
Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Organic Acids and Nature-Identical Compounds—Chemicals and Test Solutions
2.3. Minimal Inhibitory Concentration (MIC) Assay
2.4. Minimal Bactericidal Concentration (MBC) Assay
2.5. Statistical Analysis
3. Results
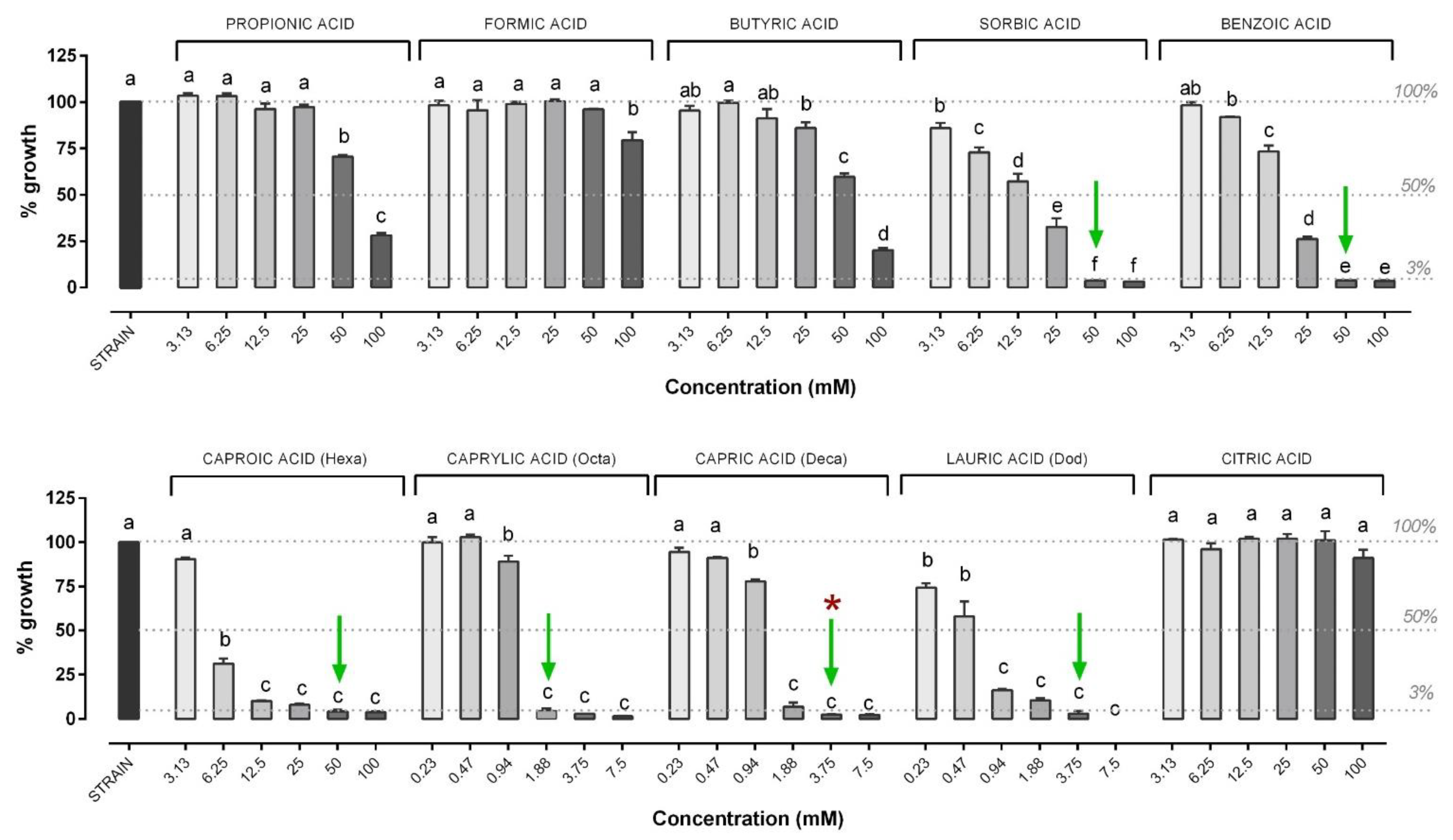
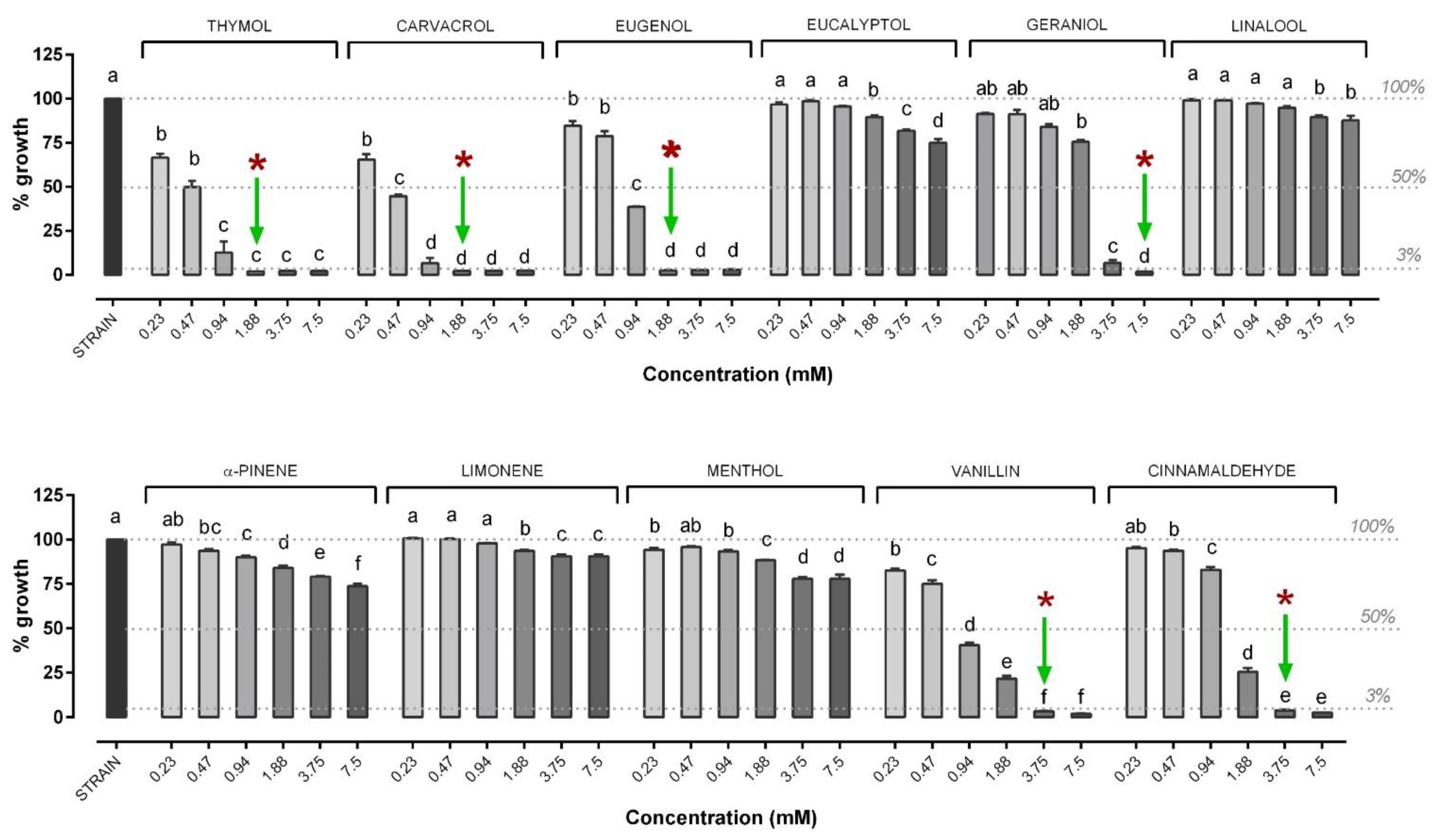
3.1. Minimal Inhibitory Concentration and Minimal Bactericidal Concentration of OA and NIC against V. anguillarum
3.2. Minimal Inhibitory Concentration and Minimal Bactericidal Concentration of OA and NIC against V. harveyi
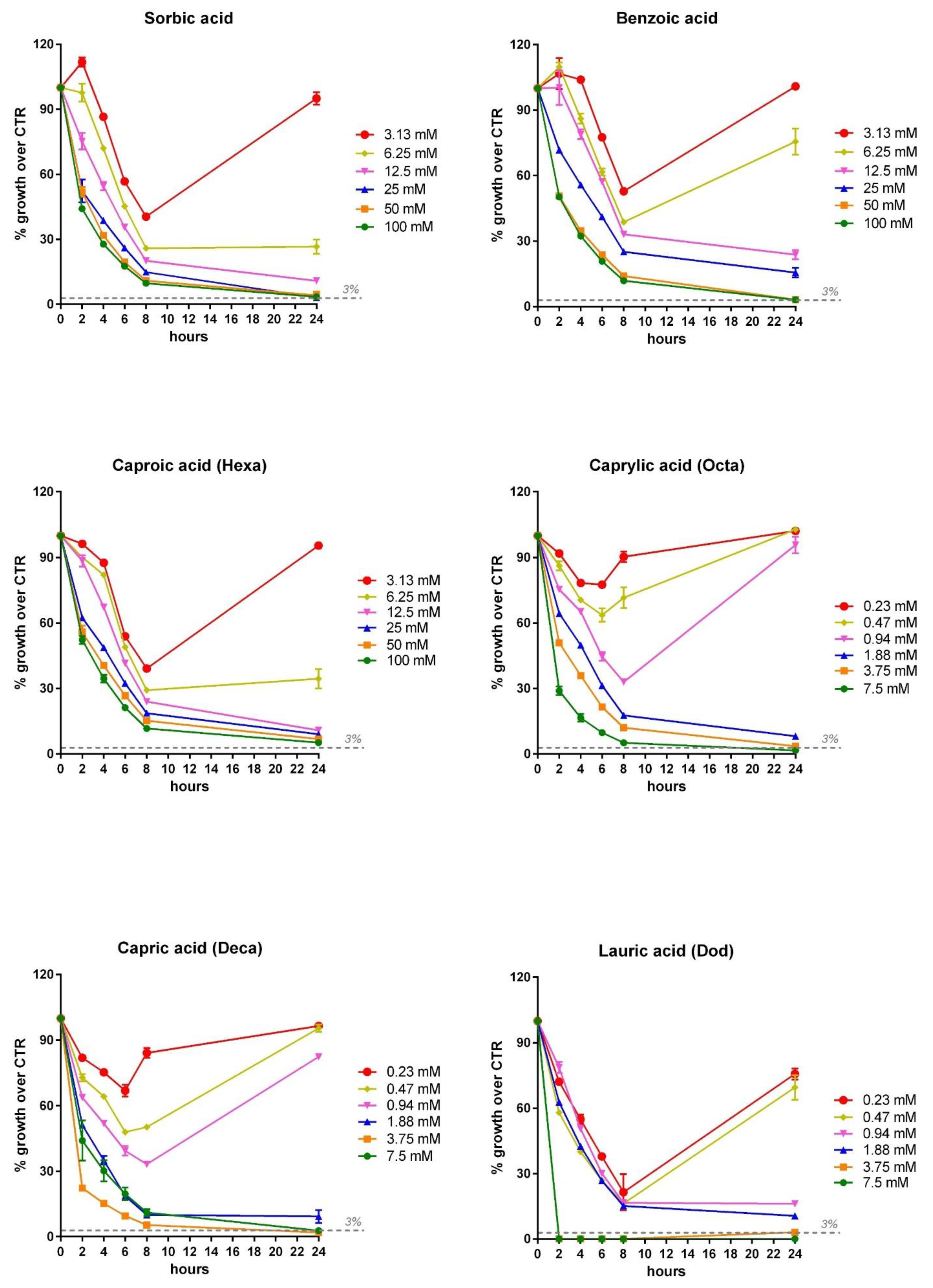
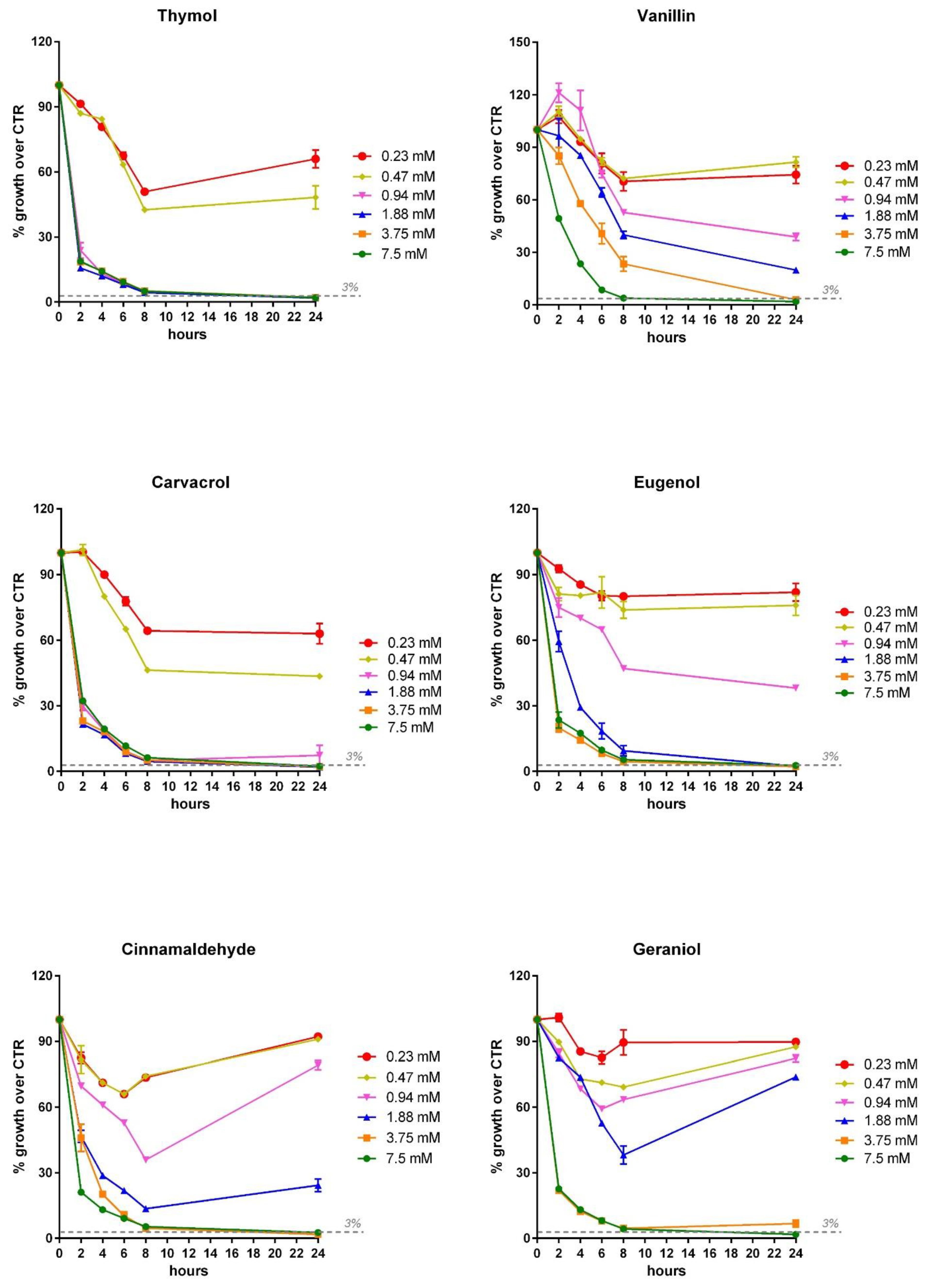
3.3. Inhibition Dose Response Curve of OA and NIC against V. harveyi and V. anguillarum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Novriadi, R. Vibriosis in Aquaculture. Omni Akuatika 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Haldar, S. Vibrio Related Diseases in Aquaculture and Development of Rapid and Accurate Identification Methods. J. Mar. Sci. Res. Dev. S 2012, 1, 1–7. [Google Scholar] [CrossRef]
- Scarano, C.; Spanu, C.; Ziino, G.; Pedonese, F.; Dalmasso, A.; Spanu, V.; Virdis, S.; De Santis, E.P.L. Antibiotic Resistance of Vibrio Species Isolated from Sparus Aurata Reared in Italian Mariculture. New Microbiol. 2014, 37, 329–337. [Google Scholar] [PubMed]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.; Ramos, F. Antimicrobial Resistance in Aquaculture: Current Knowledge and Alternatives to Tackle the Problem. Int. J. Antimicrob. Agents 2018, 52, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Roque, A.; Molina-Aja, A.; Bolán-Mejía, C.; Gomez-Gil, B. In Vitro Susceptibility to 15 Antibiotics of Vibrios Isolated from Penaeid Shrimps in Northwestern Mexico. Int. J. Antimicrob. Agents 2001, 17, 383–387. [Google Scholar] [CrossRef]
- Albuquerque Costa, R.; Araújo, R.L.; Souza, O.V.; Vieira, R.H.S.D.F. Antibiotic-Resistant Vibrios in Farmed Shrimp. BioMed Res. Int. 2015, 2015, 505914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, H.; Zhang, X.; Song, L.; Chang, Y.; Yang, G. Molecular Characterizations of Oxytetracycline Resistant Bacteria and Their Resistance Genes from Mariculture Waters of China. Mar. Pollut. Bull. 2006, 52, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-H.; Kim, Y.; Oh, S.J.; Mok, J.-S.; Cho, M.-H.; So, J.-S. Antibiotic Resistance of Vibrio Harveyi Isolated from Seawater in Korea. Mar. Pollut. Bull. 2014, 86, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Laganà, P.; Caruso, G.; Minutoli, E.; Zaccone, R.; Santi, D. Susceptibility to Antibiotics of Vibrio Spp. and Photobacterium Damsela Ssp. Piscicida Strains Isolated from Italian Aquaculture Farms. New Microbiol. 2011, 34, 53–63. [Google Scholar] [PubMed]
- Letchumanan, V.; Pusparajah, P.; Tan, L.T.-H.; Yin, W.-F.; Lee, L.-H.; Chan, K.-G. Occurrence and Antibiotic Resistance of Vibrio Parahaemolyticus from Shellfish in Selangor, Malaysia. Front. Microbiol. 2015, 6, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Aja, A.; Garcà a-Gasca, A.; Abreu-Grobois, A.; Bolán-Mejà a, C.; Roque, A.; Gomez-Gil, B. Plasmid Profiling and Antibiotic Resistance of Vibrio Strains Isolated from Cultured Penaeid Shrimp. FEMS Microbiol. Lett. 2002, 213, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raissy, M.; Moumeni, M.; Ansari, M.; Rahimi, E. Antibiotic Resistance Pattern of Some Vibrio Strains Isolated from Seafood. Iran. J. Fish. Sci. 2012, 11, 618–626. [Google Scholar]
- Snoussi, M.; Chaieb, K.; Mahmoud, R.; Bakhrouf, A. Quantitative Study, Identification and Antibiotics Sensitivity of SomeVibrionaceae Associated to a Marine Fish Hatchery. Ann. Microbiol. 2006, 56, 289–293. [Google Scholar] [CrossRef]
- Tan, C.W.; Rukayadi, Y.; Hasan, H.; Thung, T.Y.; Lee, E.; Rollon, W.D.; Hara, H.; Kayali, A.Y.; Nishibuchi, M.; Radu, S. Prevalence and Antibiotic Resistance Patterns of Vibrio Parahaemolyticus Isolated from Different Types of Seafood in Selangor, Malaysia. Saudi J. Biol. Sci. 2020, 27, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Das, R.K.; Kaur Brar, S.; Rouissi, T.; Avalos Ramirez, A.; Chorfi, Y.; Godbout, S. Alternatives to Antibiotics in Poultry Feed: Molecular Perspectives. Crit. Rev. Microbiol. 2018, 44, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Thacker, P.A. Alternatives to Antibiotics as Growth Promoters for Use in Swine Production: A Review. J. Anim. Sci. Biotechnol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single Components of Botanicals and Nature-Identical Compounds as a Non-Antibiotic Strategy to Ameliorate Health Status and Improve Performance in Poultry and Pigs. Nutr. Res. Rev. 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tugnoli, B.; Giovagnoni, G.; Piva, A.; Grilli, E. From Acidifiers to Intestinal Health Enhancers: How Organic Acids Can Improve Growth Efficiency of Pigs. Animals 2020, 10, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; Tugnoli, B.; Rossi, B.; Giovagnoni, G.; Piva, A.; Grilli, E. Nature-Identical Compounds and Organic Acids Reduce E. Coli K88 Growth and Virulence Gene Expression In Vitro. Toxins 2020, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of Minimum Inhibitory Concentrations (MICs) of Antibacterial Agents by Broth Dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar] [CrossRef] [Green Version]
- Giovagnoni, G.; Tugnoli, B.; Piva, A.; Grilli, E. Organic Acids and Nature Identical Compounds Can Increase the Activity of Conventional Antibiotics Against Clostridium Perfringens and Enterococcus Cecorum In Vitro. J. Appl. Poult. Res. 2019, 28, 1398–1407. [Google Scholar] [CrossRef]
- Hickey, M.E.; Lee, J.L. A Comprehensive Review of Vibrio (Listonella) Anguillarum: Ecology, Pathology and Prevention. Rev. Aquac. 2018, 10, 585–610. [Google Scholar] [CrossRef]
- Ng, W.K.; Koh, C.B. The Utilization and Mode of Action of Organic Acids in the Feeds of Cultured Aquatic Animals. Rev. Aquac. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Mine, S.; Boopathy, R. Effect of Organic Acids on Shrimp Pathogen, Vibrio Harveyi. Curr. Microbiol. 2011, 63, 1–7. [Google Scholar] [CrossRef]
- Austin, B.; Zhang, X.-H. Vibrio Harveyi: A Significant Pathogen of Marine Vertebrates and Invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio Anguillarum as a Fish Pathogen: Virulence Factors, Diagnosis and Prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of Plant Extracts in Fish Aquaculture as an Alternative to Chemotherapy: Current Status and Future Perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Kosiyaporn, H.; Chanvatik, S.; Issaramalai, T.; Kaewkhankhaeng, W.; Kulthanmanusorn, A.; Saengruang, N.; Witthayapipopsakul, W.; Viriyathorn, S.; Kirivan, S.; Kunpeuk, W.; et al. Surveys of Knowledge and Awareness of Antibiotic Use and Antimicrobial Resistance in General Population: A Systematic Review. PLoS ONE 2020, 15, e0227973. [Google Scholar] [CrossRef]
- Sutili, F.J.; Gatlin, D.M.; Heinzmann, B.M.; Baldisserotto, B. Plant Essential Oils as Fish Diet Additives: Benefits on Fish Health and Stability in Feed. Rev. Aquac. 2018, 10, 716–726. [Google Scholar] [CrossRef]
- Zhang, D.; Gan, R.-Y.; Zhang, J.-R.; Farha, A.K.; Li, H.-B.; Zhu, F.; Wang, X.-H.; Corke, H. Antivirulence Properties and Related Mechanisms of Spice Essential Oils: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1018–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konuk, H.B.; Ergüden, B. Phenolic –OH Group Is Crucial for the Antifungal Activity of Terpenoids via Disruption of Cell Membrane Integrity. Folia Microbiol. 2020, 65, 775–783. [Google Scholar] [CrossRef]
- Brackman, G.; Defoirdt, T.; Miyamoto, C.; Bossier, P.; Van Calenbergh, S.; Nelis, H.; Coenye, T. Cinnamaldehyde and Cinnamaldehyde Derivatives Reduce Virulence in Vibrio Spp. by Decreasing the DNA-Binding Activity of the Quorum Sensing Response Regulator LuxR. BMC Microbiol. 2008, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Brackman, G.; Celen, S.; Hillaert, U.; Van Calenbergh, S.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Structure-Activity Relationship of Cinnamaldehyde Analogs as Inhibitors of AI-2 Based Quorum Sensing and Their Effect on Virulence of Vibrio Spp. PLoS ONE 2011, 6, e16084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimoldi, S.; Gliozheni, E.; Ascione, C.; Gini, E.; Terova, G. Effect of a Specific Composition of Short- and Medium-Chain Fatty Acid 1-Monoglycerides on Growth Performances and Gut Microbiota of Gilthead Sea Bream (Sparus Aurata). PeerJ 2018, 6, e5355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngurah, B.I.G.M.; Ni Nyoman, Y.; Dafroyati, Y.; Gede Aris Gunadi, I.; Taneo, M. Antibacterial Evaluation of 2,4-Dihidroxy Benzoic Acid on Escherichia Coli and Vibrio Alginolyticus. J. Phys. Conf. Ser. 2020, 1503, 012027. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, B.; Esteban, M.A.; García-Beltran, J.M.; Giovagnoni, G.; Cuesta, A.; Piva, A.; Grilli, E. Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study. Microorganisms 2021, 9, 966. https://doi.org/10.3390/microorganisms9050966
Rossi B, Esteban MA, García-Beltran JM, Giovagnoni G, Cuesta A, Piva A, Grilli E. Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study. Microorganisms. 2021; 9(5):966. https://doi.org/10.3390/microorganisms9050966
Chicago/Turabian StyleRossi, Barbara, Maria Angéles Esteban, José María García-Beltran, Giulia Giovagnoni, Alberto Cuesta, Andrea Piva, and Ester Grilli. 2021. "Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study" Microorganisms 9, no. 5: 966. https://doi.org/10.3390/microorganisms9050966
APA StyleRossi, B., Esteban, M. A., García-Beltran, J. M., Giovagnoni, G., Cuesta, A., Piva, A., & Grilli, E. (2021). Antimicrobial Power of Organic Acids and Nature-Identical Compounds against Two Vibrio spp.: An In Vitro Study. Microorganisms, 9(5), 966. https://doi.org/10.3390/microorganisms9050966