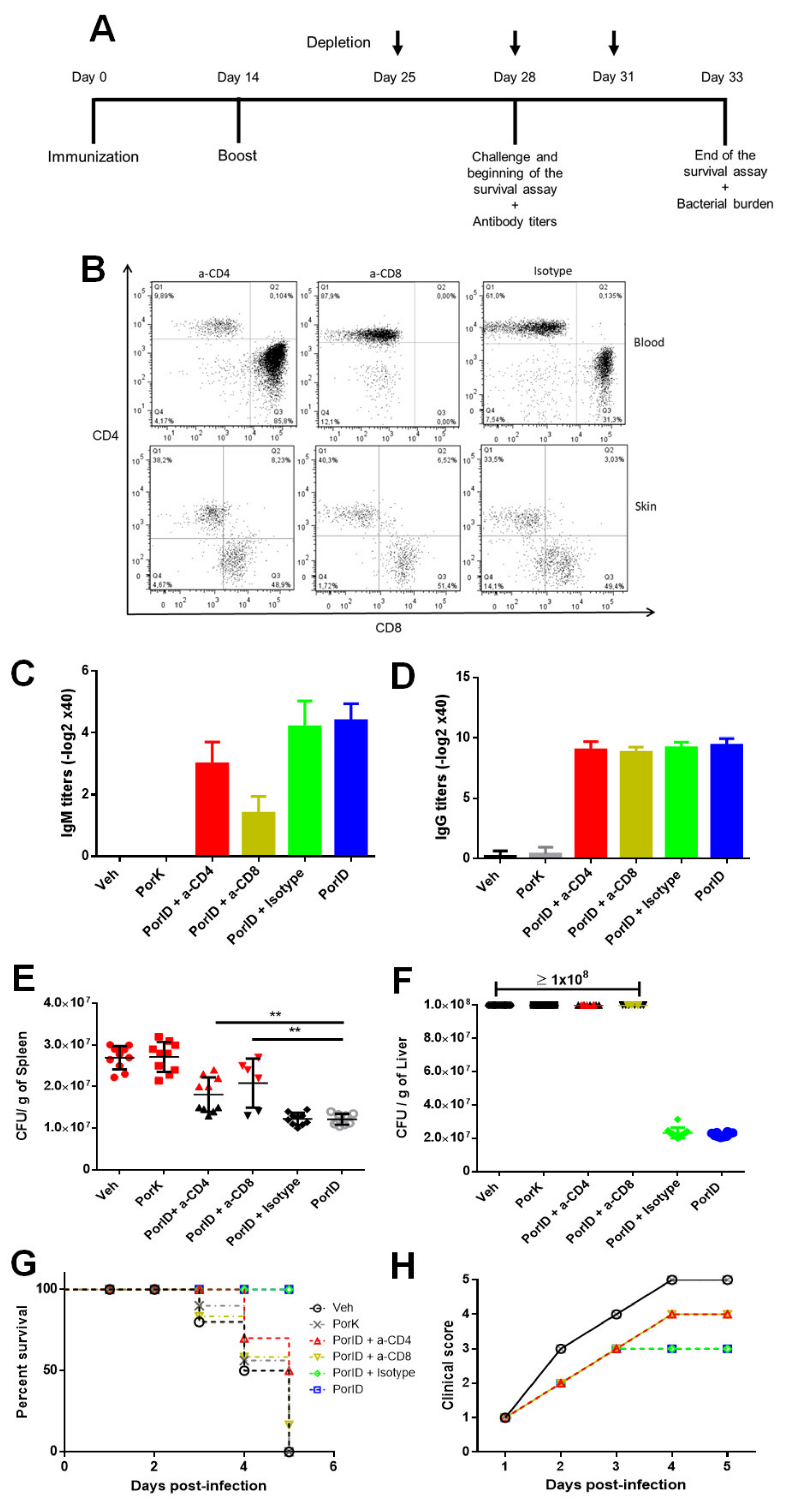
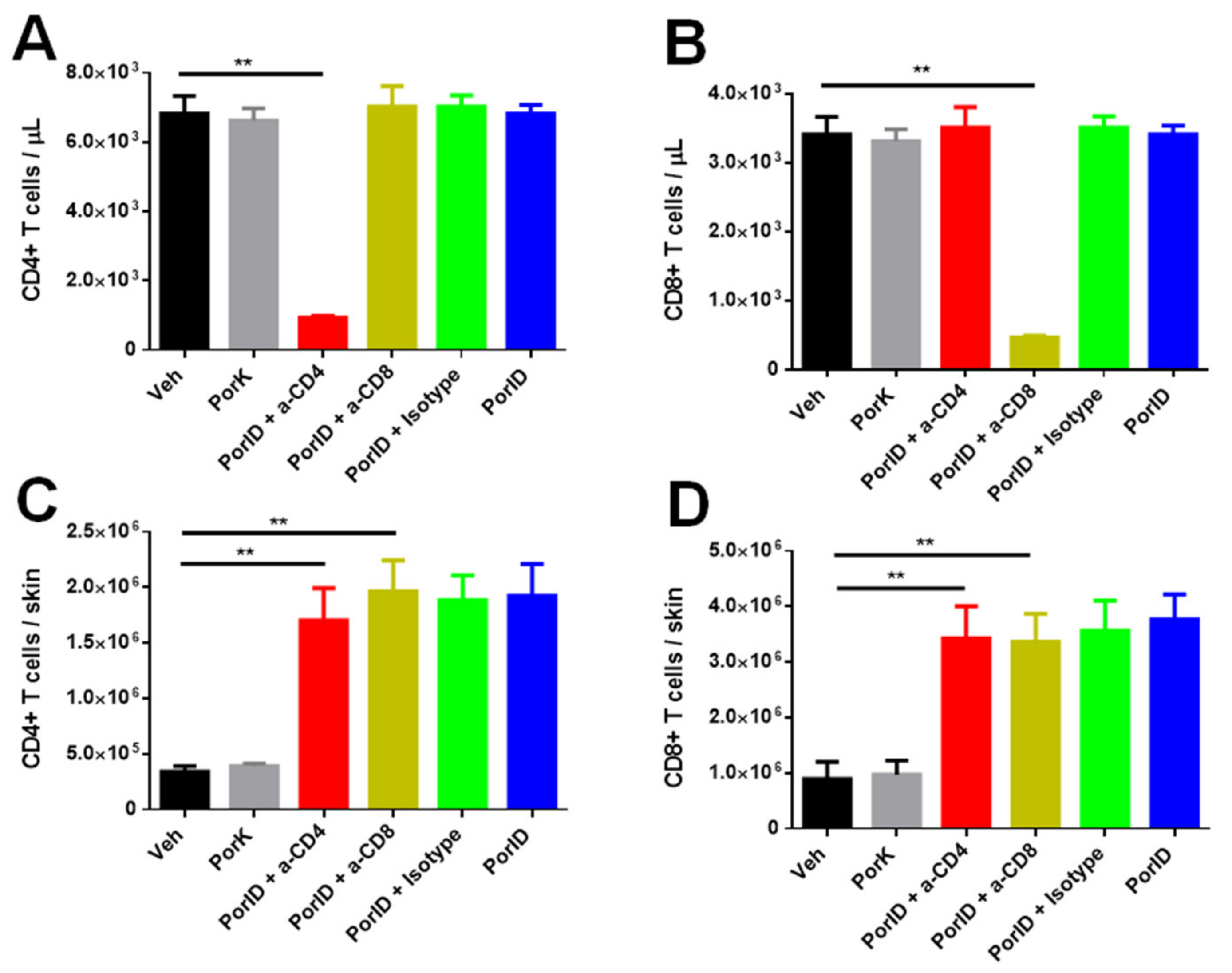
3.5. Circulating Memory CD4+ and CD8+ T Cells are Crucial in the Protection Induced by S. Typhi Porins
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




References
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://doi.org/10.1590/S0100-15742013000100018 (accessed on 3 February 2021).
- Stanaway, J.D.; Parisi, A.; Sarkar, K.; Blacker, B.F.; Reiner, R.C.; Hay, S.I.; Nixon, M.R.; Dolecek, C.; James, S.L.; Mokdad, A.H.; et al. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef] [Green Version]
- Gayet, R.; Bioley, G.; Rochereau, N.; Paul, S.; Corthésy, B. Vaccination against Salmonella Infection: The Mucosal Way. Microbiol. Mol. Biol. Rev. 2017, 81. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, V.; Isibasi, A.; Garcia-Ortigoza, E.; Kumate, J. Immunoblot detection of class-specific humoral immune response to outer membrane proteins isolated from Salmonella typhi in humans with typhoid fever. J. Clin. Microbiol. 1989, 27, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Isibasi, A.; Ortiz-Navarrete, V.; Paniagua, J.; Pelayo, R.; González, C.R.; García, J.A.; Kumate, J. Active protection of mice against Salmonella typhi by immunization with strain-specific porins. Vaccine 1992, 10, 811–813. [Google Scholar] [CrossRef]
- Secundino, I.; López-Macías, C.; Cervantes-Barragán, L.; Gil-Cruz, C.; Ríos-Sarabia, N.; Pastelin-Palacios, R.; Villasis-Keever, M.A.; Becker, I.; Puente, J.L.; Calva, E.; et al. Salmonella porins induce a sustained, lifelong specific bactericidal antibody memory response. Immunology 2006, 117, 59–70. [Google Scholar] [CrossRef]
- Perez-Shibayama, C.; Gil-Cruz, C.; Pastelin-Palacios, R.; Cervantes-Barragan, L.; Hisaki, E.; Chai, Q.; Onder, L.; Scandella, E.; Regen, T.; Waisman, A.; et al. IFN-γ–Producing CD4 + T Cells Promote Generation of Protective Germinal Center–Derived IgM + B Cell Memory against Salmonella Typhi. J. Immunol. 2014, 192, 5192–5200. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Toledo, M.; Valero-Pacheco, N.; Pastelin-Palacios, R.; Gil-Cruz, C.; Perez-Shibayama, C.; Moreno-Eutimio, M.A.; Becker, I.; Pérez-Tapia, S.M.; Arriaga-Pizano, L.; Cunningham, A.F.; et al. Salmonella Typhi porins OmpC and OmpF are potent adjuvants for T-dependent and T-independent antigens. Front. Immunol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Carrenõ, J.M.; Perez-Shibayama, C.; Gil-Cruz, C.; Lopez-Macias, C.; Vernazza, P.; Ludewig, B.; Albrich, W.C. Evolution of Salmonella Typhi outer membrane protein-specific T and B cell responses in humans following oral Ty21a vaccination: A randomized clinical trial. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Blanco, F.; Isibasi, A.; González, C.R.; Ortiz, V.; Paniagua, J.; Arreguín, C.; Kumate, J. Human cell mediated immunity to porins from Salmonella typhi. Scan. J. Infect. Dis. 1993, 25, 73–80. [Google Scholar] [CrossRef]
- Diaz-Quiñonez, A.; Martin-Orozco, N.; Isibasi, A.; Ortiz-Navarrete, V. Two Salmonella OmpC Kb-Restricted Epitopes for CD8 +-T-Cell Recognition. Infect. Immun. 2004, 72, 3059–3062. [Google Scholar] [CrossRef] [Green Version]
- Valero-Pacheco, N.; Blight, J.; Aldapa-Vega, G.; Kemlo, P.; Pérez-Toledo, M.; Wong-Baeza, I.; Kurioka, A.; Perez-Shibayama, C.; Gil-Cruz, C.; Sánchez-Torres, L.E.; et al. Conservation of the OmpC Porin Among Typhoidal and Non-Typhoidal Salmonella Serovars. Front. Immunol. 2020, 10, 2966. [Google Scholar] [CrossRef]
- González, C.R.; Isibasi, A.; Ortiz-Navarrete, V.; Paniagua, J.; Kumate, J.; García, J.A.; Blanco, F.; Kumate, J. Lymphocytic Proliferative Response to Outer-Membrane Proteins Isolated from Salmonella. Microbiol. Immunol. 1993, 37, 793–799. [Google Scholar] [CrossRef]
- Salazar-González, R.M.; Maldonado-Bernal, C.; Ramírez-Cruz, N.E.; Rios-Sarabia, N.; Beltrán-Nava, J.; Castañón-González, J.; Castillo-Torres, N.; Palma-Aguirre, J.A.; López-Macías, C.; Isibasi, A. Induction of cellular immune response and anti-Salmonella enterica serovar typhi bactericidal antibodies in healthy volunteers by immunization with a vaccine candidate against typhoid fever. Immunol. Lett. 2004, 93, 115–122. [Google Scholar] [CrossRef]
- Martin, M.D.; Badovinac, V.P. Defining memory CD8 T cell. Front. Immunol. 2018, 9, 2692. [Google Scholar] [CrossRef] [Green Version]
- Antonio-Herrera, L.; Badillo-Godinez, O.; Medina-Contreras, O.; Tepale-Segura, A.; García-Lozano, A.; Gutierrez-Xicotencatl, L.; Soldevila, G.; Esquivel-Guadarrama, F.R.; Idoyaga, J.; Bonifaz, L.C. The Nontoxic Cholera B Subunit Is a Potent Adjuvant for Intradermal DC-Targeted Vaccination. Front. Immunol. 2018, 9, 2212. [Google Scholar] [CrossRef]
- Sckisel, G.D.; Mirsoian, A.; Minnar, C.M.; Crittenden, M.; Curti, B.; Chen, J.Q.; Blazar, B.R.; Borowsky, A.D.; Monjazeb, A.M.; Murphy, W.J. Differential phenotypes of memory CD4 and CD8 T cells in the spleen and peripheral tissues following immunostimulatory therapy. J. Immunother. Cancer 2017, 5, 33. [Google Scholar] [CrossRef]
- Gray, J.I.; Westerhof, L.M.; MacLeod, M.K.L. The roles of resident, central and effector memory CD4 T-cells in protective immunity following infection or vaccination. Immunology 2018, 154, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Steinbach, K.; Vincenti, I.; Merkler, D. Resident-Memory T Cells in tissue-restricted immune responses: For better or worse? Front. Immunol. 2018, 9, 2827. [Google Scholar] [CrossRef] [Green Version]
- Szabo, P.A.; Miron, M.; Farber, D.L. Location, location, location: Tissue resident memory T cells in mice and humans. Sci. Immunol. 2019, 4, 9673. [Google Scholar] [CrossRef]
- Han, S.J.; Glatman Zaretsky, A.; Andrade-Oliveira, V.; Collins, N.; Dzutsev, A.; Shaik, J.; Morais da Fonseca, D.; Harrison, O.J.; Tamoutounour, S.; Byrd, A.L.; et al. White Adipose Tissue Is a Reservoir for Memory T Cells and Promotes Protective Memory Responses to Infection. Immunity 2017, 47, 1154–1168.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijaro, J.R.; Turner, D.; Pham, Q.; Wherry, E.J.; Lefrançois, L.; Farber, D.L. Cutting Edge: Tissue-Retentive Lung Memory CD4 T Cells Mediate Optimal Protection to Respiratory Virus Infection. J. Immunol. 2011, 187, 5510–5514. [Google Scholar] [CrossRef] [Green Version]
- Benoun, J.M.; Peres, N.G.; Wang, N.; Pham, O.H.; Rudisill, V.L.; Fogassy, Z.N.; Whitney, P.G.; Fernandez-Ruiz, D.; Gebhardt, T.; Pham, Q.; et al. Optimal protection against Salmonella infection requires noncirculating memory. Proc. Natl. Acad. Sci. USA 2018, 115, 10416–10421. [Google Scholar] [CrossRef] [Green Version]
- Burkholder, T.; Foltz, C.; Karlsson, E.; Linton, C.G.; Smith, J.M. Health Evaluation of Experimental Laboratory Mice. Curr. Protoc. Mouse Biol. 2012, 2, 145–165. [Google Scholar] [CrossRef] [Green Version]
- Alurkar, V.; Kamat, R. Immunomodulatory properties of porins of some members of the family Enterobacteriaceae. Infect. Immun. 1997, 65, 2382–2388. [Google Scholar] [CrossRef] [Green Version]
- Belson, A.; Schmidt, T.; Fernando, D.; Hardes, K.; Scott, N.; Brett, S.; Clark, D.; Oliveira, J.J.; Davies, B.; McHugh, S.; et al. Characterisation of the clinical and activated T cell response to repeat delayed-type hypersensitivity skin challenges in human subjects, with KLH and PPD, as a potential model to test T cell-targeted therapies. Inflamm. Res. 2016, 65, 389–404. [Google Scholar] [CrossRef]
- Ferraro, A.; Buonocore, S.M.; Auquier, P.; Nicolas, I.; Wallemacq, H.; Boutriau, D.; Van der Most, R. Role and plasticity of Th1 and Th17 responses in immunity to Staphylococcus aureus. Hum. Vaccines Immunother. 2019, 15, 2980–2992. [Google Scholar] [CrossRef] [Green Version]
- Amezcua Vesely, M.C.; Pallis, P.; Bielecki, P.; Esplugues, E.; Gagliani, N.; Flavell, R.A. Effector TH17 Cells Give Rise to Long-Lived TRM Cells that Are Essential for an Immediate Response against Bacterial Infection. Cell 2019, 178. [Google Scholar] [CrossRef] [PubMed]
- Li Causi, E.; Parikh, S.C.; Chudley, L.; Layfield, D.M.; Ottensmeier, C.H.; Stevenson, F.K.; Di Genova, G. Vaccination Expands Antigen-Specific CD4+ Memory T Cells and Mobilizes Bystander Central Memory T Cells. PLoS ONE 2015, 10, e0136717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Letelier, R.A.; Castro-Medina, D.I.; Badillo-Godinez, O.; Tepale-Segura, A.; Huanosta-Murillo, E.; Aguilar-Flores, C.; De León-Rodríguez, S.G.; Mantilla, A.; Fuentes-Pananá, E.M.; López-Macías, C.; et al. Induction of Progenitor Exhausted Tissue-Resident Memory CD8+ T Cells Upon Salmonella Typhi Porins Adjuvant Immunization Correlates With Melanoma Control and Anti-PD-1 Immunotherapy Cooperation. Front. Immunol. 2020, 11, 2893. [Google Scholar] [CrossRef]
- Moreno-Eutimio, M.A.; Tenorio-Calvo, A.; Pastelin-Palacios, R.; Perez-Shibayama, C.; Gil-Cruz, C.; López-Santiago, R.; Baeza, I.; Fernández-Mora, M.; Bonifaz, L.; Isibasi, A.; et al. Salmonella Typhi OmpS1 and OmpS2 porins are potent protective immunogens with adjuvant properties. Immunology 2013, 139, 459–471. [Google Scholar] [CrossRef]
- García-Lozano, A.; Toriello, C.; Antonio-Herrera, L.; Bonifaz, L.C. Sporothrix schenckii Immunization, but Not Infection, Induces Protective Th17 Responses Mediated by Circulating Memory CD4+ T Cells. Front. Microbiol. 2018, 9, 1275. [Google Scholar] [CrossRef] [PubMed]
- Labuda, J.C.; McSorley, S.J. Circulating memory is sufficient for protective immunity to infection with Chlamydia muridarum. J. Immunol. 2020, 204, 82.19. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ontiveros-Padilla, L.; García-Lozano, A.; Tepale-Segura, A.; Rivera-Hernández, T.; Pastelin-Palacios, R.; Isibasi, A.; Arriaga-Pizano, L.A.; Bonifaz, L.C.; López-Macías, C. CD4+ and CD8+ Circulating Memory T Cells Are Crucial in the Protection Induced by Vaccination with Salmonella Typhi Porins. Microorganisms 2021, 9, 770. https://doi.org/10.3390/microorganisms9040770
Ontiveros-Padilla L, García-Lozano A, Tepale-Segura A, Rivera-Hernández T, Pastelin-Palacios R, Isibasi A, Arriaga-Pizano LA, Bonifaz LC, López-Macías C. CD4+ and CD8+ Circulating Memory T Cells Are Crucial in the Protection Induced by Vaccination with Salmonella Typhi Porins. Microorganisms. 2021; 9(4):770. https://doi.org/10.3390/microorganisms9040770
Chicago/Turabian StyleOntiveros-Padilla, Luis, Alberto García-Lozano, Araceli Tepale-Segura, Tania Rivera-Hernández, Rodolfo Pastelin-Palacios, Armando Isibasi, Lourdes A. Arriaga-Pizano, Laura C. Bonifaz, and Constantino López-Macías. 2021. "CD4+ and CD8+ Circulating Memory T Cells Are Crucial in the Protection Induced by Vaccination with Salmonella Typhi Porins" Microorganisms 9, no. 4: 770. https://doi.org/10.3390/microorganisms9040770
APA StyleOntiveros-Padilla, L., García-Lozano, A., Tepale-Segura, A., Rivera-Hernández, T., Pastelin-Palacios, R., Isibasi, A., Arriaga-Pizano, L. A., Bonifaz, L. C., & López-Macías, C. (2021). CD4+ and CD8+ Circulating Memory T Cells Are Crucial in the Protection Induced by Vaccination with Salmonella Typhi Porins. Microorganisms, 9(4), 770. https://doi.org/10.3390/microorganisms9040770