3.1. Overview of Nocturne116 Genome
Intact virions of Nocturne116 demonstrate a siphovirus morphology having 59.1 ± 1.5 nm × 40.1 ± 1.2 nm prolate capsids with 112.0 ± 8.1 nm long and 9.5 ± 0.8 nm wide non-contractile tails (
Supplementary Figure S2), encompassing a 25,554 bp long dsDNA molecule with 10 bp long 3′ cohesive terminal overhangs (5’-CGCAGTAACT-3’), a GC content of 37.99% and a predicted coding capacity of 94.71% based on the length of its 52 open reading frames (counting overlaps of ORFs once), which corresponds to the complete genome of
Lactococcus phage Nocturne116 therein.
BLASTN search querying the complete nucleotide sequence of Nocturne116 revealed striking dissimilarity to other phage genomes, save for an alignment to the genome of
Lactococcus phage Q54 (DQ490056.1; Query coverage of 36%, 68.08% percent identity,
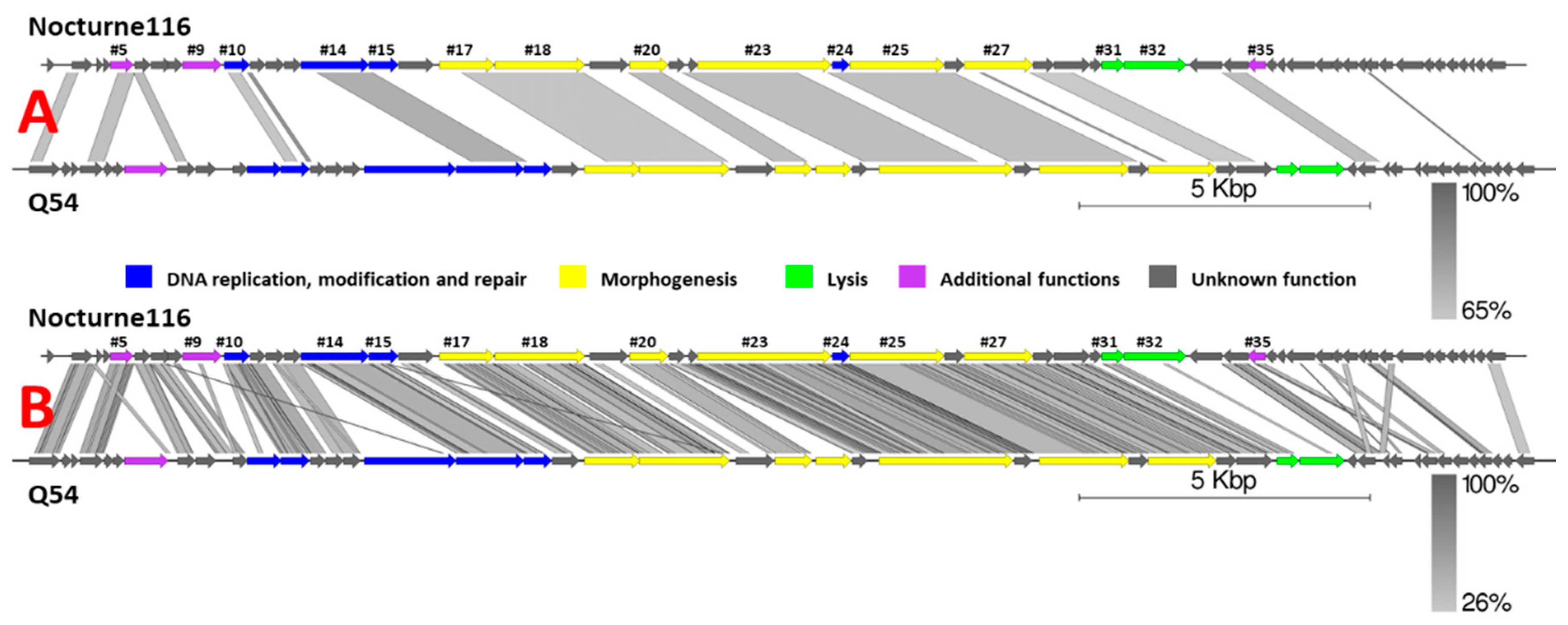
Figure 1), which, as of now, is also the best scoring BLASTN hit and shows ~24.5% similarity (query coverage x identity) to Nocturne116. Sequences from the genomes of different
Lactococcus species and other
Lactococcus phage genomes have shown considerably weaker alignments of up to 5% query coverage and up to 76.31% identity to Nocturne116, corresponding to their less than 5% overall similarity.
It is worth noting that, despite high overall nucleotide sequence divergence to Nocturne116, Q54 [
31], the closest relative of Nocturne116 boasts not only similar genome size, organization, and ORF count (26537 bp with predicted 47 ORFs for Q54 versus 25,554 bp with 52 predicted ORFs for Nocturne116), they both also show exactly the same 3′ cohesive genome termini and similar virion morphology (prolate capsids of 56 × 43 nm for Q54 and ~59 × 40 nm for Nocturne116 with 109 nm versus ~112 nm long non-contractile tails, respectively).
Figure 1.
Pairwise genome nucleotide sequence comparison of
Lactococcus phages Nocturne116 and Q54 using BLASTN (
A) and TBLASTX (
B). Genomes are drawn to scale; the scale bar indicates 5000 base pairs. Arrows representing open reading frames point in the direction of transcription and are color-coded based on the function of their putative product according to the legend. Gray boxes represent regions of similarity between genomes and are gradient-colored according to their identity; darker shade of gray represents higher identity. Numbers above selected arrows represent functionally annotated ORFs of Nocturne116 (
Supplementary Table S1). The pairwise comparisons were carried out using EasyFig [
32].
Figure 1.
Pairwise genome nucleotide sequence comparison of
Lactococcus phages Nocturne116 and Q54 using BLASTN (
A) and TBLASTX (
B). Genomes are drawn to scale; the scale bar indicates 5000 base pairs. Arrows representing open reading frames point in the direction of transcription and are color-coded based on the function of their putative product according to the legend. Gray boxes represent regions of similarity between genomes and are gradient-colored according to their identity; darker shade of gray represents higher identity. Numbers above selected arrows represent functionally annotated ORFs of Nocturne116 (
Supplementary Table S1). The pairwise comparisons were carried out using EasyFig [
32].
Out of 52 open reading frames predicted in the genome of Nocturne116, ATG was supposed to serve as a start codon 48 times, TTG—twice, while CTG and GTG each had a single occurrence. Based on the minimum free energy calculations for possible helix formation between 20 bases upstream of the predicted start codon and the last 13 bases of
L. lactis 16s rRNA tail (3’-UUUCCUCCACUAG-5’), 41/52 (78.85%) ORFs were determined to have a Shine-Dalgarno sequence candidate with reasonable binding strength identifiable. No tRNA genes were found in the genome of Nocturne116 (
Supplementary Table S1).
The proteome of Nocturne116 seemed rather interesting: 22 out of 52 predicted proteins had no homologs available in the public amino acid sequence databases, and another 15 ORF product homologs could be found only in the proteome of
Lactococcus phage Q54. Overall, 37/52 (~71%) proteins encoded by Nocturne116 remained without even the slightest functional annotation. Functions for only 15 remaining products could be somewhat reliably assigned/predicted using comparative genomics approaches (
Supplementary Table S1).
Amino acid sequences of four gene products corresponding to evolutionary conserved proteins found in phages (putative DNA polymerase, major capsid protein, terminase, and tail tape measure protein) were chosen as “markers” to inspect the putative evolutionary relationships of Nocturne116 with other publicly available phages, yet again highlighting how distinct Nocturne116 is from other known phages, save for a distant link to phage Q54 (
Figure 1 and
Figure 2,
Supplementary Table S1).
3.2. Open Reading Frame Product Functional Assignments
While we firmly believe that each annotation of a phage product exclusively in silico is a hypothesis made by the respective annotation authors, and different authors could annotate the same product based on the same degree of evidence for its putative functions differently, there seems to be a considerable shift towards the use of phage “auto-annotation” software instead of a more time-consuming so-called “manual expert-based/supervised annotation”, with cases of rough misannotation not being seldom due to relying solely on automated approaches. Although undeniably feasible in cases where numerous phage genomes or their parts need to be annotated and analyzed in a reasonable amount of time (e.g., metagenome/metavirome studies), we find a solely automated approach to phage genome annotation unacceptable for studies describing a single/few novel phages as such an approach is subject to loss of putatively valuable information. For example, there are some cases when a study detailing the determination of function/functions of a particular product or products for an already submitted and annotated complete phage genome sequence gets published, but the annotation does not get updated with the novel insights about the respective ORF product functions elucidated in vitro. While during rigorous manual annotation of phage product functions, such occurrences are readily identifiable through the relevant literature searches, corresponding functional assignment will be missed by auto-annotation software if not explicitly present in the relevant homolog sequence-associated metadata. However, even the phage papers describing the “manual” annotation process often lack the detailed “decision-making” description for their phage ORF-encoded product functional assignments that would show the rationale behind the annotation proposed by the authors to the phage researcher community. This being said, below we have tried to elaborate on the phage protein functional assignment approach for our rather peculiar phage using mainly three free web-based annotation tools (Conserved Domain Database Search, BLASTP, and HHpred), and most importantly, detailing our decision-making process before assigning products with a function.
3.2.1. ORF #5
ORF #5 in the genome of Nocturne116 encodes for a 132 aa long product with a predicted MW of 15.10 kD and is leniently assumed by us to encode a putative Cas system-associated protein. Although homologs of this protein of identical length were found mostly in proteomes of different Lactococcus garviae, L. lactis and some unclassified lactococci strains, some of the Lactococcus-infecting phages seemed to encode for a similar protein as well (12/135 hits). The most detailed annotations available were found to be a “DUF2800 domain-containing protein” for two similar sequences belonging to L. garviae. As of current knowledge, DUF2800 is a member of the CRISPR/Cas system-associated protein Cas4 conserved protein domain superfamily “Cas4_I-A_I-B_I-C_I-D_II-B”, the hit to which was identified when searching for conserved domains in ORF #5 product using CDD search. Although most of our query sequence could be aligned to pfam10926, the coverage spanned only aa positions 251–358 of the 366 aa long conserved domain (CD), which gave a bit score of 34.88 and an E-value of 7.62 × 10−3 according to the current domain model’s position-specific scoring matrix (Pssm-ID:402484). As the evidence for functional assignment of this product is rather weak, and to signify its speculative character, we have opted for the inclusion of “putative” in the corresponding product label while choosing not to ignore the possibility of trying to make a functional assignment.
3.2.2. ORF #9
ORF #9 in the genome of Nocturne116 encodes for a 224 aa long product with a predicted MW of 25.38 kD. All the BLASTP phage hits were exclusive to lactococcal phages and some of the best scoring ones were labeled as “Sak” proteins, the functions of which were previously elucidated in
Lactococcus-infecting phages [
38]. Another type of hits having lesser scores was found with various bacteria, the vast majority of which belonged to either enterococci or lactococci, with annotations corresponding to the “DUF1071 domain-containing protein”.
While it was lucrative to label this product “Sak” protein, the absence of significant similarity was noted in the aa sequence of Lactococcus phage ul36 Sak (ORF252; AAM75760.1), for which the function was determined experimentally. Moreover, ul36 Sak (ORF252) has shown a different conserved domain composition to that of Nocturne116 ORF #9. CDD search for ORF #9 revealed only a presence of DUF1071 domain (pfam06378), for which we could not find even a piece of remote functional evidence. The coverage spanned aa positions 6–170 of ORF #9 to positions 3–148 of 152 aa long conserved domain with a bit score of 176.22 and E-value of 2.20 × 10−56, showing aa “insertions” in the query (ORF #9) when compared to CD, according to Pssm-ID: 399403. While HHpred search querying ORF #9 did, indeed, reveal a hit to Rad52_Rad22 domain (pfam04098) found in ul36 Sak, the hit to DUF1071 domain (pfam06378) was more convincing. It is worth mentioning, however, that DUF1071 can be found in some of the aforementioned BLASTP phage hits labeled as Sak proteins as well, implying differences in the subjective evaluation of similar degrees of evidence while performing the annotation using comparative genomics approaches. As for ORF #9 of Nocturne116, we could not establish a confident link to experimental functional evidence that would show DUF1071 correspondence with Sak protein functions; thus, we have opted to annotate the ORF #9 of Nocturne116 as “protein of unknown function DUF1071”.
3.2.3. ORF #10
ORF #10 in the genome of Nocturne116 encodes for a 147 aa long product with a predicted MW of 16.29 kD. When querying the amino acid sequence of this protein using BLASTP, an overwhelming amount of significant hits can be documented with homologs found in various species of lactococci, all of which are single-stranded DNA-binding proteins. CDD search reveals a plethora of hits to conserved single-stranded DNA-binding protein clusters, with the best match to ssDNA-binding protein cluster PRK07275 from Streptococcaceae. The coverage spanned the complete length of PRK07275 conserved domain, showing two stretches of aa deletions in ORF #10 of Nocturne116, giving a bit score of 207.43 and E-value of 1.01 × 10−60, according to Pssm-ID: 180915. Although the previous observations were enough to make a confident function assignment, HHpred search of ORF #10 has also revealed significant hits to crystal structures of ssDNA-binding proteins of various bacteria, thus complementing the evidence to call the product of Nocturne116 ORF #10 a single-stranded DNA-binding protein.
3.2.4. ORF #14
ORF #14 in the genome of Nocturne116 encodes for a 389 aa long product with a predicted MW of 44.11 kD. BLASTP search using aa sequence of Nocturne116 ORF #14 product has revealed hits from a wide array of different organisms. Topped by hits to
Lactococcus phages (
Figure 2A), similar proteins were also found in various other bacteria from orders
Bacillales and
Lactobacillales, labeled either as DNA polymerases or metallophosphoesterases with or without the “putative” prefix among entries named “hypothetical proteins”. Interestingly, sequences from bacterial species belonging to the genus
Lactococcus were not found among the significant hits at all. CDD search did not reveal even a single significant alignment to any of the conserved domains, while HHpred showed hits to C-terminal metallophosphatase domain of archaea DNA polymerase II, small subunit (cd07386), and structures of the corresponding proteins from
Thermococcus and
Pyrococcus archaea.
Although metallo-dependent phosphatase domains are found in a range of different proteins, we have opted to make an assumption that Nocturne116 ORF #14 is a putative DNA polymerase, taking the genomic context and annotations made by the other Lactococcus phage genome submission authors into account.
3.2.5. ORF #15
ORF #15 in the genome of Nocturne116 encodes for a 169 aa long product with a predicted MW of 18.90 kD. The best scoring BLASTP hit was to a RuvC endonuclease from
Lactococcus phage Q54, which was followed by hits to sequences of similar length labeled either “hypothetical protein” or “crossover junction endodeoxyribonuclease RuvC” from different
Streptococcus species and
Streptococcus phages. CDD search has, indeed, revealed a presence of crossover junction endodeoxyribonuclease RuvC domain (cd16962), albeit the alignment of the query spanned only the C-terminal (aa 2 to 90) of 153 aa long conserved domain and gave a bit score of 45.52 and E-value of 1.58 × 10
−6, according to Pssm-ID: 340813. In the meantime, the best hit from the HHpred query was found to be to a RuvC endonuclease structure from
Lactococcus phage bIL67 (4KTW_A; ref. [
39]) followed by close hits to its counterparts from proteomes of
E. coli,
Thermus thermophilus, and
Pseudomonas aeruginosa. This evidence was deemed to be convincing enough to assign the product of ORF #15 from Nocturne116 with a Holliday junction resolvase function.
3.2.6. ORF #17
ORF #17 in the genome of Nocturne116 encodes for a 314 aa long product with a predicted MW of 35.08 kD. The sole BLASTP hit was to a putative portal protein from
Lactococcus phage Q54 (YP_762589.1). CDD search did not reveal any conserved domains and the HHpred hits, although topped by ten entries corresponding to portal proteins either with or without “probable” prefix, were chosen not to be trusted on the basis of their probabilities (less than or equal to 95.25%) and E-values (more than or equal to 1). A PSI-blast among sequences of viral origin (taxid: 10239) was employed in this case, resulting in hits to “minor structural”, “capsid and scaffold”, as well as more “portal” proteins, either labeled with or without the “putative” prefix. Apart from the aforementioned hit to
Lactococcus phage Q54, the other alignments were rather ambiguous and have shown up in 61% of query coverage with 24.34% identity for “capsid and scaffold protein” of
Lactococcus phage CHPC1242 (QGT52752.1) and ≤ 49% coverage of ≤ 23.68% identity for other hits. It was noted that labeling of these somewhat similar products was inconsistent, although they aligned to the same spot of Nocturne116 ORF #17 product. However, the paper of Fortier and colleagues describing the genome and proteome of phage Q54, the closest known relative of Nocturne116, had employed a mass-spectrometry analysis of structural proteins and revealed that homolog of ORF #17 in Q54 (YP_762589.1) is a structural protein and unlikely to be a major capsid protein (MCP) based on its quantity in the SDS-gel presented therein. In addition, MCP was identified to be encoded by another ORF in the genome of Q54 [
31]. These findings, along with the genomic location of Nocturne116 ORF #17 preceding the better MCP candidate by far, have prompted us to predict the portal protein function for its product due to the lack of a better ORF candidate, as the presence of a portal protein is essential for the replication of tailed bacteriophages.
3.2.7. ORF #18
ORF #18 in the genome of Nocturne116 encodes for a 516 aa long product with a predicted MW of 57.50 kD. The highest scoring BLASTP hit was to major capsid protein (MCP) of phage Q54 (YP_762590.1; 99% query coverage and 64.35% identity), while all the following hits were to proteins of varying length, mostly from either
Lactococcus or
Streptococcus phages (
Figure 2B) and some prophages/prophage remnants of
Listeria innocua,
L. monocytogenes and
Streptococcus pneumoniae, often annotated as major capsid proteins. These “follow-up” hits were notably more different (up to 98% query coverage, although only of 25.37% identity to a second-best hit—MCP of
Lactococcus phage GE1 (YP_009226660.1), and the majority of the alignments were only to the C-terminal part of the queried protein, beginning roughly at aa position 200 onwards). CDD search revealed an HK97 family phage major capsid protein conserved domain (TIGR01554)—a member of the phage capsid domain superfamily (cl27082). Alignment of ORF #18 aa sequence to TIGR01554 spanned aa positions 145–427 corresponding to positions 31–295 of 386 long conserved domain, showing aa stretch “insertions” in both the query and the conserved domain, and gave a bit score of 54.27 and E-value of 8.96 × 10
−8, according to Pssm-ID: 273690. As expected, HHpred did reveal convincing hits to MCP UniProt entries and even to determined capsid structures from other phages. It was also noted that some hits have aligned just to an N or C terminus of Nocturne116 ORF #18 product, with distinct prohead protease hits aligning to N terminus, while MCP hits aligned to C terminus. The best HHpred hit, however, did align to a whole length of our query and was aligned to a capsid polyprotein from
Pseudomonas phage PAJU2 (P85500), for which it was proposed that MCP protein, indeed, might be fused to a prohead protease and autocatalytically cleaved [
40]. Combining these observations, we believe that the presence of prohead protease function for ORF #18 cannot be ruled out; thus, the ORF #18 of Nocturne116 might present yet another occurrence of natural fusion between prohead protease and MCP. However, due to the lack of in vitro experimental evidence for Nocturne116 ORF #18 function, we have decided to view this particular ORF as one of the peculiar major capsid protein encoding cases; therefore, we have labeled the Nocturne116 ORF #18 product as MCP in the annotated genome entry (MW791312.1) while detailing our extended finding herein.
3.2.8. ORF #20
ORF #20 in the genome of Nocturne116 encodes for a 220 aa long product with a predicted MW of 23.73 kD. All the BLASTP hits were revealed to be exclusively to Lactococcus phage proteins of 205–212 aa length. The best scoring hit was to a major tail shaft protein of phage Q54 (YP_762592.1; 93% query coverage of 67.48% identity), with other hits, mostly labeled as putative tail fiber proteins, being less similar (67–93% query coverage of up to 33.76% identity). CDD search did not reveal any conserved domains, while the top two HHpred search hits (both having E-values ≤ 1.5 × 10−31) were to the protein family PF06488 (Siphoviridae phage tail tube proteins including several proteins from Lactococcus lactis) and UniProt entry of tail tube proteins from Lactococcus phage SK1 (O21879). As no substantial evidence to view the product of Nocturne116 ORF #20 as a tail fiber protein was present, “major tail shaft protein” was chosen for product annotation as a synonym of “tail tube protein”, ensuring correspondence to product labels of similar proteins from Lactococcus phages Q54 and GE1 (YP_762592.1 and YP_009226662.1, respectively).
3.2.9. ORF #23
ORF #23 in the genome of Nocturne116 encodes for a 765 aa long product with a predicted MW of 81.45 kD. BLASTP search revealed a 99% query coverage of 63.54% identity hit to the tail tape measure protein (TMP) of phage Q54 (YP_762595.1) followed by numerous hits to proteins from other
Lactococcus-infecting phages (mainly hits from
Ceduovirus genus;
Figure 2D), all of whom seemed to align only to about first 500 aa from the N terminus of Nocturne116 ORF #23 product, giving query coverage of up to 64% and 38.19% identity—these were labeled as either TMP, “tail assembly”, “tail adsorption”, or “tail/minor tail” proteins. CDD search revealed alignment to a cluster of orthologous proteins COG3941, comprising phage tail tape-measure proteins. Alignment of the query spanned aa 86–413 of ORF #23 product to positions 4–344 of 633 aa long domain, showing aa stretch “insertions” for both the query and the hit, giving a bit score of 49.11 and E-value of 9.12 × 10
−6, according to Pssm-ID: 226450.
It was previously noted by other researchers that TMP is usually encoded by the longest ORF in siphophage or myophage genomes and that length of TMP protein roughly corresponds to the tail length of a phage [
41,
42], which explains the observed protein aa length differences among the probable tape measure protein hits to Nocturne116 ORF #23 when employing HHpred search. In addition to the aforementioned evidence, taking into account that ORF #23 is also the longest open reading frame in the genome of Nocturne116, the tape measure protein function was assigned to its product.
3.2.10. ORF #24
ORF #24 in the genome of Nocturne116 encodes for a 99 aa long product with a predicted MW of 11.80 kD. BLASTP search using Nocturne116 ORF #24 product as a query revealed hits from many Lactococcus phages (mainly belonging Ceduovirus genera) and a set of similar proteins from a taxonomically diverse range of bacteria representing Bacteroidetes, Firmicutes, and Proteobacteria phyla, implying immense evolutionary conservatism for ORF #24 product. CDD search revealed an alignment to the HNHc nuclease domain (smart00507). Alignment spanned aa positions 30–79 of ORF #23 product to positions 13–52 of 52 aa long HNHc domain (smart00507), showing aa stretch “insertion” in the query and resulting in a bit score of 35.13 and E-value of 5.26 × 10−4 according to multi-domain Pssm-ID: 214702. The most probable HHpred hit (98.65% probability) represented an HNH endonuclease structure from Geobacillus virus E2 (5H0M_A; Target length 130; E-value of 7.2 × 10−8) followed by a hit to UniProt entry of NinG protein of phage Lambda (PF05766.14; Target length 197; E-value of 1 × 10−7). Although two of the BLASTP-determined most similar proteins from the proteomes of phages Q54 and GE1 were labeled as “hypothetical” (YP_762596.1; 100% query coverage of 65.05% identity and YP_009226667.1; 96% query coverage of 47.42% identity, respectively), some of the less similar sequences from a few other phage hits, as well as the majority of bacterial hit products, were annotated as “HNH endonuclease”. Thus, the product of Nocturne116 ORF #24 was assigned an HNH endonuclease function.
3.2.11. ORF #25
ORF #25 open reading frame in the genome of Nocturne116 encodes for a 542 aa long product with a predicted MW of 62.44 kD. After querying Nocturne116 ORF #25 product aa sequence, BLASTP hits comprised
Lactococcus-infecting phage proteins (
Figure 2C; again, mainly from the
Ceduovirus genera, as with some of the preceding ORF products) and similar proteins found in genomes of bacteria from phylum
Firmicutes, labeled either “terminase” or “terminase large subunit”, with or without “putative” prefix. The best scoring hit was to a terminase protein from phage Q54 (YP_762597.1) which had a query coverage of 94% with an identity of 75.69%, while other hits showed up to 97% query coverage with up to 38.66% identity. It was noted that homologous proteins from the majority of the phages from genus
Ceduovirus were of identical length (518 aa) and showed identical or near-identical alignment scores (300–303), query coverages (95%), and identity percentages (35.63–36.21%). CDD search has shown that Terminase_1 domain (pfam03354) is present in the aa sequence of Nocturne116 ORF #25 product, with an alignment spanning query positions 71–525 to positions 2–452 of 466 aa long conserved domain, giving a bit score of 188.65 and an E-value of 1 × 10
−53 according to Pssm-ID: 281364. As was expected, HHpred revealed convincing hits to phage terminase large subunits or terminase UniProt entries as well. The terminase is responsible for phage genetic material packaging into procapsids, thus being an essential protein for tailed phages [
43].
Although two ORFs encoding, respectively, terminase small (TerS) and large (TerL) subunits, are frequently found in phage genomes, often being in proximity of each other, we have failed to reliably identify a potential TerS-encoding ORF candidate in the genome of Nocturne116 as TerS sequences are not as evolutionarily conserved as their large subunit counterparts. Taking the aforementioned evidence into account, as is usual for cases where no two terminase subunits coding genes can be identified in a phage genome, the product of Nocturne 116 OF #25 was labeled just as a “terminase”.
3.2.12. ORF #27
ORF #27 in the genome of Nocturne116 encodes for a 392 aa long product with a predicted MW of 42.71 kD. CDD and HHpred searches did not reveal any hits and the sole BLASTP hit was found to be a minor structural protein of
Lactococcus phage Q54 (YP_762599.1; query coverage 100%, identity 51.02%, length 391). It is worth noting, however, that this ~43 kD homolog encoded by ORF30 of phage Q54 (YP_762599.1) was originally annotated based on tandem mass spectrometry analysis and could be seen on SDS-PAGE; thus, we believe that the product of Nocturne116 ORF #27 can be reliably annotated as “minor structural protein” as well, owing to the previous in vitro elucidation of phage Q54 structural proteome by Fortier, Bransi, and Moineau [
31].
3.2.13. ORF #31
ORF #31 in the genome of Nocturne116 encodes for a 130 aa long product with a predicted MW of 14.35 kD. HHpred and CDD searches did not reveal any hits, while yet again the sole BLASTP hit was to a similar protein from proteome of Q54—its holin (YP_762602.1; query coverage 94%, identity 45.53%, length 125 aa). TMHMM Server v.2.0 (
http://www.cbs.dtu.dk/services/TMHMM/; accessed on 29 April 2021) was used to check the presence of transmembrane helices in the aa sequence of Nocturne116 ORF #31 product—a characteristic signature of holins. The search revealed the presence of three transmembrane helices at ORF #31 product aa 13–32, 47–69, and 76–95, similar to the findings in the aa sequence of Q54 holin by Fortier and colleagues [
31]. Therefore, the product of Nocturne116 ORF #31 was subsequently annotated as holin, more specifically, as holins are classified based on the number of transmembrane helices, belonging to class I holins [
44].
3.2.14. ORF #32
ORF #32 in the genome of Nocturne116 encodes for a 355 aa long product with a predicted MW of 38.59 kD. Although the majority of the most high-scoring hits were found to a LysM peptidoglycan-binding domain containing proteins from various Lactococcus species, confident hits to several Lactococcus phages were also present, with the highest scoring phage-derived hit being to an endolysin from Lactococcus phage 62,502 (ANT43648.1; query coverage 100%, identity 51.83%, length 429 aa). CDD search revealed alignments to GH25_muramidase (cl10448; glycosyl hydrolase family 25 (GH25) catalytic domain-containing muramidases; positions 1–196/196 of CD aligned to positions 29–222 of the query; bit score of 227.28 and E-value of 5.45 × 10−74, according to Pssm-ID: 415848) and LysM (cd00118; lysin motif; positions 1–45/45 of CD aligned to positions 257–301 of the query; bit score of 69.05 and E-value of 2.35 × 10−15, according to Pssm-ID: 212,030 [Multi-domain]) domains, as well as short alignment to mltD (cl32574; membrane-bound lytic murein transglycosylase D; 345–438/456 of CD aligned to positions 258–346 of the query; bit score of 68.22 and E-value of 1.32 × 10−12, according to Pssm-ID: 182,727 [Multi-domain]) domain. HHpred search results have shown various structures of different phage lysins among the top hits as well. All of the aforementioned evidence, as well as its genomic location next to a predicted holin, suggests that the product of ORF #32 from the Nocturne116 genome is an endolysin.
3.2.15. ORF #35
ORF #35 in the genome of Nocturne116 encodes for a 103 aa long product with a predicted MW of 12.46 kD. BLASTP search revealed numerous hits to either hypothetical proteins or DUF3310 domain-containing proteins from different
Lactococcus phages (belonging to two of the most-populated currently recognized
Lactococcus phage genera, with hits belonging to
Skunavirus being more prominent than those of
Ceduovirus) and some bacteria. The majority of the hits showed similar query coverages in the range of 88–98% with identities in the range of 29.41–40.85%, and although hit protein lengths varied from 72 to 163 aa, most of them were within 101–116 aa length range. CDD search has indicated the presence of a domain of unknown function 3310 (DUF3310; pfam11753) in the query. Alignment spanned positions 15–45 of the query to positions 15–45 of 60 aa long conserved domain, giving a bit score of 34.99 and an E-value of 5.77 × 10
−4, according to Pssm-ID: 403068. While it was noted in Pfam entry that DUF3310 represents a family of conserved bacteriophage proteins, their function yet remains to be elucidated. However, the HHpred search top hit was to a UniProt entry of SaV protein from
Lactococcus phage SK1 (O21894; [
45]), with a close second being to the aforementioned DUF3310 present also in CDD search. SaV protein of SK1, indeed, had an identifiable conserved DUF3310 present in its aa sequence as well, presenting a better alignment of CD to SK1 query (7–64 aa positions of SK1 SaV (O21894) aligned to 5–59 aa positions of 60 aa long DUF3310, showing bit score of 40.38 and an E-value of 8.61 × 10
−6, according to the Pssm-ID: 403068) than the Nocturne116 ORF #35 product query.
Haaber and colleagues have previously demonstrated that SaV is an early-transcription phage protein that is involved in the sensitivity of some lactococcal phages to the then recently described abortive infection mechanism AbiV, and that SaV shows a toxic effect in
L. lactis and
E. coli; thus, it was suggested by the authors of that study that the function of SaV during the phage infection is to shut down or redirect essential host functions [
45,
46]. While it may seem that DUF3310 might, indeed, be a characteristic of
Lactococcus-infecting phage SaV-like proteins (although present in proteins from phages of other hosts as well), in ORF #35 of Nocturne116, only a half of the DUF3310 domain length seems to be conserved and, although already noted by Haaber and colleagues that the middle regions of SaV proteins are conserved even in distantly related phages [
45] (which is yet again highlighted here hypothetically), we have decided to stray away from SaV protein annotation for the product of Nocturne116 ORF #35 and went for a “protein of unknown function DUF3310” annotation for its product.
3.3. The Current State of Lactococcus Phage Genomic Diversity
At the time of writing, there were 349 complete Lactococcus-infecting phage genome entries publicly available at the RefSeq and INSDC, which makes Lactoccocus one of the most popular host genera for phage isolation. These entries span in length from 18,762 (Lactococcus phage asccphi28; EU438902.1) to 132,949 (Lactococcus phage AM4; KY554771.1) base pairs, and, respectively, encode from 28 to 194 products. Deduplication of these 349 complete genome entries using CD-HIT v4.8.1 at 0.95 nucleotide sequence identity threshold (current ICTV BAVS phage species demarcation criterion) resulted in 294 putative phage species, more than a half of which are officially recognized by the ICTV.
It was noted that genera-level taxonomical information associated with these entries listed only two genera of Lactococcus-infecting phages, with genus Skunavirus comprising 214 Lactococcus-infecting phage genomes and 47 for genus Ceduovirus. Respectively, this made 88 Lactococcus-infecting phages still unclassified at the genus level, and we believed that hardly would there be no additional genomic similarity clusters corresponding to additional putative genera among these 88 phages.
The bacteriophage taxonomy is known to be in rapid flux since the affordability of sequencing has grown and the unraveling of phage diversity began happening at unprecedented rates worldwide. Thus, ICTV’s Master Species List 2020.v1 ratified in March, 2021 and published online on 18 May 2021 was consulted for the most up-to-date official taxonomy/classification of Lactococcus phages. The official taxonomy was found to list 158 species and, to our surprise, these 158 species were already scattered among 14 genera in contrast to 2, as seen in the phage genome entry associated metadata, which shows that taxonomical information seen in the public sequence repositories strongly lags behind the officially recognized phage taxonomy overseen by ICTV BAVS. Out of these 14 genera, 7 (Chertseyvirus, Chopinvirus, Negarvirus, Questintvirus, Sandindevirus, Uwajimavirus, Whiteheadvirus) were found to contain just a single species each. While Audreyjarvisvirus had 7 species listed in the viral species master list, Vedamuthuvirus and Teubervirus both had 5, Nevevirus had 4, and Fremauxvirus had 2 species. Skunavirus and Ceduovirus genera had 94 and 34 species in the official taxonomy, respectively.
Based on the annotation of the Nocturne116 complete genome sequence and its genome encoded protein similarity values to those of other phages, it became obvious that Nocturne116 would be a singleton on the genus level, although having a fair amount of proteome homologous to its distant relative, phage Q54 (a single representative of Questintvirus genus). However, its characterization has prompted us to have a look at the overall context of the currently available lactococcal phage genome diversity, with special attention to Lactococcus phage genomes yet “unclassified” at the genus level based on the metadata from respective genome sequences.
This being said, intergenomic similarities between yet “unclassified”
Lactococcus phage genomes and nine randomly selected representatives from the two
Lactococcus phage genera listed within the complete genome entries in public sequence repositories, along with the respective type species (
Skunavirus—
Lactococcus phage sk1,
Ceduovirus—
Lactococcus phage c2), were carried out using VIRIDIC (
n = 108), while proteome comparisons of
Lactococcus-infecting phages were done employing the vConTACT2 pipeline and including all the
Lactococcus-infecting phage genomes available at the time (
n = 349). Both analyses revealed largely coinciding clusters of phages representing novel recently recognized
Lactococcus-infecting phage genera. However, as was expected due to the redundancy of genetic code, clustering based on the proteome was more sensitive and able to find evolutionary links between phages very distantly related based on the genome nucleotide sequence alone (
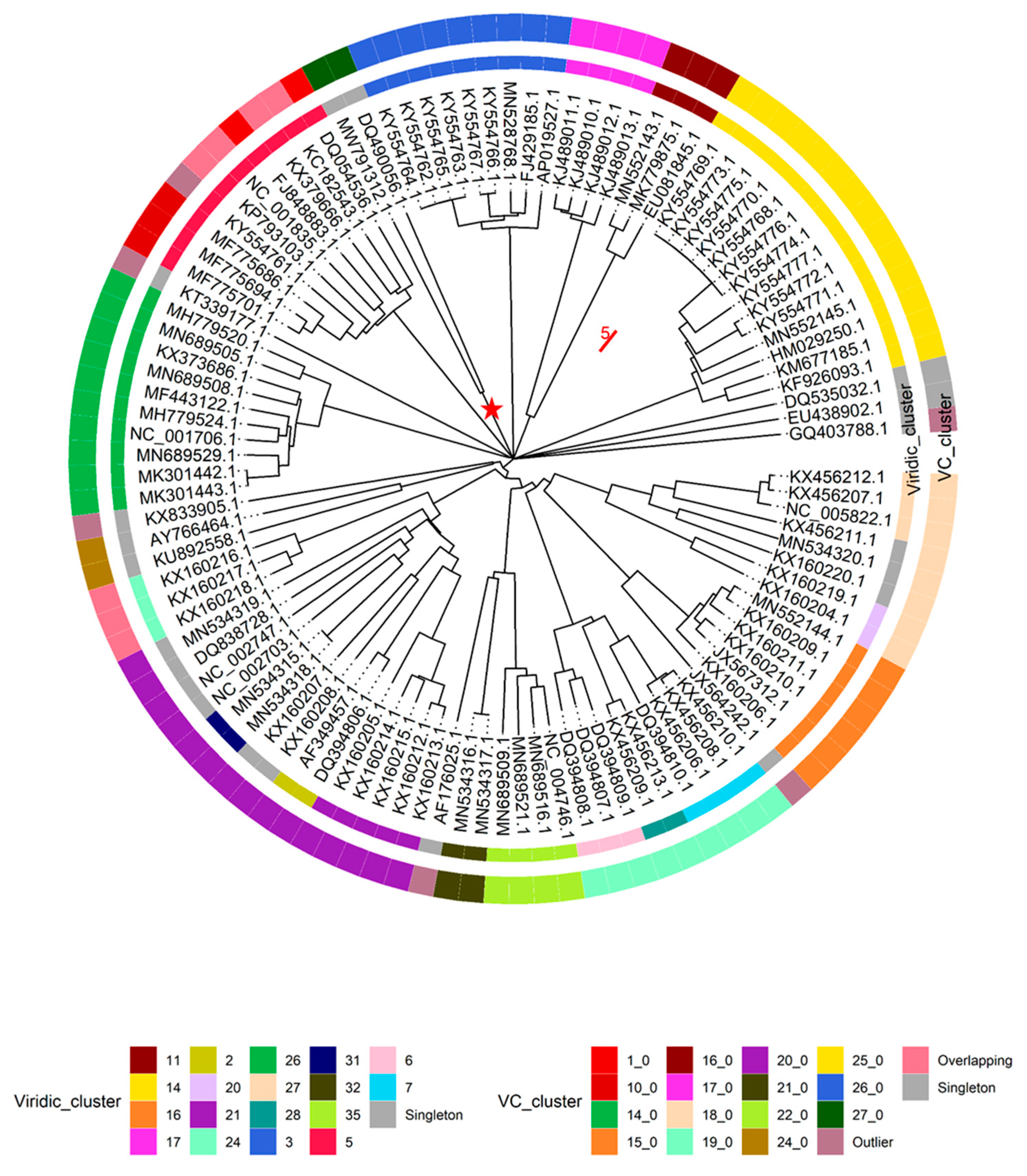
Figure 3,
Supplementary Table S2).
It was also noted that under default settings VIRIDIC did not cluster 9 + 1 selected genomes of
Skunavirus phages (based on genome metadata-associated taxonomic information) in a single cluster at the ICTV BAVS set threshold for phage assignment into the existing genera based on nucleotide sequence similarity (70% sequence similarity [
27]), so the 65% nucleotide sequence similarity threshold was employed to respect the
Skunavirus genus level taxonomy as seen in the respective genome submissions. With this clustering threshold taken into account, none of the “unclassified” viruses (
n = 88) were found to belong to either of the two
Lactococcus phage genera currently recognized in the taxonomical metadata of the submissions, while overall forming 36 clusters (two clusters corresponding to
Skunavirus (VIRIDIC cluster 5) and
Ceduovirus (VIRIDIC cluster 26) genera excluded). Only 16 of the nucleotide sequence similarity clusters consisted of two or more phages, with the other 20 “clusters”, including the genome of Nocturne116, being singletons based on the pairwise genome nucleotide sequence similarity demarcation/clustering at 65% (
Supplementary Table S2). The most populated cluster designated VIRIDIC cluster “14” was found to contain 14 out of the 88 “unclassified” lactococcal phage genomes, corresponding to ICTV recognized genus
Audreyjarvisvirus; VIRIDIC cluster “3” had nine representatives corresponding to genus
Teubervirus; VIRIDIC cluster “16”—six phages from genus
Vedamuthuvirus; VIRIDIC cluster “21”—5 phages yet without standing in the official ICTV taxonomy. The remaining 12 VIRIDIC clusters that were not singletons had between 2 to 4 corresponding phages, respectively (VIRIDIC cluster 17—genera
Nevevirus, VIRIDIC cluster 11—genera
Fremauxvirus).
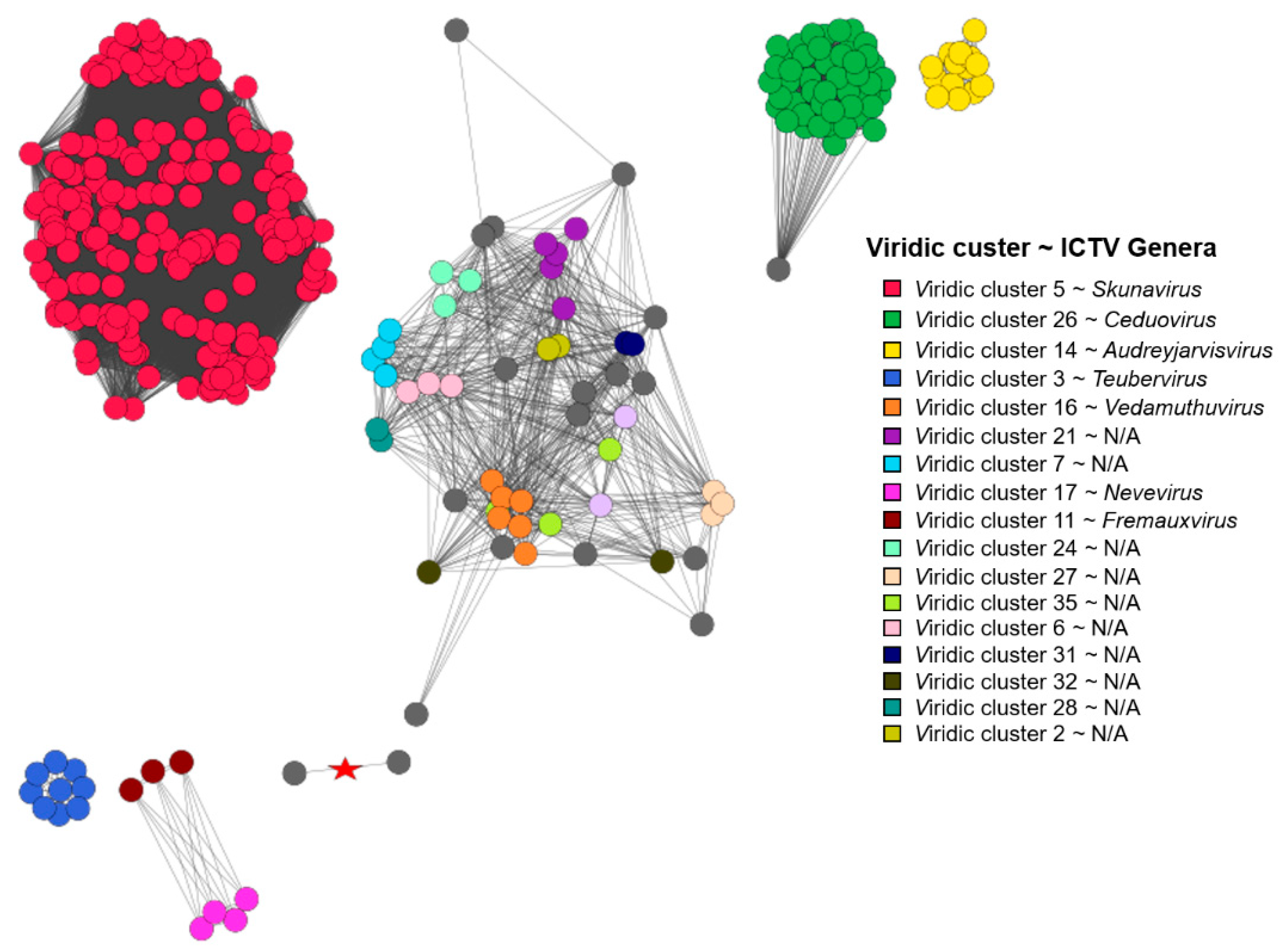
Based on the vConTACT2 clustering of proteomes derived from 349 Lactococcus phages, 252 were found to belong to one of the 17 distinct vConTACT (VC) clusters, 78 of the phages showed overlaps between two VC clusters, while proteomes of nine phages overlapped between three or more VC clusters, eight phages were deemed “outliers”, and two of the phages formed singletons. It was noted that all 47 of the phages belonging to genus Ceduovirus were clustered into a single proteome cluster (VC 14_0), while the situation with skunaviruses was more ambiguous. Out of 214 phages belonging to genus Skunavirus, only 127 were found to be clustered within one of the four proteome clusters (VC1_0—68/127, VC10_0—34/127, VC5_0—22/127, and VC13_0—3/127, respectively), while the majority of the remaining 87 phages identified as belonging to “Skunavirus” genera based on the taxonomy associated with the genome submissions have shown considerable overlaps between two or more proteome clusters, and three of them were “outliers”.
These observations might be either suggestive that the time has come to revise the definition of
Skunavirus genus (at least as was seen from the sequence-associated taxonomy available at INSDC and Refseq at the time of writing), or that the usage possibilities of NCBI taxonomy database [
49] are very limited for phages, as even nine randomly selected presumably
Skunavirus phages failed to make a single cluster around the type species reference strain based on mechanistic 70% nucleotide sequence similarity genus demarcation criterion (as seen from VIRIDIC-generated heatmap [
27],
Supplementary Figure S3). The nonhomogeneity of (presumably)
Skunavirus representatives becomes also evident when comparing the proteome contents of corresponding phages, showing not only well-defined proteome clusters, but a number of phages overlapping their boundaries as well, in contrast to
Ceduovirus, which holds coherent by both nucleotide sequence similarity and proteome content comparisons (
Figure 3).
Combining the results of these comparative analyses, the necessity of the latest changes in the
Lactococcus phage taxonomy recently ratified by ICTV and not yet taken into the account by public biological sequence repositories becomes evident. At least two novel and comparatively populated genera of
Lactococcus-infecting phages distinct from other
Lactococcus phages were recently defined (corresponding to “Viridic_cluster 14 = VC_25_0 ~ ICTV
Audreyjarvisvirus” with 14 representatives and “Viridic_cluster 3 = VC_26_0 ~ ICTV
Teubervirus” with 9 representatives, as seen herein). Other sets of phages corresponding to a less populated novel genera are also easily identifiable from these results (e.g., Viridic_cluster 16 = VC_cluster 15_0 ~ ICTV
Vedamuthuvirus (
n = 6), Viridic_cluster 17 = VC_cluster 17_0 ~ ICTV
Nevevirus (
n = 4), Viridic_cluster 11 = VC_cluster 16_0 ~ ICTV
Fremauxvirus, etc.) as well (
Figure 4).
Some other evolutionary-interconnected, but, nonetheless, distinct enough clusters of
Lactococcus phages presented herein (
Figure 3 and
Figure 4) suggest that the taxonomy of yet unclassified
Lactococcus phages might be worthy of additional attention from ICTV BAVS, while the definition of these putative new genera based on their genomic contents could, however, potentially be hampered by the great variation in the quality of the genome annotation efforts for similar phages by different research groups that does not seem to be related to the genome sizes of phages being annotated (
Supplementary Table S2,
Supplementary Figure S4). Although the temporal differences (scientific knowledge available on the year of annotation and submission of each particular phage) might explain the variation in fractions of unannotated gene products for any given group of related phages to a degree, with older submissions potentially having less of their ORFs functionally annotated, the phage genome functional annotation process should not be taken lightly.
As an additional note, while making revisions to the initial version of this manuscript during the review process, it was found by us that on 15.07.2021, the genus level taxonomy associated with the Lactococcus phage complete genome submissions had finally been updated in public biological sequence repositories to conform with the current official ICTV taxonomy ratified in March 2021 after a lag of a few months.




{kind=link}
{kind=link}
{kind=link}
{kind=link}



