
Behaviour of Listeria monocytogenes and Natural Microflora during the Manufacture of Riojano Chorizo (Spanish Dry Cured Sausage)



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Inoculum Preparation
2.2. Chorizo Formulation and Processing
2.3. Experimental Design
2.4. Physicochemical Analyses
2.5. Microbiological Analyses
2.6. Statistical Analysis
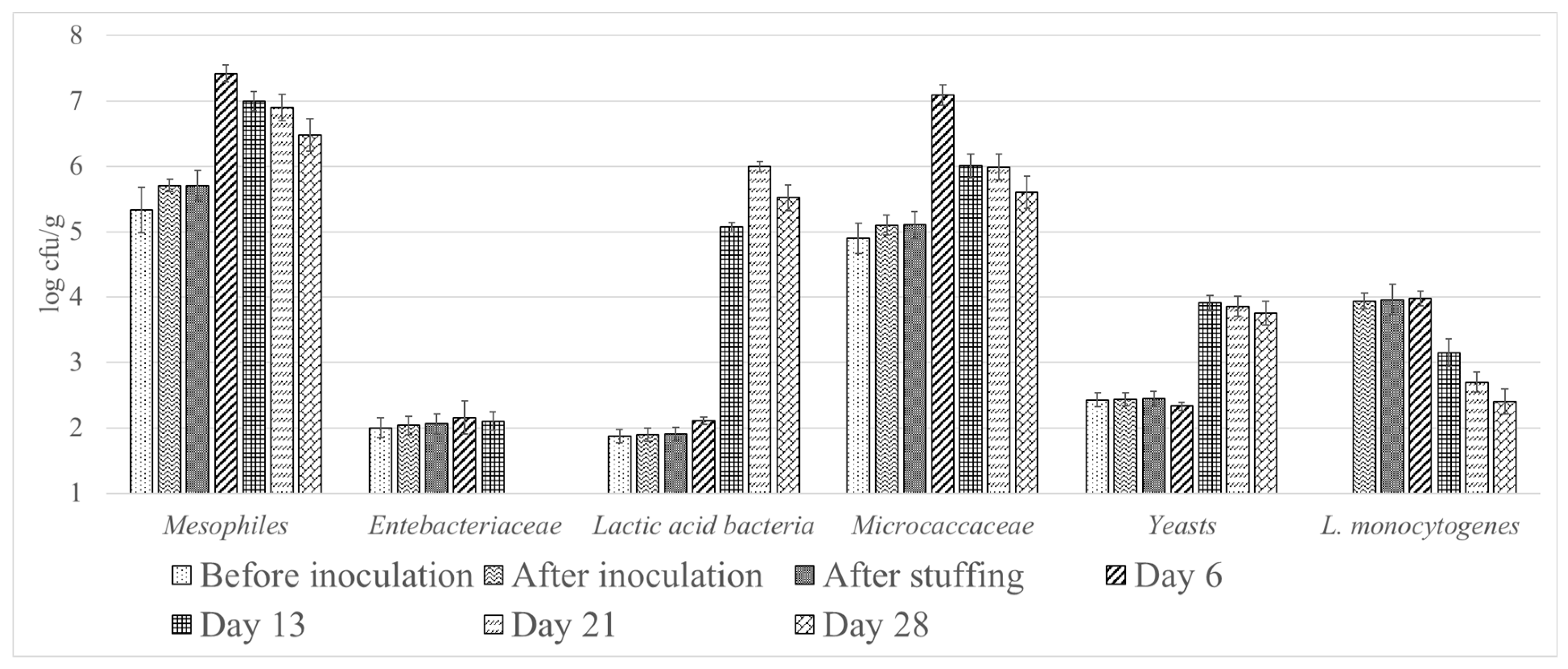
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ordoñez, J.A.; La Hoz, L. Mediterranean products. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 333–348. [Google Scholar]
- Gonzalez-Fandos, M.E.; Sierra, M.; Garcia-Lopez, M.L.; Garcia-Fernandez, C.; Otero, A. The influence of manufacturing and drying conditions on the survival and toxinogenesis of Staphylococcus aureus in two Spanish dry fermented sausages (chorizo and salchichón). Meat Sci. 1999, 52, 411–419. [Google Scholar] [CrossRef]
- Gonzalez-Fandos, E.; Vazquez de Castro, A.; Perez-Arnedo, I. Behabiour of Listeria monocytogenes and other microorganism in slice Riojano chorizo (Spanish dried-cured sausage) during storage under modified atmosphere. Microorganisms 2021, 9, 1384. [Google Scholar] [CrossRef] [PubMed]
- European Commision. Commission Regulation (EC). No 249/2010 of 24 March 2010 entering a name in the register of protected designations of origin and protected geographical indications [Riojano Chorizo (PGI)]. Off. J. Eur. Union 2010, L79, 3–4. [Google Scholar]
- Christieans, S.; Picgirard, L.; Parafita, E.; Lebert, A.; Gregoric, T. Impact of reducing nitrate/nitrite levels on the behavior of Salmonella Typhimurium and Listeria monocytogenes in French dry fermented sausages. Meat Sci. 2018, 137, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Ganan, M.; Hierro, E.; Hospital, X.F.; Barroso, E.; Fernández, M. Use of pulsed light to increase the safety of ready-to-eat cured meat products. Food Control. 2013, 32, 512–517. [Google Scholar] [CrossRef]
- Gomez, D.; Iguácel, L.; Rota, M.; Carramiñana, J.; Ariño, A.; Yangüela, J. Occurrence of Listeria monocytogenes in ready-to-eat meat products and meat processing plants in Spain. Food 2015, 4, 271–282. [Google Scholar] [CrossRef]
- Gonzalez-Fandos, E.; Martínez Loza, J.J.; Maya, N.; Vázquez de Castro, M. Prevalence of Salmonella in Riojano chorizo during processing and final product. In Proceedings of the 3rd International Congress HACCP, Córdoba, Spain, 25 April 2008. [Google Scholar]
- Omer, M.K.; Álvarez-Ordoñez, A.; Prieto, M.; Skjerve, E.; Asehun, T.; Alvseike, A. A Systematic Review of Bacterial Foodborne Outbreaks Related to Red Meat and Meat. Foodborne Pathog. Dis. 2018, 15, 598–611. [Google Scholar] [CrossRef]
- Lücke, F.K.; Zangerl, P. Food safety challenges associated with traditional foods in German-speaking regions. Food Control. 2014, 43, 217–230. [Google Scholar] [CrossRef]
- EFSA. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar]
- Thévenot, D.; Dernburg, A.; Vernozy, C. An updated review of Listeria monocytogenes in the pork meat industry and its products. J. Appl. Microbiol. 2006, 101, 7–17. [Google Scholar] [CrossRef]
- Meloni, D. Presence of Listeria monocytogenes in Mediterranean-style dry fermented sausages. Foods 2015, 4, 34–50. [Google Scholar] [CrossRef]
- Possas, A.; Valdramidis, V.; García-Gimeno, R.M.; Pérez-Rodríguez, F. High hydrostatic pressure processing of sliced fermented sausages: A quantitative exposure assessment for Listeria monocytogenes. Innov. Food Sci. Emerg. Technol. 2019, 52, 406–419. [Google Scholar] [CrossRef]
- Martin, B.; Garriga, M.; Aymerich, T. Prevalence of Salmonella spp. and Listeria monocytogenes at small-scale Spanish factories producing traditional fermented sausages. J. Food Prot. 2011, 174, 812–815. [Google Scholar] [CrossRef]
- Cabedo, L.; Barrot, L.; Canelles, A. Prevalence of Listeria monocytogenes and Salmonella in ready-to-eat food in Catalonia, Spain. J. Food Protect. Prot. 2008, 71, 855–859. [Google Scholar] [CrossRef]
- Hoelzer, K.; Puillot, R.; Dennis, S. Update on Listeria monocytogenes. In Advances in Microbial Food Safety Volume 2; Sofos, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 149–194. [Google Scholar]
- Knudsen, G.M.; Nielsen, J.B.; Marvig, R.L.; Ng, Y.; Worning, P.; Westh, H.; Gram, L. Genome-wide-analyses of Listeria monocytogenes from food processing plants reveal clonal diversity and date the emergence of persisting sequence types. Environ. Microbiol. Rep. 2017, 9, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Lebert, I.; Leroy, S.; Giammarinaro, P.; Lebert, A.; Chacornac, J.P.; Bover-Cid, S.; Vidal-Carou, M.C.; Talon, R. Diversity of microorganisms in the environment and dry fermented sausages of small traditional French processing units. Meat Sci. 2007, 77, 570–579. [Google Scholar] [CrossRef]
- Encinas, J.P.; Sanz, J.J.; Garcia-Lopez, M.L.; Otero, A. Behaviour of Listeria spp. in naturally contaminated chorizo (Spanish fermented sausage). Int. J. Food Microbiol. 1999, 46, 167–171. [Google Scholar] [CrossRef]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–551. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, E.J.; Jackson, K.A.; Johnson, S.D.; Graves, L.M.; Silk, B.J.; Mahon, B.E. Listeriosis outbreaks and associated food vehicles, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Escámez, P.S.F.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Scientific opinion on the Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, 5134. [Google Scholar]
- Roccato, A.; Uyttendaele, M.; Barrucci, F.; Cibin, V.; Favretti, M.; Cereser, A.; Cin, M.D.; Pezzuto, A.; Piovesana, A.; Longo, A.; et al. Artisanal Italian salami and soppresse: Identification of control strategies to manage microbiological hazards. Food Microbiol. 2017, 61, 5–13. [Google Scholar] [CrossRef]
- García-Díez, J.G.; Patarata, L. Behavior of Salmonella spp., Listeria monocytogenes, and Staphylococcus aureus in chouriço de vinho, a dry fermented sausage made from wine-marinated meat. J. Food Prot. 2013, 76, 588–594. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No. 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, L338, 1–26. [Google Scholar]
- Encinas, J.P.; Lopez, T.; Garcia, M.L.; Otero, A.; Moreno, B. Yeast populations on Spanish fermented sausages. Meat Sci. 2000, 54, 203–208. [Google Scholar] [CrossRef]
- Aquilanti, L.; Garofalo, C.; Osimani, A.; Clementi, F. Ecology of lactic acid bacteria and coagulase negative cocci in fermented dry sausages manufactured in Italy and other Mediterranean countries: An overview. Int. Food Res. J. 2016, 23, 429–445. [Google Scholar]
- Al-Zeyara, S.A.; Jarvis, B.; Mackey, B.M. The inhibitory effect of natural microflora of food on growth of Listeria monocytogenes in enrichment broths. Int. J. Food Microbiol. 2011, 145, 98–105. [Google Scholar] [CrossRef]
- Cornu, M.; Billoir, E.; Bergis, H.; Beaufort, A.; Zuliani, V. Modeling microbial competition in food: Application to the behavior of Listeria monocytogenes and lactic acid flora in pork meat products. Food Microbiol. 2011, 28, 639–647. [Google Scholar] [CrossRef]
- Vorst, K.L.; Todd, E.C.D.; Ryser, E.T. Transfer of Listeria monocytogenes during mechanical slicing of turkey breast, bologna, and salami. J. Food Protect. Prot. 2006, 69, 619–696. [Google Scholar] [CrossRef]
- Cava, R.; García-Parra, J.; Ladero, L. Effect of high hydrostatic pressure processing and storage temperature on food safety, microbial counts, colour and oxidative changes of a traditional dry-cured sausage. LWT-Food Sci. Technol. 2020, 28, 109462. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Microbiology of the Food Chain. Horizontal Method for the Enumeration of Microorganisms. Part 2: Colony Count at 30 °C by the Surface Plating Technique; ISO 4833-2:2013; International Organization for Standardization: Geneva, Switzerland, 2013. [Google Scholar]
- International Organization for Standardization (ISO). Meat and Meat Products. Enumeration of Lactic Acid Bacteria. Colony-Count Technique at 30 Degrees C; ISO 15214:1998; International Organization for Standardization: Geneva, Switzerland, 1998. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs. Horizontal Method for the Enumeration of Sulfite-Reducing Bacteria Growing under Anaerobic Conditions; ISO 15213:2003; International Organization for Standardization: Geneva, Switzerland, 2003. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain. Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species). Part 1: Method Using Baird-Parker Agar Medium; ISO 6888-1:2021; International Organization for Standardization: Geneva, Switzerland, 2021. [Google Scholar]
- Simon, A.; Gonzalez-Fandos, E. Influence of modified atmosphere packaging and storage temperature on the sensory and microbiological quality of fresh peeled white asparagus. Food Control. 2011, 22, 369–374. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Microbiology of the Food Chain. Horizontal Method for the Detection and enumeration of Listeria Monocytogenes and of Listeria spp. Part 1: Detection Method; ISO 11290-1:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain. Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and of Listeria spp. Part 2: Enumeration Method; ISO 11290-2:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- González-Fandos, E.; Martínez-Laorden, A.; Pérez-Arnedo, I. Efficacy of combinations of lactic acid and potassium sorbate against Listeria monocytogenes in chicken stored under modified atmospheres. Food Microbiol. 2021, 93, 103596. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Fandos, E.; Herrera, B. Efficacy of acetic acid against Listeria monocytogenes attached in poultry skin during refrigerated storage. Foods 2014, 3, 527–540. [Google Scholar] [CrossRef]
- Fonseca, S.; Cachaldora, A.; Gomez, M.; Franco, I.; Carballo, J. Monitoring the bacterial population dynamics during the ripening of Galician chorizo, a traditional dry fermented Spanish sausage. Food Microbiol. 2013, 33, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, I.; Ammor, S.; Laguet, A.; Labayle, S.; Castanet, V.; Dufour, E.; Talon, R. Microbial ecology of a small-scale facility producing traditional dry sausage. Food Control. 2006, 17, 446–453. [Google Scholar] [CrossRef]
- Stollewerk, K.; Jofré, A.; Comaposada, J.; Ferrini, G.; Garriga, M. Ensuring food safety by an innovative fermented sausage manufacturing system. Food Control. 2011, 22, 1984–1991. [Google Scholar] [CrossRef]
- Thevenot, D.; Delignette-Muller, M.L.; Christieans, S.; Vernozy-Rozand, C. Prevalence of Listeria monocytogenes in 13 dried sausage processing plants and their products. Int. J. Food Microbiol. 2005, 102, 85–94. [Google Scholar] [CrossRef]
- Marcos, B.; Aymerich, T.; Garriga, M. Evaluation of high pressure processing as an additional hurdle to control Listeria monocytogenes and Salmonella enterica in low-acid fermented sausages. J. Food Sci. 2005, 70, M339–M344. [Google Scholar] [CrossRef]
- Tirloni, E.; Di Pietro, V.; Rizzi, G.; Pomilio, F.; Cattaneo, P.; Bernardi, C.; Stella, S. Non-thermal inactivation of Listeria spp. in a typical dry-fermented sausage. Ital. J. Food Saf. 2019, 8, 168–173. [Google Scholar]
- Belleggia, L.; Milanović, V.; Ferrocino, I.; Cocolin, L.; Haouet, M.N.; Scuota, S.; Maoloni, A.; Garofalo, C.; Cardinali, F.; Aquilanti, L.; et al. Is there any still undisclosed biodiversity in Ciauscolo salami? A new glance into the microbiota of an artisan production as revealed by high-throughput sequencing. Meat Sci. 2020, 165, 108128. [Google Scholar] [CrossRef]
- Beriain, M.J.; Gómez, I.; Petri, E.; Insausti, K.; Sarriés, M.V. The effects of olive oil emulsified alginate on the physico-chemical, sensory, microbial, and fatty acid profiles of low-salt, inulin-enriched sausages. Meat Sci. 2011, 88, 189–197. [Google Scholar] [CrossRef]
- Benito, M.J.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Córdoba, M.G. Characterization and selection of autochthonous lactic acid bacteria isolated from traditional Iberian dry-fermented salchichón and chorizo sausages. J. Food Sci. 2007, 72, M193–M201. [Google Scholar] [CrossRef] [PubMed]
- Drosinos, E.H.; Mataragas, M.; Xiraphi, N.; Moschonas, G.; Gaitis, F.; Metaxopoulos, J. Characterization of the microbial flora from a traditional Greek fermented sausage. Meat Sci. 2005, 69, 307–317. [Google Scholar] [CrossRef]
- Fontana, C.; Cocconcelli, P.S.; Vignolo, G. Monitoring the bacterial population dynamics during fermentation of artisanal Argentinean sausages. Int. J. Food Microbiol. 2005, 103, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Metaxopoulos, J.; Vlassi, M.; Pappa, A. Stability and safety of traditional Greek salami. A microbiological ecology study. Int. J. Food Microbiol. 1998, 44, 69–82. [Google Scholar] [CrossRef]
- Cardinali, F.; Milanović, V.; Osimani, A.; Aquilanti, L.; Taccari, M.; Garofalo, C.; Polverigiani, S.; Clementi, F.; Franciosi, E.; Tuohy, K.; et al. Microbial dynamics of model Fabriano-like fermented sausages as affected by starter cultures, nitrates and nitrites. Int. J. Food Microbiol. 2018, 278, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Prado, N.; Sampayo, M.; González, P.; Lombó, F.; Díaz, J. Physicochemical, sensory and microbiological characterization of Asturian Chorizo, a traditional fermented sausage manufactured in Northern Spain. Meat Sci. 2019, 156, 118–124. [Google Scholar] [CrossRef]
- Janssens, M.; Myter, N.; Vuyst, L.; Leroy, F. Community dynamics of coagulase-negative staphylococci during spontaneous artisan-type meat fermentations differ between smoking and moulding treatments. Int. J. Food Microbiol. 2013, 166, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Barron, U.; Cadavez, V.; Pereira, A.P.; Gomes, A.; Araujo, J.P.; Saavedra, M.J.; Dias, T. Relating physicochemical and microbiological safety indicators during processing of linguica, a Portuguese traditional dry-fermented sausage. Food Res. Int. 2015, 78, 50–61. [Google Scholar] [CrossRef][Green Version]
- Hospital, X.F.; Hierro, E.; Fernandez, M. Effect of reducing nitrate and nitrite added to dry fermented sausages on the survival of Salmonella Typhimurium. Food Res. Int. 2014, 62, 410–415. [Google Scholar] [CrossRef]
- Lebert, I.; Leroy, S.; Talon, R. Microorganims in traditional fermented sausages. In Mediterranean products. In Handbook of Fermented Meat and Poultry; Toldrá, F., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 113–124. [Google Scholar]
- Degenhardt, R.; Sant’Anna, E.S. Survival of Listeria monocytogenes in low acid Italian sausage produced under Brazilian conditions. Braz. J. Microbiol. 2007, 38, 309–314. [Google Scholar] [CrossRef]
- Garriga, M.; Marcos, B.; Martin, B.; Veciana-Nogue, M.T.; Bover-Cid, S.; Hugas, M.; Aymerich, T. Starter cultures and high-pressure processing to improve the hygiene and safety of slightly fermented sausages. J. Food Prot. 2005, 68, 2341–2348. [Google Scholar] [CrossRef] [PubMed]
- Branciari, R.; Ortenzi, R.; Roila, R.; Miraglia, D.; Ranucci, D.; Valiani, A. Listeria monocytogenes in soft spreadable salami: Study of the pathogen behaviour and growth prediction during manufacturing process and shelf life. Appl. Sci. 2020, 10, 4438. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Thippareddi, H.; Phebus, R.K.; Marsden, J.L.; Nutsch, L. Validation of a traditional Italian-style salami manufacturing process for control of Salmonella and Listeria monocytogenes. J. Food Prot. 2006, 69, 794–800. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Novelli, E.; Santo, D.L.; Balzan, S.; Cardazzo, B.; Spolaor, D.; Lombardi, A.; Carraro, L.; Fasolato, L. Analysis of process factors of dry fermented salami to control Listeria monocytogenes. Ital. J. Food Saf. 2017, 6, 6184. [Google Scholar] [CrossRef]
- Mataragas, M.; Bellio, A.; Rovetto, F.; Astegiano, S.; Greci, C.; Hertel, C.; Decastelli, L.; Cocolin, L. Quantification of persistence of the food-borne pathogens Listeria monocytogenes and Salmonella enterica during manufacture of Italian fermented sausages. Food Control. 2015, 47, 552–559. [Google Scholar] [CrossRef]
- Mataragas, M.; Bellio, A.; Rovetto, F.; Astegiano, S.; Decastelli, L.; Cocolin, L. Risk-based control of food-borne pathogens Listeria monocytogenes and Salmonella enterica in the Italian fermented sausages Cacciatore and Felino. Meat Sci. 2015, 103, 19–45. [Google Scholar] [CrossRef] [PubMed]
- Gounadaki, A.; Skandamis, P.; Drosinos, E.H.; Nychas, G.J.E. Survival of Listeria monocytogenes during the fermentation and ripening process of sausages. In Proceedings of the 1st Hellenic Symposium in Food Biotechnology and Technology; Association of Greek Chemists and Hellenic Association of Chemical Engineers: Athens, Greece, 2015; pp. 436–441. [Google Scholar]
- Ruggeri, A.; Carraro, V.; Succa, S.; Meloni, B.; Sanna, A.; Sanna, C.; Espa, A.; Pinna, A.; Carrucciu, G.; Grosso, M.; et al. Influence of water activity on Listeria monocytogenes growth in “Salsiccia Sarda” fermented sausage. Ital. J. Food Sci. 2018, 30, 75–89. [Google Scholar]
- Campanini, M.; Pedrazzoni, I.; Barbuti, S.; Baldini, P. Behaviour of Listeria monocytogenes during the maturation of naturally and artificially contaminated salami: Effect of lactic-acid bacteria starter cultures. Int. J. Food Microbiol. 1993, 20, 169–175. [Google Scholar] [CrossRef]
- Papamanoli, E.; Kotzekidou, P.; Tzanetakis, N.; Litopoulou-Tzanetaki, E. Characterization of Micrococcaceae isolated from dry fermented sausage. Food Microbiol. 2002, 19, 441–449. [Google Scholar] [CrossRef]
- González-Fandos, M.E.; Sierra, M.L.; García-López, M.L.; Otero, A.; Sanz, J.; Moreno, B. Staphylococcal growth and enterotoxin production in the presence of meat cultures (non LAB). Meat Sci. 1996, 43, 255–263. [Google Scholar] [CrossRef]
- Vilani, F.; Sannino, L.; Moschetti, G.; Mauriello, G.; Pepe, O.; Cocchhieri, R.; Coppoloa, S. Partial characterization of an antagonistic substance produced by Staphylococcus xylosus 1E and determination of the effectiveness of the producer to inhibit Listeria monocytogenes in Italian sausages. Food Microbiol. 1997, 14, 555–566. [Google Scholar] [CrossRef]
- Goerges, S.; Aigner, U.; Silakowski, B.; Scherer, S. Inhibition of Listeria monocytogenes by food-borne yeasts. Appl. Environ. Microbiol. 2006, 72, 313–318. [Google Scholar] [CrossRef]
- Martínez, G.N.; Bastida, P.; Castillo, J.; Ros, G.; Nieto, G. Green Alternatives to Synthetic Antioxidants, Antimicrobials, Nitrates, and Nitrites in Clean Label Spanish Chorizo. Antioxidants 2019, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Doyle, M.P.; Cassens, R.G.; Schoeni, J.L. Fate of Listeria monocytogenes in tissues of experimentally infected cattle and in hard salami. Appl. Environ. Microbiol. 1988, 54, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Thevenot, D.; Delignette-Muller, M.L.; Christieans, S.; Vernozy-Rozand, C. Fate of Listeria monocytogenes in experimentally contaminated French sausages. Int. J. Food Microbiol. 2005, 101, 189–200. [Google Scholar] [CrossRef]


{kind=link}
| Stage | pH | Water Activity | Moisture (%) |
|---|---|---|---|
| Sausage mixture before inoculation (day 0) | 5.90 ± 0.01 a | 0.960 ± 0.005 a | 51.57 ± 0.12 a |
| Sausage mixture after inoculation (day 0) | 5.92 ± 0.01 a | 0.969 ± 0.002 a | 51.68 ± 0.15 a |
| After stuffing (day 0) | 5.93 ± 0.02 a | 0.965 ± 0.003 a | 52.00 ± 0.22 a |
| Day 6 of drying | 5.88 ± 0.03 a | 0.942 ± 0.004 b | 46.60 ± 0.54 b |
| Day 13 of drying | 5.72 ± 0.04 b | 0.912 ± 0.005 c | 39.94 ± 0.65 c |
| Day 21 of drying | 5.78 ± 0.02 b | 0.860 ± 0.005 d | 33.96 ± 0.52 d |
| Day 28 of drying | 5.79 ± 0.01 b | 0.793 ± 0.004 e | 25.00 ± 0.62 e |
| Microoganism | ʎ | µmax | R2 | SE (Fit) |
|---|---|---|---|---|
| L. monocytogenes | 5.212 | −0.093 | 0.910 | 0.216 |
| Lactic acid bacteria | 6.539 | 0.500 a | 0.970 | 0.339 |
| Micrococcaceae | 0.899 | 0.339 a | 0.981 | 0.131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Fandos, E.; Vazquez de Castro, M.; Martinez-Laorden, A. Behaviour of Listeria monocytogenes and Natural Microflora during the Manufacture of Riojano Chorizo (Spanish Dry Cured Sausage). Microorganisms 2021, 9, 1963. https://doi.org/10.3390/microorganisms9091963
Gonzalez-Fandos E, Vazquez de Castro M, Martinez-Laorden A. Behaviour of Listeria monocytogenes and Natural Microflora during the Manufacture of Riojano Chorizo (Spanish Dry Cured Sausage). Microorganisms. 2021; 9(9):1963. https://doi.org/10.3390/microorganisms9091963
Chicago/Turabian StyleGonzalez-Fandos, Elena, Maria Vazquez de Castro, and Alba Martinez-Laorden. 2021. "Behaviour of Listeria monocytogenes and Natural Microflora during the Manufacture of Riojano Chorizo (Spanish Dry Cured Sausage)" Microorganisms 9, no. 9: 1963. https://doi.org/10.3390/microorganisms9091963
APA StyleGonzalez-Fandos, E., Vazquez de Castro, M., & Martinez-Laorden, A. (2021). Behaviour of Listeria monocytogenes and Natural Microflora during the Manufacture of Riojano Chorizo (Spanish Dry Cured Sausage). Microorganisms, 9(9), 1963. https://doi.org/10.3390/microorganisms9091963