Pegbovigrastim Treatment around Parturition Enhances Postpartum Immune Response Gene Network Expression of whole Blood Leukocytes in Holstein and Simmental Cows

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Treatments, and PAXgene Blood Sampling
2.2. RNA Extraction, cDNA Synthesis, Target Genes, and Quantitative PCR
2.3. RT-PCR Data Analysis
2.4. Statistical Analysis
3. Results
3.1. Expression of Genes Involved in the TLR Pathway and Immune Mediation
3.2. Expression of Genes Involved in Cell Migration and Adhesion
3.3. Expression of Genes Involved in Antimicrobial Functions
3.4. Expression of Genes Involved in the Inflammatory Response
3.5. Expression of Genes Involved in the Oxidative Stress and Leukotrienes
4. Discussion
5. Study Limitation
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kehrli, M.E.; Nonnecke, B.J.; Wood, R.L.; Roth, J.A. In vivo effects of a thymosin α1-containing colostral whey product on neutrophils and lymphocytes from lactating cows without and with experimentally induced Staphylococcus aureus mastitis. Vet. Immunol. Immunopathol. 1989, 20, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrli, M.E.; Nonnecke, B.J.; Roth, J.A. Alterations in bovine neutrophil function during the periparturient period. Am. J. Vet. Res. 1989, 50, 207–214. [Google Scholar] [PubMed]
- Batistel, F.; Arroyo, J.M.; Garces, C.I.M.; Trevisi, E.; Parys, C.; Ballou, M.A.; Cardoso, F.C.; Loor, J.J. Ethyl-cellulose rumen-protected methionine alleviates inflammation and oxidative stress and improves neutrophil function during the periparturient period and early lactation in Holstein dairy cows. J. Dairy Sci. 2017, 101, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Drackley, J.K. Biology of Dairy Cows during the Transition Period: The Final Frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Loor, J.J.; Bionaz, M.; Drackley, J.K. Systems Physiology in Dairy Cattle: Nutritional Genomics and Beyond. Annu. Rev. Anim. Biosci. 2013, 1, 365–392. [Google Scholar] [CrossRef]
- Kehrli, M.E.; Goff, J.P.; Stevens, M.G.; Boone, T.C. Effects of Granulocyte Colony-Stimulating Factor Administration to Periparturient Cows on Neutrophils and Bacterial Shedding. J. Dairy Sci. 1991, 74, 2448–2458. [Google Scholar] [CrossRef]
- Nagata, S.; Fukunaga, R. Granulocyte colony-stimulating factor and its receptor. Prog. Growth Factor Res. 1991, 3, 131–141. [Google Scholar] [CrossRef]
- Hercus, T.R.; Thomas, D.; Guthridge, M.A.; Ekert, P.G.; King-Scott, J.; Parker, M.W.; Lopez, A.F. The granulocyte-macrophage colony-stimulating factor receptor: Linking its structure to cell signaling and its role in disease. Blood 2009, 114, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Goff, J.P.; Canning, P.; Wang, C.; Roth, J.A. Effect of recombinant bovine granulocyte colony-stimulating factor covalently bound to polyethylene glycol injection on neutrophil number and function in periparturient dairy cows. J. Dairy Sci. 2014, 97, 4842–4851. [Google Scholar] [CrossRef] [Green Version]
- McDougall, S.; LeBlanc, S.J.; Heiser, A. Effect of prepartum energy balance on neutrophil function following pegbovigrastim treatment in periparturient cows. J. Dairy Sci. 2017, 100, 7478–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, P.S.D.; Madsen-Bouterse, S.A.; Rosa, G.J.M.; Sipkovsky, S.; Ren, X.; Almeida, P.E.; Kruska, R.; Halgren, R.G.; Barrick, J.L.; Burton, J.L. Analysis of the bovine neutrophil transcriptome during glucocorticoid treatment. Physiol. Genom. 2006, 28, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Moyes, K.M.; Graugnard, D.E.; Khan, M.J.; Mukesh, M.; Loor, J.J. Postpartal immunometabolic gene network expression and function in blood neutrophils are altered in response to prepartal energy intake and postpartal intramammary inflammatory challenge. J. Dairy Sci. 2014, 97, 2165–2177. [Google Scholar] [CrossRef] [PubMed]
- Crookenden, M.A.; Heiser, A.; Murray, A.; Dukkipati, V.S.R.; Kay, J.K.; Loor, J.J.; Meier, S.; Mitchell, M.D.; Moyes, K.M.; Walker, C.G.; et al. Parturition in dairy cows temporarily alters the expression of genes in circulating neutrophils. J. Dairy Sci. 2016, 99, 6470–6483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiser, A.; LeBlanc, S.J.; McDougall, S. Pegbovigrastim treatment affects gene expression in neutrophils of pasture-fed, periparturient cows. J. Dairy Sci. 2018, 101, 8194–8207. [Google Scholar] [CrossRef]
- Lopreiato, V.; Minuti, A.; Morittu, V.M.; Britti, D.; Piccioli-Cappelli, F.; Loor, J.J.; Trevisi, E. Short communication: Inflammation, migration, and cell-cell interaction-related gene network expression in leukocytes is enhanced in Simmental compared with Holstein dairy cows after calving. J. Dairy Sci. 2019, 103, 1908–1913. [Google Scholar] [CrossRef]
- Lopreiato, V.; Minuti, A.; Trimboli, F.; Britti, D.; Morittu, V.M.; Cappelli, F.P.; Loor, J.J.; Trevisi, E. Immunometabolic status and productive performance differences between periparturient Simmental and Holstein dairy cows in response to pegbovigrastim. J. Dairy Sci. 2019, 102, 9312–9327. [Google Scholar] [CrossRef]
- Sandri, M.; Stefanon, B.; Loor, J.J. Transcriptome profiles of whole blood in Italian Holstein and Italian Simmental lactating cows diverging for genetic merit for milk protein. J. Dairy Sci. 2015, 98, 6119–6127. [Google Scholar] [CrossRef]
- Pošćić, N.; Montanari, T.; D’Andrea, M.; Licastro, D.; Pilla, F.; Ajmone-Marsan, P.; Minuti, A.; Sgorlon, S. Breed and adaptive response modulate bovine peripheral blood cells’ transcriptome. J. Anim. Sci. Biotechnol. 2017, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Franzke, A. The role of G-CSF in adaptive immunity. Cytokine Growth Factor. Rev. 2006, 17, 235–244. [Google Scholar]
- Powell, E.J.; Reinhardt, T.A.; Casas, E.; Lippolis, J.D. The effect of pegylated granulocyte colony-stimulating factor treatment prior to experimental mastitis in lactating Holsteins. J. Dairy Sci. 2018, 101, 8182–8193. [Google Scholar] [CrossRef] [PubMed]
- Taraktsoglou, M.; Szalabska, U.; Magee, D.A.; Browne, J.A.; Sweeney, T.; Gormley, E.; MacHugh, D.E. Transcriptional profiling of immune genes in bovine monocyte-derived macrophages exposed to bacterial antigens. Vet. Immunol. Immunopathol. 2011, 140, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen Recognition by the Innate Immune System. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, K.; Roesler, J.; Emmendoerffer, A.; Elsner, J.; Welte, K. Functional features of neutrophils induced by G-CSF and GM-CSF treatment: Differential effects and clinical implications. Leukemia 1997, 11, 466–478. [Google Scholar] [CrossRef] [Green Version]
- Kurt-Jones, E.A.; Mandell, L.; Whitney, C.; Padgett, A.; Gosselin, K.; Newburger, P.E.; Finberg, R.W. Role of Toll-like receptor 2 (TLR2) in neutrophil activation: GM-CSF enhances TLR2 expression and TLR2-mediated interleukin 8 responses in neutrophils. Blood 2002, 100, 1860–1868. [Google Scholar] [CrossRef] [Green Version]
- Buzzeo, M.P.; Yang, J.; Casella, G.; Reddy, V. Hematopoietic stem cell mobilization with G-CSF induces innate inflammation yet suppresses adaptive immune gene expression as revealed by microarray analysis. Exp. Hematol. 2007, 35, 1456–1465. [Google Scholar] [CrossRef]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Schmidt, S.; Moser, M.; Sperandio, M. The molecular basis of leukocyte recruitment and its deficiencies. Mol. Immunol. 2013, 55, 49–58. [Google Scholar] [CrossRef]
- Fukuzono, S.; Kato, T.; Fujita, H.; Watanabe, N.; Kitagawa, S. Granulocyte colony-stimulating factor negatively regulates Toll-like receptor agonist-induced cytokine production in human neutrophils. Arch. Biochem. Biophys. 2010, 495, 144–151. [Google Scholar] [CrossRef]
- Kim, S.O.; Sheikh, H.I.; Ha, S.-D.; Martins, A.; Reid, G. G-CSF-mediated inhibition of JNK is a key mechanism for Lactobacillus rhamnosus-induced suppression of TNF production in macrophages. Cell. Microbiol. 2006, 8, 1958–1971. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Sumagin, R.; Prizant, H.; Lomakina, E.; Waugh, R.E.; Sarelius, I.H. LFA-1 and Mac-1 Define Characteristically Different Intralumenal Crawling and Emigration Patterns for Monocytes and Neutrophils In Situ. J. Immunol. 2010, 185, 7057–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmi, M.; Jalkanen, S. How Do Lymphocytes Know Where to Go: Current Concepts and Enigmas of Lymphocyte Homing; Academic Press: Cambridge, MA, USA, 1997; pp. 139–218. [Google Scholar]
- Springer, T.A. Traffic signals for lymphocyte recirculation and leukocyte emigration: The multistep paradigm. Cell 1994, 76, 301–314. [Google Scholar] [CrossRef]
- Hanthorn, C.J.; Dewell, G.A.; Dewell, R.D.; Cooper, V.L.; Wang, C.; Plummer, P.J.; Lakritz, J. Serum concentrations of haptoglobin and haptoglobin-matrix metalloproteinase 9 (Hp-MMP 9) complexes of bovine calves in a bacterial respiratory challenge model. BMC Vet. Res. 2014, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lohr, K.M.; Kurth, C.A.; Xie, D.L.; Seyer, J.M.; Homandberg, G.A. The amino-terminal 29- and 72-Kd fragments of fibronectin mediate selective monocyte recruitment. Blood 1990, 76, 2117–2124. [Google Scholar] [CrossRef]
- Lee, M.; Lee, Y.; Song, J.; Lee, J.; Chang, S.-Y. Tissue-specific Role of CX 3 CR1 Expressing Immune Cells and Their Relationships with Human Disease. Immune Netw. 2018, 18. [Google Scholar] [CrossRef]
- Gonen, E.; Nedvetzki, S.; Naor, D.; Shpigel, N.Y. CD44 is highly expressed on milk neutrophils in bovine mastitis and plays a role in their adhesion to matrix and mammary epithelium. Vet. Res. 2008, 39, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Teng, T.S.; Ji, A.L.; Ji, X.Y.; Li, Y.Z. Neutrophils and immunity: From bactericidal action to being conquered. J. Immunol. Res. 2017, 2017. [Google Scholar] [CrossRef]
- Trimboli, F.; Morittu, V.M.; Di Loria, A.; Minuti, A.; Spina, A.A.; Piccioli-Cappelli, F.; Trevisi, E.; Britti, D.; Lopreiato, V. Effect of Pegbovigrastim on Hematological Profile of Simmental Dairy Cows during the Transition Period. Animal 2019, 9, 841. [Google Scholar] [CrossRef] [Green Version]
- Blaschitz, A.; Gauster, M.; Fuchs, D.; Lang, I.; Maschke, P.; Ulrich, D.; Karpf, E.; Takikawa, O.; Schimek, M.G.; Dohr, G.; et al. Vascular endothelial expression of indoleamine 2,3-dioxygenase 1 forms a positive gradient towards the feto-maternal interface. PLoS ONE 2011, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Joo, Y.D.; Lee, S.M.; Lee, S.W.; Lee, W.S.; Lee, S.M.; Park, J.K.; Choi, I.W.; Park, S.G.; Choi, I.; Seo, S.K. Granulocyte colony-stimulating factor-induced immature myeloid cells inhibit acute graft-versus-host disease lethality through an indoleamine dioxygenase-independent mechanism. Immunology 2009, 128, e632–e640. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, G.; Procoli, A.; Mariotti, A.; Corallo, M.; Perillo, A.; Danese, S.; De Cristofaro, R.; Scambia, G.; Rutella, S. Effects of pegylated G-CSF on immune cell number and function in patients with gynecological malignancies. J. Transl. Med. 2010, 8, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rado, T.A.; Bollekens, J.; St Laurent, G.; Parker, L.; Benz, E.J. Lactoferrin biosynthesis during granulocytopoiesis. Blood 1984, 64, 1103–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bainton, D.F. Neutrophil Granules. Br. J. Haematol. 1975, 29, 17–22. [Google Scholar] [CrossRef]
- Sanchez, L.; Calvo, M.; Brock, J.H. Biological role of lactoferrin. Arch. Dis. Child. 1992, 67, 657–661. [Google Scholar] [CrossRef] [Green Version]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Influence of body condition score on relationships between metabolic status and oxidative stress in periparturient dairy cows. J. Dairy Sci. 2005, 88, 2017–2026. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative Stress, Antioxidants, and Animal Function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Markers of oxidative status in plasma and erythrocytes of transition dairy cows during hot season. J. Dairy Sci. 2002, 85, 2173–2179. [Google Scholar] [CrossRef] [Green Version]
- Piccinini, R.; Binda, E.; Belotti, M.; Casirani, G.; Zecconi, A. The evaluation of non-specific immune status of heifers in field conditions during the periparturient period. Vet. Res. 2004, 35, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Rubin, B.B.; Downey, G.P.; Koh, A.; Degousee, N.; Ghomashchi, F.; Nallan, L.; Stefanski, E.; Harkin, D.W.; Sun, C.X.; Smart, B.P.; et al. Cytosolic phospholipase A2-α is necessary for platelet-activating factor biosynthesis, efficient neutrophil-mediated bacterial killing, and the innate immune response to pulmonary infection: cPLA2-α does not regulate neutrophil NADPH oxidase activity. J. Biol. Chem. 2005, 280, 7519–7529. [Google Scholar] [CrossRef] [Green Version]
- Zipser, R.D.; Laffi, G. Prostaglandins, thromboxanes and leukotrienes in clinical medicine. West. J. Med. 1985, 143, 485–497. [Google Scholar] [PubMed]
- Bottons, G.D.; Adams, H.R. Involvement of prostaglandins and leukotrienes in the pathogenesis of endotoxemia and sepsis. J. Am. Vet. Med. Assoc. 1992, 200, 1842–1848. [Google Scholar]
- Williams, K.I.; Higgs, G.A. Eicosanoids and inflammation. J. Pathol. 1988, 156, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, R.; Zuo, X.; Jaoude, J.; Mao, F.; Colby, J.; Shureiqi, I. ALOX15 as a suppressor of inflammation and cancer: Lost in the link. Prostaglandins Lipid Mediat. 2017, 132, 77–83. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Values | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed 1 | Parity 2 | Group X | ||||||||||
| Target | Group | H | S | P | M | SEM3 | Group | Breed | Parity | Breed | Parity | Breed × Parity |
| CD14 * | CTR | 4.33 | 4.11 | 4.20 | 4.25 | 0.17 | < 0.01 | 0.39 | 0.95 | 0.33 | 0.70 | 0.26 |
| PEG | 4.74 | 4.75 | 4.76 | 4.72 | ||||||||
| CD16 | CTR | 89.13 | 111.84 | 104.90 | 96.07 | 21.28 | < 0.01 | 0.71 | 0.58 | 0.06 | 0.96 | 0.33 |
| PEG | 178.66 | 145.28 | 165.63 | 158.31 | ||||||||
| MYD88 * | CTR | 5.16 | 5.23 | 5.29 | 5.10 | 0.16 | < 0.01 | 0.22 | 0.04 | 0.07 | 0.68 | 0.91 |
| PEG | 6.38 | 6.03 | 6.34 | 6.06 | ||||||||
| TLR2 * | CTR | 3.95 | 3.74 | 3.93 | 3.76 | 0.13 | < 0.01 | < 0.01 | 0.02 | 0.40 | 0.59 | 0.64 |
| PEG | 4.78 | 4.42 | 4.73 | 4.47 | ||||||||
| TLR4 * | CTR | −1.64 | −1.61 | −1.38 | −1.87 | 0.70 | < 0.01 | 0.32 | 0.20 | 0.30 | 0.81 | 0.14 |
| PEG | 0.31 | −0.67 | 0.18 | −0.55 | ||||||||
| p-Values | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed 1 | Parity 2 | Group X | ||||||||||
| Target | Group | H | S | P | M | SEM 3 | Group | Breed | Parity | Breed | Parity | Breed × Parity |
| CCR2 * | CTR | 3.58 | 3.60 | 3.53 | 3.65 | 0.18 | 0.27 | 0.53 | 0.36 | 0.66 | 0.91 | 0.49 |
| PEG | 3.66 | 3.79 | 3.68 | 3.78 | ||||||||
| CX3CR1 | CTR | 17.57 | 24.85 | 20.31 | 22.11 | 3.32 | 0.70 | 0.02 | 0.34 | 0.48 | 0.86 | 0.78 |
| PEG | 18.31 | 22.36 | 19.02 | 21.65 | ||||||||
| ITGB2 | CTR | 196.64 | 229.40 | 209.78 | 216.26 | 20.67 | < 0.01 | 0.05 | 0.70 | 0.72 | 0.94 | 0.98 |
| PEG | 311.46 | 334.14 | 320.59 | 325.01 | ||||||||
| ITGAL4 | CTR | 45.98 | 51.49 | 50.17 | 47.31 | 6.08 | < 0.01 | 0.46 | 0.47 | 0.04 | 0.97 | 0.92 |
| PEG | 72.02 | 60.33 | 67.79 | 64.57 | ||||||||
| TLN1 | CTR | 236.39 | 252.30 | 256.96 | 231.73 | 27.71 | < 0.01 | 0.35 | 0.20 | 0.07 | 0.89 | 0.81 |
| PEG | 376.92 | 328.25 | 362.71 | 342.45 | ||||||||
| SELL * | CTR | 6.25 | 6.26 | 6.32 | 6.19 | 0.13 | < 0.01 | 0.09 | 0.05 | 0.06 | 0.55 | 0.91 |
| PEG | 7.29 | 6.97 | 7.25 | 7.02 | ||||||||
| SELPLG *,4 | CTR | 5.44 | 5.61 | 5.59 | 5.45 | 0.14 | < 0.01 | 0.70 | 0.13 | 0.03 | 0.90 | 0.67 |
| PEG | 6.48 | 6.23 | 6.43 | 6.27 | ||||||||
| CD44 | CTR | 160.76 | 202.64 | 172.41 | 190.99 | 15.79 | < 0.01 | < 0.01 | 0.93 | 0.28 | 0.11 | 0.70 |
| PEG | 236.07 | 254.13 | 253.50 | 236.70 | ||||||||
| LGALS8 * | CTR | 3.57 | 3.70 | 3.67 | 3.60 | 0.17 | 0.47 | 0.60 | 0.68 | 0.51 | 0.87 | 0.25 |
| PEG | 3.73 | 3.71 | 3.73 | 3.70 | ||||||||
| p-Values | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed1 | Parity2 | Group X | ||||||||||
| Target | Group | H | S | P | M | SEM3 | Group | Breed | Parity | Breed | Parity | Breed × Parity |
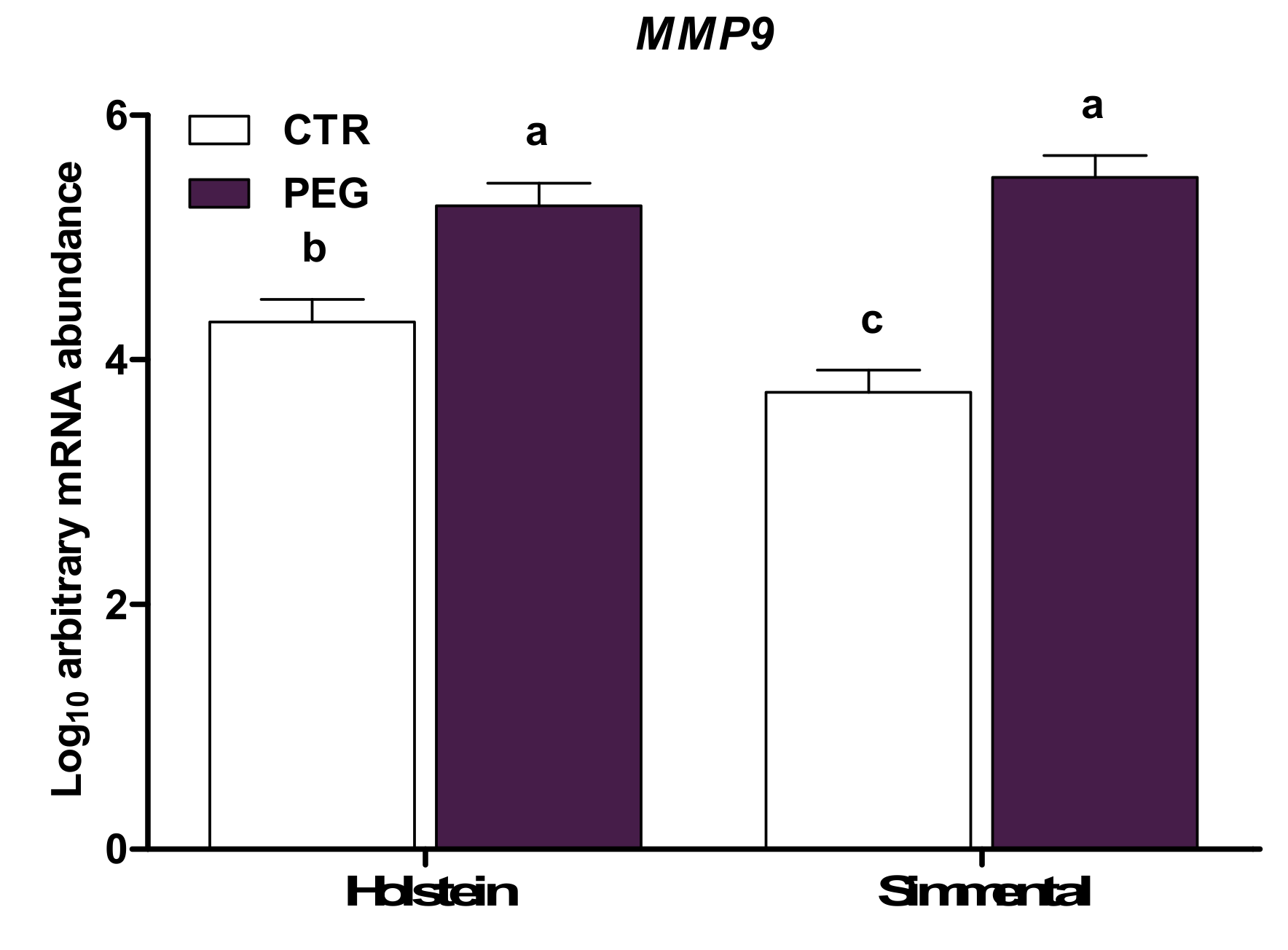
| MMP9*(4) | CTR | 4.31 | 3.73 | 3.74 | 4.31 | 0.27 | < 0.01 | 0.35 | 0.04 | 0.03 | 0.33 | 0.84 |
| PEG | 5.26 | 5.49 | 5.27 | 5.48 | ||||||||
| LTF*(4) | CTR | 1.57 | −0.07 | 0.26 | 1.23 | 0.59 | < 0.01 | 0.17 | 0.17 | 0.01 | 0.32 | 0.29 |
| PEG | 3.78 | 4.28 | 3.95 | 4.10 | ||||||||
| MPO* | CTR | 0.80 | 1.30 | 0.91 | 1.20 | 0.40 | < 0.01 | 0.13 | 0.47 | 0.74 | 0.73 | 0.29 |
| PEG | 2.28 | 2.61 | 2.39 | 2.50 | ||||||||
| LCN2* | CTR | 3.41 | 2.87 | 3.05 | 3.24 | 0.41 | < 0.01 | 0.92 | 0.39 | 0.50 | 0.84 | 0.54 |
| PEG | 4.78 | 5.38 | 4.93 | 5.23 | ||||||||
| IDO1(4) | CTR | 33.38 | 57.36 | 42.23 | 48.52 | 7.52 | 0.07 | < 0.01 | 0.40 | 0.14 | 0.04 | 0.38 |
| PEG | 31.65 | 39.98 | 43.32 | 28.31 | ||||||||
| p-Values | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed 1 | Parity 2 | Group X | |||||||||||
| Target | Group | H | S | P | M | SEM 3 | Group | Breed | Parity | Breed | Parity | Breed × Parity | |
| CASP1 | CTR | 27.84 | 27.79 | 28.79 | 26.83 | 2.72 | < 0.01 | 0.57 | 0.34 | 0.59 | 0.93 | 0.72 | |
| PEG | 35.09 | 33.02 | 34.87 | 33.25 | |||||||||
| TNFRSF1A* | CTR | 5.24 | 5.36 | 5.28 | 5.32 | 0.11 | < 0.01 | 0.83 | 0.61 | 0.16 | 0.35 | 0.84 | |
| PEG | 6.08 | 5.99 | 6.09 | 5.98 | |||||||||
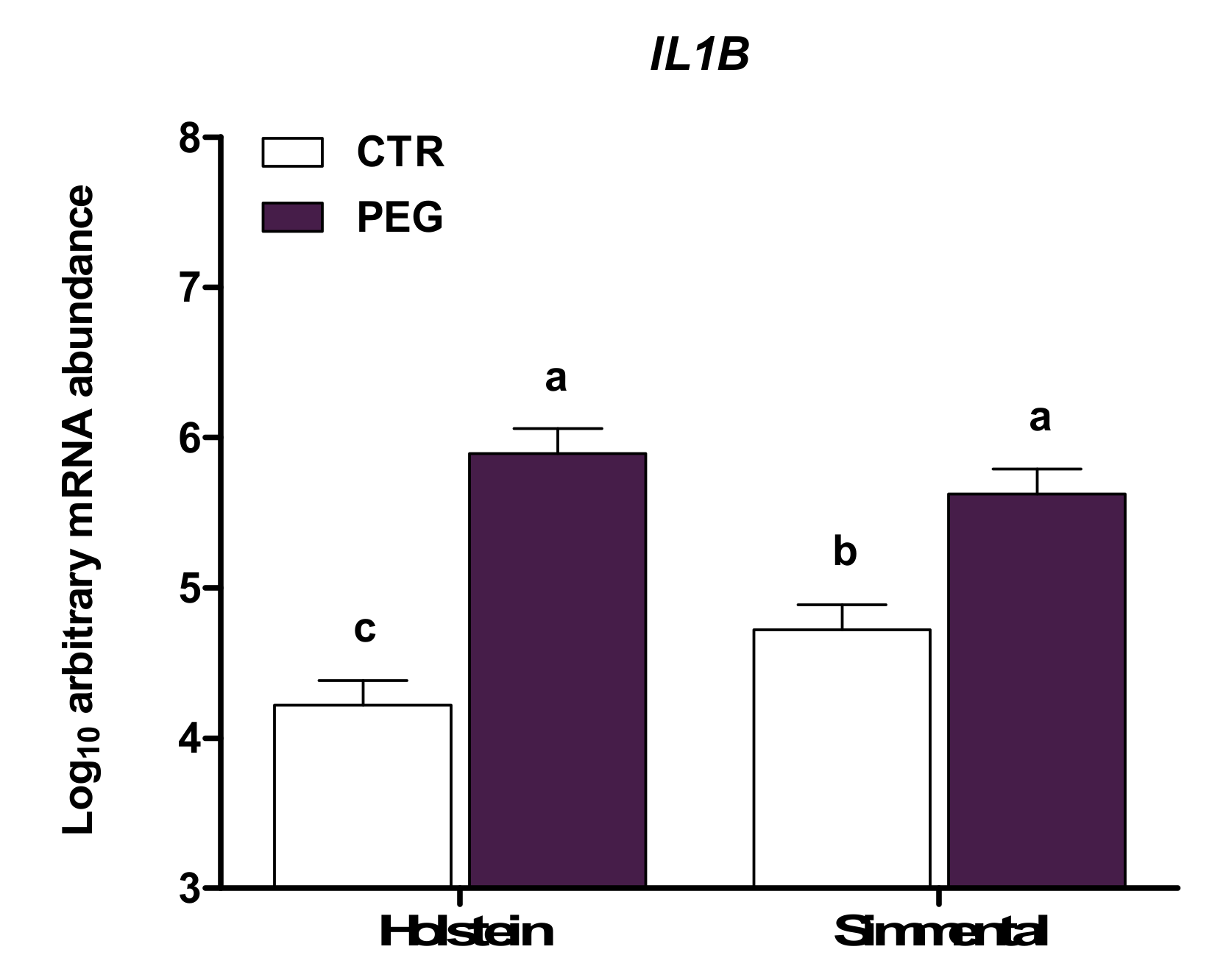
| IL1B*(4) | CTR | 4.22 | 4.72 | 4.66 | 4.28 | 0.24 | < 0.01 | 0.48 | 0.02 | 0.02 | 0.92 | 0.55 | |
| PEG | 5.89 | 5.63 | 5.97 | 5.55 | |||||||||
| IL1R* | CTR | 3.31 | 3.44 | 3.38 | 3.37 | 0.12 | < 0.01 | 0.24 | 0.83 | 0.71 | 0.95 | 0.75 | |
| PEG | 3.96 | 4.02 | 4.00 | 3.98 | |||||||||
| IL8* | CTR | 3.23 | 2.80 | 3.00 | 3.03 | 0.29 | 0.02 | 0.13 | 0.67 | 0.55 | 0.55 | 0.61 | |
| PEG | 2.62 | 2.43 | 2.63 | 2.42 | |||||||||
| IL18* | CTR | 2.48 | 2.43 | 2.58 | 2.33 | 0.45 | < 0.01 | 0.25 | 0.04 | 0.31 | 0.22 | 0.24 | |
| PEG | 4.93 | 4.26 | 5.11 | 4.08 | |||||||||
| IRAK1 | CTR | 41.60 | 40.30 | 42.11 | 39.79 | 3.32 | < 0.01 | 0.65 | 0.24 | 0.90 | 0.87 | 0.51 | |
| PEG | 58.82 | 58.08 | 59.99 | 56.91 | |||||||||
| TNF | CTR | 9.30 | 10.97 | 9.88 | 10.39 | 0.82 | 0.02 | < 0.01 | 0.93 | 0.55 | 0.41 | 0.39 | |
| PEG | 7.66 | 10.00 | 9.04 | 8.62 | |||||||||
| NLRP3* | CTR | 3.46 | 3.44 | 3.50 | 3.40 | 0.13 | < 0.01 | 0.20 | 0.30 | 0.28 | 0.98 | 0.51 | |
| PEG | 4.29 | 4.07 | 4.23 | 4.13 | |||||||||
| S100A8 | CTR | 1235.74 | 819.56 | 951.22 | 1104.09 | 386.69 | < 0.01 | 0.29 | 0.99 | 0.62 | 0.57 | 0.83 | |
| PEG | 2467.31 | 2314.12 | 2464.71 | 2316.71 | |||||||||
| RPL13A | CTR | 931.33 | 1089.30 | 1063.98 | 956.66 | 96.06 | < 0.01 | 0.05 | 0.13 | 0.22 | 0.91 | 0.93 | |
| PEG | 752.68 | 878.89 | 869.84 | 761.73 | |||||||||
| p-Values | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed 1 | Parity 2 | Group X | |||||||||||
| Target | Group | H | S | P | M | SEM 3 | Group | Breed | Parity | Breed | Parity | Breed × Parity | |
| SOD1 * | CTR | 4.08 | 4.11 | 4.16 | 4.03 | 0.11 | 0.29 | 0.55 | 0.10 | 0.35 | 0.94 | 0.34 | |
| PEG | 4.07 | 3.96 | 4.08 | 3.95 | |||||||||
| SOD2 * | CTR | 5.48 | 5.60 | 5.66 | 5.42 | 0.28 | < 0.01 | 0.49 | 0.07 | 0.17 | 0.64 | 0.95 | |
| PEG | 7.25 | 6.89 | 7.27 | 6.87 | |||||||||
| ALOX5 | CTR | 17.61 | 23.71 | 17.40 | 23.92 | 4.28 | < 0.01 | < 0.01 | 0.06 | 0.23 | 0.76 | 0.60 | |
| PEG | 22.30 | 35.48 | 26.52 | 31.26 | |||||||||
| ALOX15 | CTR | 23.83 | 34.06 | 27.75 | 30.14 | 5.40 | 0.02 | 0.09 | 0.68 | 0.30 | 0.82 | 0.77 | |
| PEG | 18.92 | 21.37 | 19.79 | 20.50 | |||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopreiato, V.; Palma, E.; Minuti, A.; Loor, J.J.; Lopreiato, M.; Trimboli, F.; Morittu, V.M.; Spina, A.A.; Britti, D.; Trevisi, E. Pegbovigrastim Treatment around Parturition Enhances Postpartum Immune Response Gene Network Expression of whole Blood Leukocytes in Holstein and Simmental Cows. Animals 2020, 10, 621. https://doi.org/10.3390/ani10040621
Lopreiato V, Palma E, Minuti A, Loor JJ, Lopreiato M, Trimboli F, Morittu VM, Spina AA, Britti D, Trevisi E. Pegbovigrastim Treatment around Parturition Enhances Postpartum Immune Response Gene Network Expression of whole Blood Leukocytes in Holstein and Simmental Cows. Animals. 2020; 10(4):621. https://doi.org/10.3390/ani10040621
Chicago/Turabian StyleLopreiato, Vincenzo, Ernesto Palma, Andrea Minuti, Juan J. Loor, Mariangela Lopreiato, Francesca Trimboli, Valeria Maria Morittu, Anna Antonella Spina, Domenico Britti, and Erminio Trevisi. 2020. "Pegbovigrastim Treatment around Parturition Enhances Postpartum Immune Response Gene Network Expression of whole Blood Leukocytes in Holstein and Simmental Cows" Animals 10, no. 4: 621. https://doi.org/10.3390/ani10040621