
Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolate Collection
2.2. Determination of Fungicide Sensitivity
2.3. Identifying Target Site Mutations in Fungicide Target Proteins
2.4. Testing for Potential Overexpression of the CYP51 and MFS1 Genes
2.5. Statistical Analysis
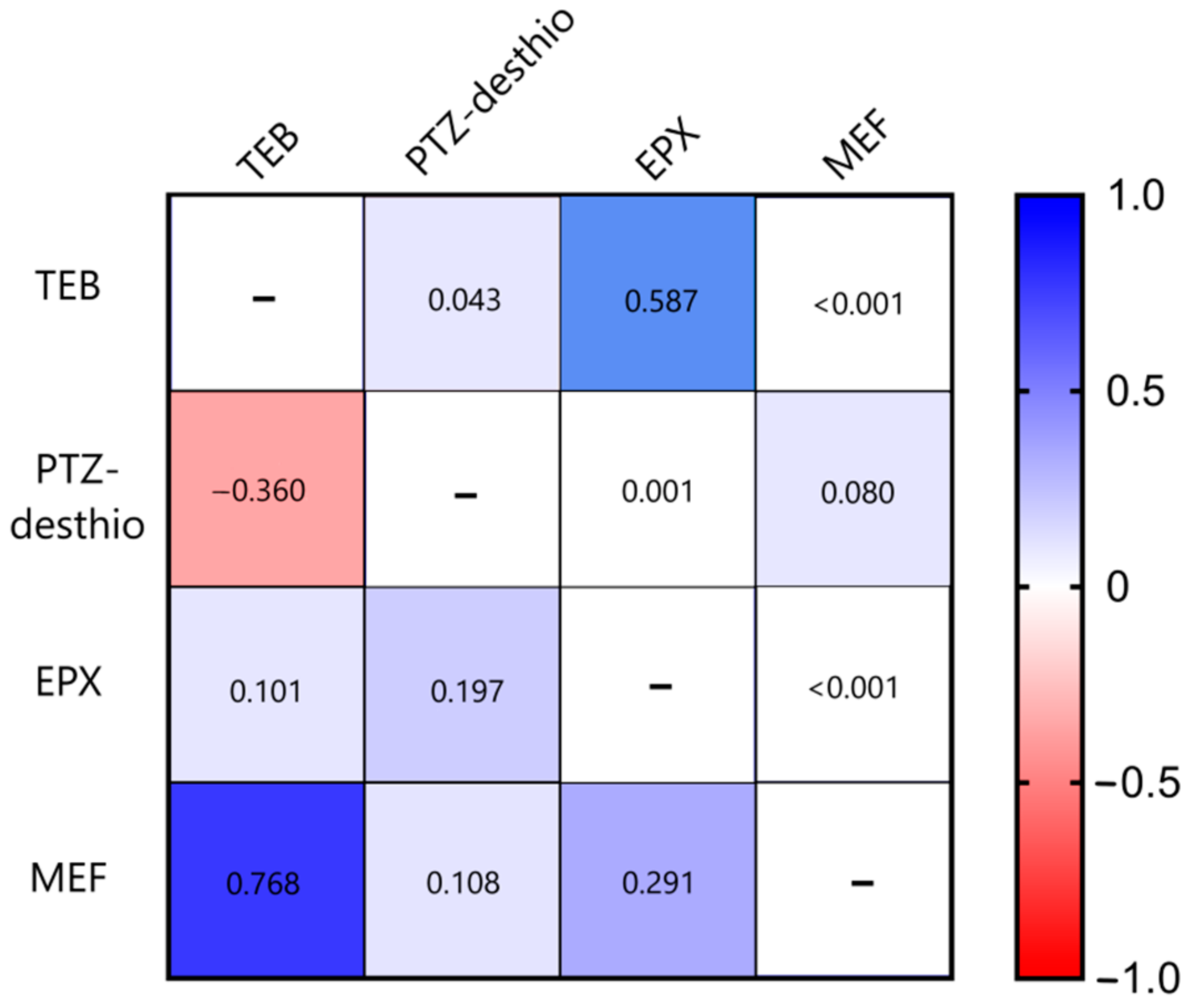
3. Results
3.1. Status of DMI Fungicide Sensitivity
3.2. Mutations in CYP51 Gene
3.3. Analysis of CYP51 and MFS1 Promoter Region
3.4. Status of SDHI Resistance and Mutations in Sdh Protein Subunits
3.5. Mutation G143A Prevalence in CytB Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, J.K.M.; Chartrain, L.; Lasserre-Zuber, P.; Saintenac, C. Genetics of resistance to Zymoseptoria tritici and applications to wheat breeding. Fungal Genet. Biol. 2015, 79, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; McDonald, M.C.; Solomon, P.S.; Milgate, A.W. Genetic Mapping of Stb19, a New Resistance Gene to Zymoseptoria Tritici in Wheat. Theor. Appl. Genet. 2018, 131, 2765–2773. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.N.; Matzen, N.; Hansen, J.G.; Semaskiene, R.; Korbas, M.; Danielewicz, J.; Glazek, M.; Maumene, C.; Rodemann, B.; Weigand, S.; et al. Four Azoles’ Profile in the Control of Septoria, Yellow Rust and Brown Rust in Wheat Across Europe. Crop Prot. 2018, 105, 16–27. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Matzen, N.; Heick, T.M.; Havis, N.; Holdgate, S.; Clark, B.; Blake, J.; Glazek, M.; Korbas, M.; Danielewicz, J.; et al. Decreasing Azole Sensitivity of Z. Tritici in Europe Contributes to Reduced and Varying Field Efficacy. J. Plant Dis. Prot. 2020. [Google Scholar] [CrossRef]
- FRAC Fungicide Resistance Action Committee. Available online: https://www.frac.info/ (accessed on 10 January 2021).
- Zhan, J.; McDonald, B.A. The Interaction Among Evolutionary Forces in the Pathogenic Fungus Mycosphaerella Graminicola. Fungal Genet. Biol. 2004, 41, 590–599. [Google Scholar] [CrossRef]
- Cools, H.J.; Fraaije, B.A. Update on Mechanisms of Azole Resistance in Mycosphaerella Graminicola and Implications for Future Control. Pest Manag. Sci. 2013, 69, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Dooley, H.; Shaw, M.W.; Mehenni-Ciz, J.; Spink, J.; Kildea, S. Detection of Zymoseptoria Tritici SDHI-insensitive Field Isolates Carrying the SdhC-H152R and SdhD-R47W Substitutions. Pest Manag. Sci. 2016, 72, 2203–2207. [Google Scholar] [CrossRef]
- Wieczorek, T.M.; Berg, G.; Semaškienė, R.; Mehl, A.; Sierotzki, H.; Stammler, G.; Justesen, A.F.; Jørgensen, L.N. Impact of DMI and SDHI Fungicides on Disease Control and CYP51 Mutations in Populations of Zymoseptoria Tritici from Northern Europe. Eur. J. Plant Pathol. 2015, 143, 861–871. [Google Scholar] [CrossRef]
- Dubos, T.; Pasquali, M.; Pogoda, F.; Casanova, A.; Hoffmann, L.; Beyer, M. Differences Between the Succinate Dehydrogenase Sequences of Isopyrazam Sensitive Zymoseptoria Tritici and Insensitive Fusarium Graminearum Strains. Pestic. Biochem. Physiol. 2013, 105, 28–35. [Google Scholar] [CrossRef]
- Sierotzki, H.; Scalliet, G. A Review of Current Knowledge of Resistance Aspects for the Next-Generation Succinate Dehydrogenase Inhibitor Fungicides. Phytopathology 2013, 103, 880–887. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Sanglard, D.; Calabrese, D.; Sirjusingh, C.; Anderson, J.B.; Kohn, L.M. Evolution of Drug Resistance in Experimental Populations of Candida Albicans. J. Bacteriol. 2000, 182, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Grimmer, M.K.; van den Bosch, F.; Powers, S.J.; Paveley, N.D. Fungicide Resistance Risk Assessment Based on Traits Associated with the Rate of Pathogen Evolution. Pest Manag. Sci. 2015, 71, 207–215. [Google Scholar] [CrossRef]
- Russell, P.E. A Century of Fungicide Evolution. J. Agric. Sci. 2005, 143, 11–25. [Google Scholar] [CrossRef]
- Cools, H.J.; Mullins, J.G.L.; Fraaije, B.A.; Parker, J.E.; Kelly, D.E.; Lucas, J.A.; Kelly, S.L. Impact of Recently Emerged Sterol 14α-demethylase (CYP51) Variants of Mycosphaerella Graminicola on Azole Fungicide Sensitivity. Appl. Environ. Microbiol. 2011, 77, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Cools, H.J.; Bayon, C.; Atkins, S.; Lucas, J.A.; Fraaije, B.A. Overexpression of the Sterol 14α-demethylase gene (MgCYP51) in Mycosphaerella Graminicola Isolates Confers a Novel Azole Fungicide Sensitivity Phenotype. Pest Manag. Sci. 2012, 68, 1034–1040. [Google Scholar] [CrossRef]
- Leroux, P.; Walker, A.S. Multiple Mechanisms Account for Resistance to Sterol 14α-demethylation Inhibitors in Field Isolates of Mycosphaerella Graminicola. Pest Manag. Sci. 2011, 67, 44–59. [Google Scholar] [CrossRef]
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a New Nomenclature for CYP51 Haplotypes in Zymoseptoria Tritici and Analysis of their Distribution in Europe. Plant Pathol. 2018, 67, 1706–1712. [Google Scholar] [CrossRef]
- Leroux, P.; Albertini, C.; Gautier, A.; Gredt, M.; Walker, A. Mutations in the CYP51 Gene Correlated with Changes in Sensitivity to Sterol 14α-demethylation Inhibitors in Field Isolates of Mycosphaerella Graminicola. Pest Manag. Sci. 2007, 63, 688–698. [Google Scholar] [CrossRef]
- Garnault, M.; Duplaix, C.; Leroux, P.; Couleaud, G.; Carpentier, F.; David, O.; Walker, A.S. Spatiotemporal Dynamics of Fungicide Resistance in the Wheat Pathogen Zymoseptoria Tritici in France. Pest Manag. Sci. 2019, 75, 1794–1807. [Google Scholar] [CrossRef]
- Heick, T.M.; Matzen, N.; Jørgensen, L.N. Reduced Field Efficacy and Sensitivity of Demethylation Inhibitors in the Danish and Swedish Zymoseptoria Tritici Populations. Eur. J. Plant Pathol. 2020, 157, 625–636. [Google Scholar] [CrossRef]
- Kildea, S.; Bucar, D.E.; Hutton, F.; de la Rosa, S.; Welch, T.E.; Phelan, S. Prevalence of QoI Resistance and mtDNA Diversity in the Irish Zymoseptoria Tritici Population. Irish J. Agric. Food Res. 2019, 58, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Mäe, A.; Fillinger, S.; Sooväli, P.; Heick, T.M. Fungicide Sensitivity Shifting of Zymoseptoria Tritici in the Finnish-Baltic Region and a Novel Insertion in the MFS1 Promoter. Front. Plant Sci. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rehfus, A.; Miessner, S.; Achenbach, J.; Strobel, D.; Bryson, R.; Stammler, G. Emergence of Succinate Dehydrogenase Inhibitor Resistance of Pyrenophora Teres in Europe. Pest Manag. Sci. 2016, 72, 1977–1988. [Google Scholar] [CrossRef]
- Stammler, G.; Wolf, A.; Glaettli, A.; Klappach, K. Respiration Inhibitors: Complex II. In Fungicide Resistance in Plant Pathogens; Ishii, H., Hollomon, D.W., Eds.; Springer: Tokyo, Japan, 2015; pp. 105–117. [Google Scholar]
- Succinate Dehydrogenase Inhibitor (SDHI) Working Group of the Fungicide Resistance Action Committee (FRAC). Available online: https://www.frac.info/docs/default-source/working-groups/sdhi-fungicides/sdhi-meeting-minutes/minutes-of-the-2020-sdhi-meeting-jan-june-september-2020-with-recommendations-for-2020.pdf?sfvrsn=d7b8499a_2 (accessed on 23 February 2021).
- Hellin, P.; Duvivier, M.; Clinckemaillie, A.; Bataille, C.; Legrève, A.; Heick, T.M.; Jørgensen, L.N.; Andersson, B.; Samils, B.; Rodemann, B.; et al. Multiplex qPCR Assay for Simultaneous Quantification of CYP51-S524T and SdhC-H152R Substitutions in European Populations of Zymoseptoria Tritici. Plant Pathol. 2020, 69, 1666–1677. [Google Scholar] [CrossRef]
- Fraaije, B.A.; Bayon, C.; Atkins, S.; Cools, H.J.; Lucas, J.A.; Fraaije, M.W. Risk Assessment Studies on Succinate Dehydrogenase Inhibitors, the New Weapons in the Battle to Control Septoria Leaf Blotch in Wheat. Mol. Plant Pathol. 2012, 13, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Scalliet, G.; Bowler, J.; Luksch, T.; Kirchhofer-Allan, L.; Steinhauer, D.; Ward, K.; Niklaus, M.; Verras, A.; Csukai, M.; Daina, A.; et al. Mutagenesis and Functional Studies with Succinate Dehydrogenase Inhibitors in the Wheat Pathogen Mycosphaerella Graminicola. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms Influencing the Evolution of Resistance to Qo Inhibitor Fungicides. Pest Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef]
- Fraaije, B.A.; Cools, H.J.; Fountaine, J.; Lovell, D.J.; Motteram, J.; West, J.S.; Lucas, J.A. Role of Ascospores in Further Spread of QoI-Resistant Cytochrome B Alleles (G143A) in Field Populations of Mycosphaerella Graminicola. Phytopathology 2005, 95, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Torriani, S.F.F.; Brunner, P.C.; McDonald, B.A.; Sierotzki, H. QoI Resistance Emerged Independently at least 4 Times in European Populations of Mycosphaerella Graminicola. Pest Manag. Sci. 2009, 65, 155–162. [Google Scholar] [CrossRef]
- Heick, T.M.; Justesen, A.F.; Jørgensen, L.N. Resistance of Wheat Pathogen Zymoseptoria Tritici to DMI and QoI Fungicides in the Nordic-Baltic Region—A Status. Eur. J. Plant Pathol. 2017, 149, 669–682. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, S.; Liu, X.Z.; Wen, H.A.; Wang, M. A Simple Method of Genomic DNA Extraction Suitable for Analysis of Bulk Fungal Strains. Lett. Appl. Microbiol. 2010, 51, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Kildea, S.; Mehenni-Ciz, J.; Spink, J.; O’Sullivan, E. Changes in the frequency of Irish Mycosphaerella graminicola CYP51 variants 2006–2011. In Proceedings of the Modern Fungicides and Antifungal Compounds, Friedrichroda, Germany, 21–25 April 2014; Volume VII, pp. 143–144. [Google Scholar]
- Rehfus, A.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in sdh Genes in Field Isolates of Zymoseptoria Tritici and Impact on the Sensitivity to Various Succinate Dehydrogenase Inhibitors. Plant Pathol. 2018, 67, 175–180. [Google Scholar] [CrossRef]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.S.; Fillinger, S. Fungicide Efflux and the MgMFS1 Transporter Contribute to the Multidrug Resistance Phenotype in Zymoseptoria Tritici Field Isolates. Environ. Microbiol. 2015, 17, 2805–2823. [Google Scholar] [CrossRef] [PubMed]
- Goedhart, J. SuperPlotsOfData—A Web App for the Transparent Display and Quantitative Comparison of Continuous Data from Discrete Conditions. bioRxiv 2020. [Google Scholar] [CrossRef]
- Blake, J.J.; Gosling, P.; Fraaije, B.A.; Burnett, F.J.; Knight, S.M.; Kildea, S.; Paveley, N.D. Changes in Field Dose–Response Curves for Demethylation Inhibitor (DMI) and Quinone Outside Inhibitor (QoI) Fungicides Against Zymoseptoria Tritici, Related to Laboratory Sensitivity Phenotyping and Genotyping Assays. Pest Manag. Sci. 2018, 74, 302–313. [Google Scholar] [CrossRef]
- Kildea, S.; Marten-Heick, T.; Grant, J.; Mehenni-Ciz, J.; Dooley, H. A Combination of Target-Site Alterations, Overexpression and Enhanced Efflux Activity Contribute to Reduced Azole Sensitivity Present in the Irish Zymoseptoria Tritici Population. Eur. J. Plant Pathol. 2019, 154, 529–540. [Google Scholar] [CrossRef]
- Dooley, H.; Shaw, M.W.; Spink, J.; Kildea, S. The Effect of Succinate Dehydrogenase Inhibitor/Azole Mixtures on Selection of Zymoseptoria Tritici Isolates with Reduced Sensitivity. Pest Manag. Sci. 2016, 72, 1150–1159. [Google Scholar] [CrossRef]
- Yamashita, M.; Fraaije, B. Non-target site SDHI Resistance is Present as Standing Genetic Variation in Field Populations of Zymoseptoria Tritici. Pest Manag. Sci. 2018, 74, 672–681. [Google Scholar] [CrossRef] [Green Version]
- Cheval, P.; Siah, A.; Bomble, M.; Popper, A.D.; Reignault, P.; Halama, P. Evolution of QoI Resistance of the Wheat Pathogen Zymoseptoria Tritici in Northern France. Crop Prot. 2017, 92, 131–133. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | County * | No of Fields | No of Isolates |
|---|---|---|---|
| 2019 | JÕ | 3 | 19 |
| LV | 8 | 49 | |
| VA | 1 | 6 | |
| VI | 5 | 19 | |
| VÕ | 2 | 11 | |
| 2020 | IV | 4 | 19 |
| JÕ | 5 | 24 | |
| JÄ | 4 | 20 | |
| LV | 4 | 21 | |
| PÕ | 3 | 17 | |
| TA | 4 | 21 | |
| VA | 3 | 15 | |
| VI | 5 | 21 | |
| VÕ | 4 | 21 | |
| Total | 55 | 282 |
| Commercial Product | Field Treatment (L ha−1) | Active Ingredients | Concentration (g ha−1) |
|---|---|---|---|
| Viverda (BASF) | 0.8 | Epoxiconazole * | 40 |
| Boscalid * | 112 | ||
| Pyraclostrobin | 48 | ||
| Input (Bayer) | 0.4 | Prothioconazole * | 64 |
| Spiroxamine | 120 | ||
| Curbatur (Bayer) | 0.4 | Prothioconazole * | 100 |
| Tango Super (BASF) | 0.6 | Epoxiconazole * | 50.4 |
| Fenpropimorf | 150 | ||
| Priaxor (BASF) | 0.4 | Fluxapyroxad * | 30 |
| Pyraclostrobin | 60 |
| County * | Year | EPX | PTZ-desthio | MEF | |||
|---|---|---|---|---|---|---|---|
| EC50 | RF | EC50 | RF | EC50 | RF | ||
| IV | 2019 | NA ** | NA | NA | NA | NA | NA |
| 2020 | 0.3 | 12 | 0.5 | 35 | 0.17 | 163 | |
| JÕ | 2019 | 2.79 | 109 | 0.1 | 10 | 0.41 | 412 |
| 2020 | 0.66 | 26 | 0.23 | 16 | 0.22 | 218 | |
| JÄ | 2019 | NA | NA | NA | NA | NA | NA |
| 2020 | 0.68 | 27 | 0.2 | 14 | 0.06 | 58 | |
| LV | 2019 | 2.72 | 105 | 0.13 | 13 | 0.3 | 300 |
| 2020 | 0.72 | 28 | 0.23 | 16 | 0.13 | 124 | |
| PÕ | 2019 | NA | NA | NA | NA | NA | NA |
| 2020 | 0.95 | 37 | 0.42 | 29 | 0.09 | 93 | |
| TA | 2019 | NA | NA | NA | NA | NA | NA |
| 2020 | 0.43 | 17 | 0.07 | 5 | 0.07 | 73 | |
| VA | 2019 | 1.37 | 53 | NA | NA | 0.01 | 5 |
| 2020 | 0.48 | 19 | 0.08 | 6 | 0.06 | 63 | |
| VI | 2019 | 2.98 | 114 | 0.07 | 7 | 0.1 | 95 |
| 2020 | 0.5 | 19 | 0.08 | 6 | 0.11 | 105 | |
| VÕ | 2019 | 3.34 | 131 | 0.16 | 16 | 0.1 | 98 |
| 2020 | 0.63 | 25 | 0.11 | 8 | 0.16 | 157 | |
| Ref. IPO323 | 0.026 | NA | 0.014 | NA | 0.001 | NA | |
| County * | CYP51 Mutation Frequency (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D134G | V136A | V136C | A379G | I381V | S524T | |||||||
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| IV | NA ** | 5 | NA | 5 | NA | 0 | NA | 42 | NA | 100 | NA | 0 |
| JÕ | 0 | 38 | 0 | 38 | 0 | 25 | 31 | 8 | 100 | 92 | 0 | 33 |
| JÄ | NA | 60 | NA | 55 | NA | 0 | NA | 10 | NA | 90 | NA | 40 |
| LV | 12 | 33 | 33 | 24 | 22 | 19 | 33 | 33 | 100 | 95 | 20 | 24 |
| PÕ | NA | 29 | NA | 35 | NA | 0 | NA | 24 | NA | 100 | NA | 35 |
| TA | NA | 62 | NA | 19 | NA | 0 | NA | 10 | NA | 38 | NA | 14 |
| VA | 67 | 40 | 67 | 33 | 0 | 0 | 6 | 0 | 100 | 100 | 0 | 33 |
| VI | 46 | 62 | 46 | 48 | 12 | 14 | 23 | 48 | 100 | 86 | 12 | 19 |
| VÕ | 60 | 42 | 80 | 5 | 0 | 16 | 0 | 16 | 100 | 95 | 20 | 16 |
| Average | 37 | 42 | 45 | 30 | 7 | 10 | 19 | 21 | 100 | 88 | 10 | 24 |
| County * | Year | BOS | FLX | ||
|---|---|---|---|---|---|
| EC50 | RF | EC50 | RF | ||
| IV | 2019 | NA ** | NA | NA | NA |
| 2020 | 0.34 | 2 | 0.1 | 1 | |
| JÕ | 2019 | 0.52 | 4 | 0.09 | 1 |
| 2020 | 0.48 | 3 | 0.16 | 2 | |
| JÄ | 2019 | NA | NA | NA | NA |
| 2020 | 0.84 | 6 | 0.23 | 3 | |
| LV | 2019 | 0.74 | 5 | 0.08 | 1 |
| 2020 | 0.47 | 3 | 0.09 | 1 | |
| PÕ | 2019 | NA | NA | NA | NA |
| 2020 | 0.35 | 2 | 0.08 | 1 | |
| TA | 2019 | NA | NA | NA | NA |
| 2020 | 0.7 | 5 | 0.14 | 2 | |
| VA | 2019 | 0.2 | 1 | NA | NA |
| 2020 | 0.78 | 5 | 0.15 | 2 | |
| VI | 2019 | 0.58 | 4 | 0.08 | 1 |
| 2020 | 0.46 | 3 | 0.1 | 1 | |
| VÕ | 2019 | 1.3 | 9 | 0.25 | 3 |
| 2020 | 0.75 | 5 | 0.14 | 2 | |
| Ref. IPO323 | 0.144 | NA | 0.083 | NA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiiker, R.; Juurik, M.; Heick, T.M.; Mäe, A. Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020. Microorganisms 2021, 9, 814. https://doi.org/10.3390/microorganisms9040814
Kiiker R, Juurik M, Heick TM, Mäe A. Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020. Microorganisms. 2021; 9(4):814. https://doi.org/10.3390/microorganisms9040814
Chicago/Turabian StyleKiiker, Riinu, Marite Juurik, Thies Marten Heick, and Andres Mäe. 2021. "Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020" Microorganisms 9, no. 4: 814. https://doi.org/10.3390/microorganisms9040814