1. Introduction
Soybean meal (SBM), a byproduct of oil extraction from soy seed, is the most commonly used plant protein source in the poultry and swine feed industries. However, a variety of anti-nutritional factors (ANFs) present in SBM, such as antigenic proteins, trypsin inhibitors, and oligosaccharides, interfere with digestion and absorption and have negative effects on animal health [
1]. Accumulating evidence has demonstrated that microbial fermentation improves the nutritional quality of SBM by reducing the content of ANFs and increasing nutrient bioavailability [
2,
3,
4]. Fermented SBM (FSBM) is produced by the addition of fungal or bacterial microorganisms, including
Lactobacillus plantarum,
Bacillus subtilis,
Aspergillus oryzae, and
Neurospora crassa [
5,
6,
7]. Zhang et al. [
6] showed that 92.36% of glycinin and 88.44% of β-conglycinin was eliminated from SBM, and the amount of trichloroacetic acid-soluble protein (TCA-SP) was increased 4.6-fold after the solid-state fermentation of SBM with
B. subtilis BS12 for 24 h. A previous study reported that the solid-state fermentation of SBM with
Bacillus amyloliquefaciens for 48 h degraded soybean macro-molecular proteins to less than 25 kDa and completely decomposed raffinose and stachyose in SBM [
2].
Several studies have demonstrated that partial or total replacement of SBM with FSBM improves the growth performance, digestive enzyme activity, and gut morphology of broilers [
8,
9,
10]. Feeding broilers FSBM produced by mixed strains, including
L. plantarum,
Lactobacillus. acidophilus,
B. subtilis, and
A. oryzae, significantly increased body weight gain and decreased feed conversion ratio (FCR) between d 11 and 24 and d 25 and 42, improved apparent ileal digestibility of crude protein and gross energy, and increased villus height (VH) and villus height to crypt depth (VH: CD) ratios in the duodenum and jejunum [
9]. In another study, Jazi et al. [
10] reported that dietary FSBM mitigated the growth performance suppression and decrease of VH and VH: CD in the duodenum and jejunum of young broiler chickens challenged by
Salmonella typhimurium from d 1 to 24. Interestingly, Kim et al. [
11] found that feeding broilers 3% FSBM fermented by
Bacillus alone or in combination with a yeast byproduct in the first 7 days post-hatching significantly increased average daily gain (ADG) and decreased FCR throughout the entire growth phase. In contrast, a diet containing 10% FSBM with complete elimination of glycinin and β-conglycinin did not improve the growth performance of broilers but did increase average feed intake (ADFI) during d 1 to 21 [
4]. Therefore, further study of the effects of FSBM produced by different kinds of microorganisms, fermentation times, and processing methods on the growth performance of broilers is needed.
The intestinal microbiota of animals plays an important role in maintaining intestinal homeostasis and animal health by modulating nutrient digestion, protecting against enteric pathogens, enhancing intestinal immunity, and performing other physiological functions [
12]. It was reported that nutrient absorption, feed digestibility, and energy harvest are strongly linked to the gut microbiota and, therefore, animal productivity is influenced by microbial composition and diversity. Furthermore, the gut microbial composition and diversity can be modulated by diet, including feed components and feed additives [
13]. FSBM fermented by microorganisms not only increases the nutritional quality of SBM but also provides prebiotics, probiotics, and their metabolites to animals, thereby exerting growth-promoting effects [
14]. A previous study in weaning piglets demonstrated that dietary supplementation with FSBM modulated the microbial composition of the colon and feces and increased the abundance of the potentially beneficial bacteria
Lachnospira and
Lactobacillus [
15]. Xie et al. [
16] also found that feeding piglets 15% fresh FSBM altered intestinal bacterial community structure and increased the relative abundance of butyrate-producing bacteria. A recent study using a traditional culture-based approach reported that FSBM supplementation of broiler diets increased the number of lactic acid bacteria, decreased the number of
Coli-form bacteria and
Clostridium perfringens in the ileum and cecum [
9]. However, the broiler gut microbiota includes hundreds of bacterial species, and it is difficult to cultivate and study the composition, diversity, and structure of intestinal microbiota using traditional culture-based approaches [
17]. In recent years, 16s rDNA gene amplicon deep sequencing has been widely applied in a range of studies to investigate microbial composition and diversity and their relationship to the growth performance [
17,
18] and immunity of animals [
15,
19,
20].
Currently, to the best of our knowledge, there is very little information on the effects of FSBM on cecal microbial composition and diversity in broilers. Therefore, the objective of this study was to investigate the effects of FSBM supplementation on the growth performance, serum immunity, and microflora composition of the cecum of broilers using Quantitative Insights Into Microbial Ecology (QIIME2) and high-throughput sequencing.
4. Discussion
SBM is the most commonly used plant protein source in poultry and swine production. However, various ANFs, such as antigenic protein and trypsin inhibitors, limit its wide application in animal feed [
26]. A growing number of studies indicate that microbial fermentation is an economically and widely available method to solve the issues of SBM [
2,
21,
27]. In pig production, extensive evidence has shown that FSBM or fermented feed improves growth performance, nutrient digestibility, and immune function [
16,
19,
26,
28]. However, to date, information on the effects of FSBM on the growth performance of broilers is limited and inconsistent. In the present study, the results demonstrate that substituting 25% of the SBM in the diets with FSBM (8.18% FSBM addition) significantly increased the ADG and FCR of broilers during both the growth (d 22–36) and whole (d 1–36) phases, compared with those in the control group, but did not affect ADG, ADFI, and FCR during the starter (d 1–22 ) phase. Similarly, Chachaj et al. [
29] reported that replacing SBM with 6% FSBM significantly increased bodyweight gain at d 32 and d 40, but did not affect feed intake or FCR. A study of bacteria and fungi-mixed fermented SBM found that supplementing the diets of broiler chickens with 4.5–6.0% wet or dry FSBM had better effects on growth performance than 5% SBM [
30]. Feng et al. [
8] reported that replacing all SBM in the diets of broiler with FSBM fermented by
A. oryzae 3.042 significantly improved ADG and ADFI. The enhanced growth performance of broilers fed FSBM in these studies was associated with a reduction in ANFs from SBM, improved intestinal morphology, and increased digestive enzyme activity. However, Guo et al. [
31] found that supplementation with 7.5% FSBM, fermented with
B. subtilis,
L. spp., and yeasts, did not affect the growth performance of broilers over the entire rearing period. Additionally, feeding broilers 10% FSBM, produced by
B.subtilis combined with protease, increased higher ADFI but did not improve ADG throughout the entire feeding period [
4]. These discrepancies between our observations and previous findings might be attributable to the different types of microorganisms used for fermentation, the various methods used to process FSBM, or the different amounts of FSBM added to the diets. Interestingly, in our study, the ADG and ADFI of broilers in the TC group (50% of SBM in the basal diet replaced with FSBM) did not significantly improve compared with those of the CC group during the whole phase. This result probably relates to the increase in free amino acids that occurs in SBM after fermentation. Previous studies have demonstrated that the absorption of amino acids in the form of free amino acids is much slower and requires more energy than the absorption of oligopeptides [
32,
33]. Therefore, higher levels of free amino acid in the diets may have affected the growth performance of broilers.
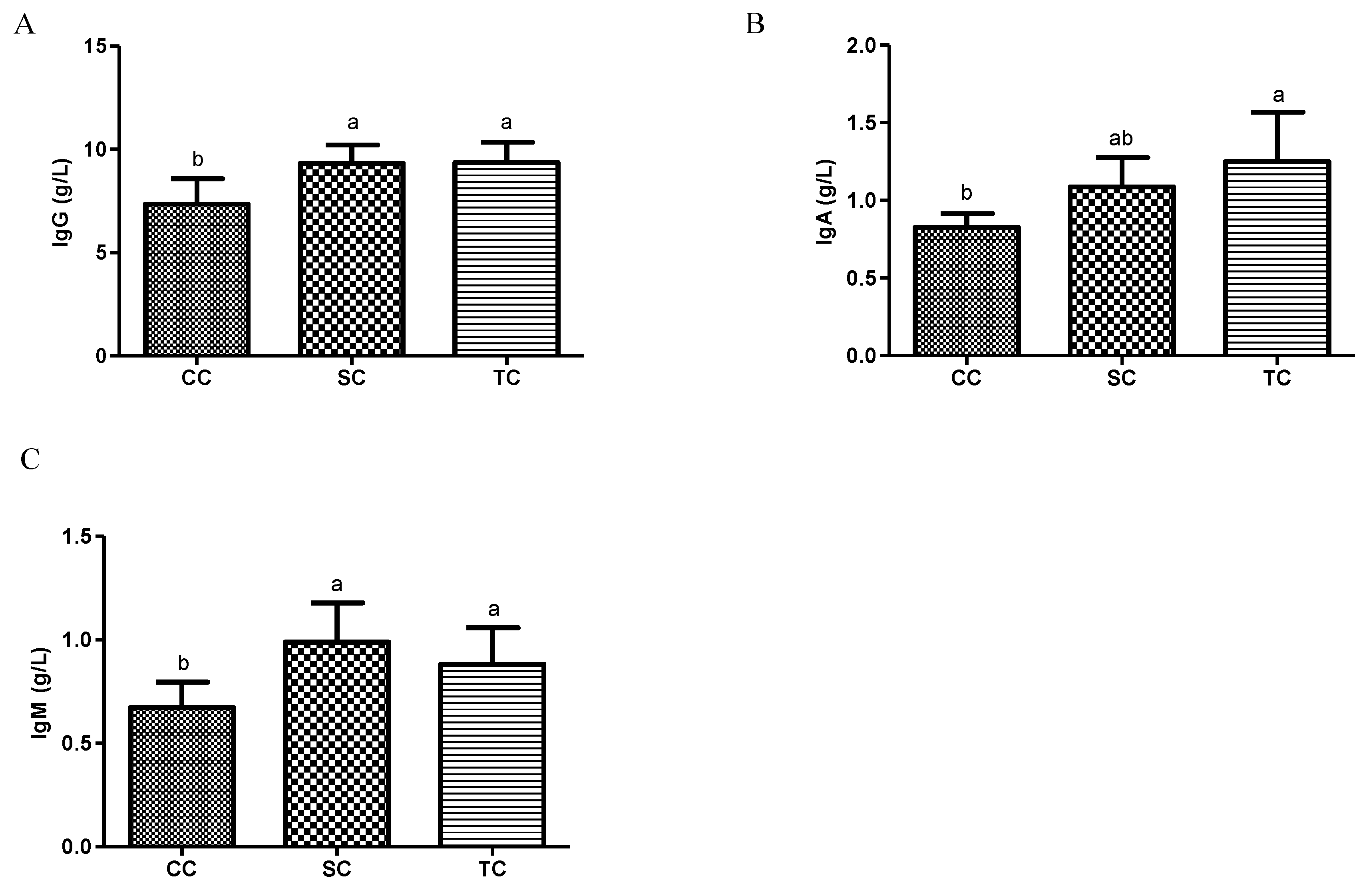
Serum immunoglobulin concentrations are commonly considered to be vital indicators of the humoral immunity of animals, as immunoglobulins are immune-active molecules that play important roles in defending the host against pathogenic viruses and microorganisms [
19,
34]. IgG, IgM, and IgA are the three major immunoglobulin classes in avian species. In the present study, concentrations of IgG, IgM, and IgA were significantly higher in the two dietary treatment groups, compared with the CC group, suggesting that FSBM enhanced the immune function of broilers. Our results are in agreement with the findings of Zhu et al. [
28], who noted a significant increase in serum concentrations of IgG and IgM in weaning piglets fed FSBM. Previous research showed that piglet immunity significantly decreased when β-conglycinin was not adequately deactivated during fermentation [
35,
36]. Our research revealed an increase in the immune function of broilers in the SC and TC groups that was directly related to the lower levels of glycinin and β-conglycinin in FSBM compared with SBM. Several previous studies observed that the formation of small sized peptides during fermentation were associated with increased immunoglobulin levels in broilers [
37,
38].
The gut microbiota plays an important role in maintaining the normal physiological structure and function of the intestinal tract. Previous evidence has revealed a close correlation between the growth performance and gut microbiota of different species of animals [
17,
18,
19]. The gut microbial composition is extensively influenced by many factors, such as diet, age, feeding patterns, host genotype, pathogen infections, and feed additives [
12]. A previous study indicated that diet has a significant effect (estimated to be 57%, compared with 12% for genetic factors) on the intestinal microbial community structure [
39]. Intestinal microbial composition can quickly be altered by changes in dietary components [
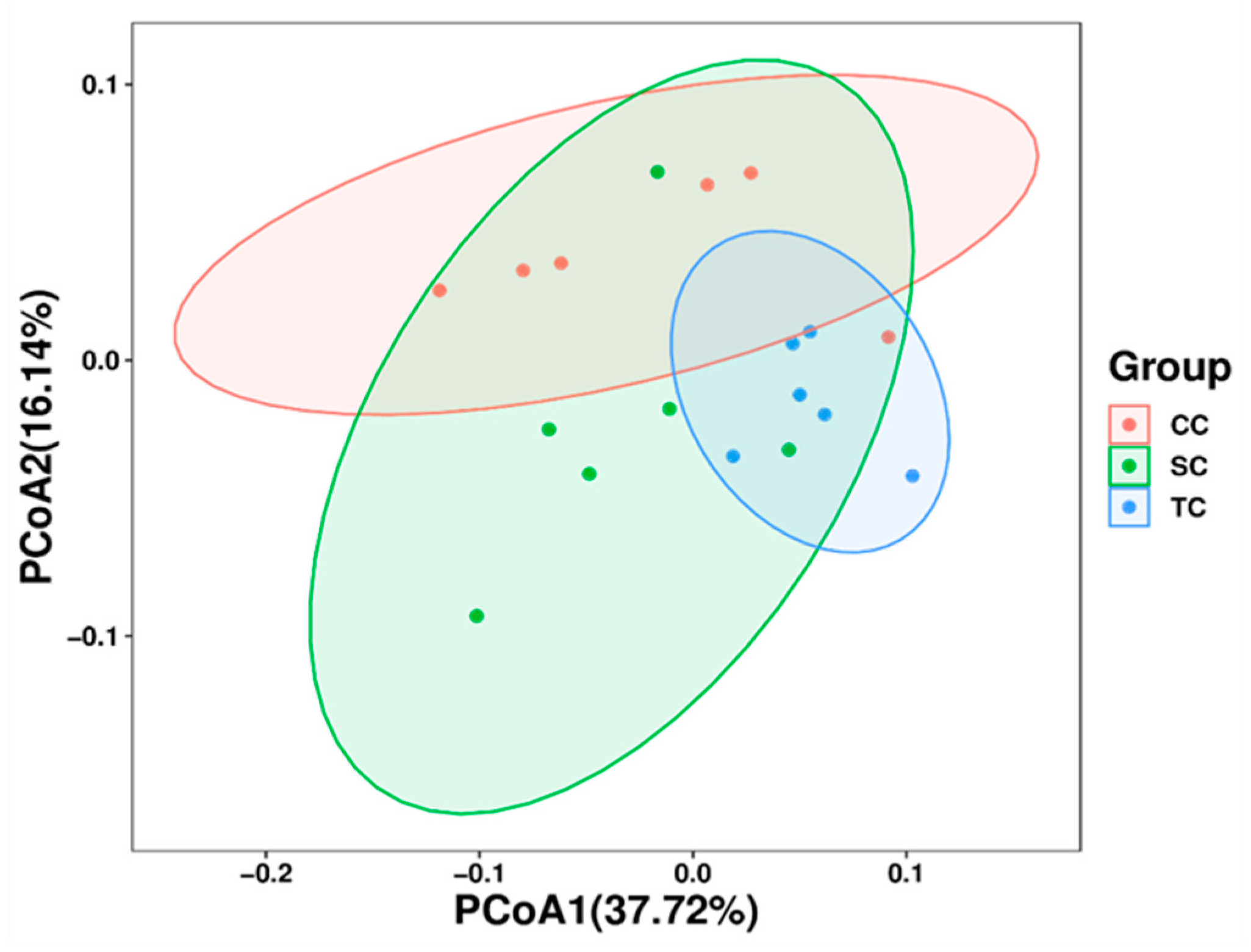
40]. In the present study, OS and Chao1 richness estimators increased significantly in broilers fed FSBM, indicating greater species richness in the cecal digesta. It has been reported that Chao1 and Shannon indices of alpha diversity were highest in the colon of pigs fed diets containing FSBM [
15]. Previous research demonstrated that a highly diverse gut microbiota is more stable and improves the health of animals compared with less diverse microbiota [
17,
39], as harboring a broad range of bacteria in the intestinal tract enables the flora to better deal with environmental perturbations [
41].
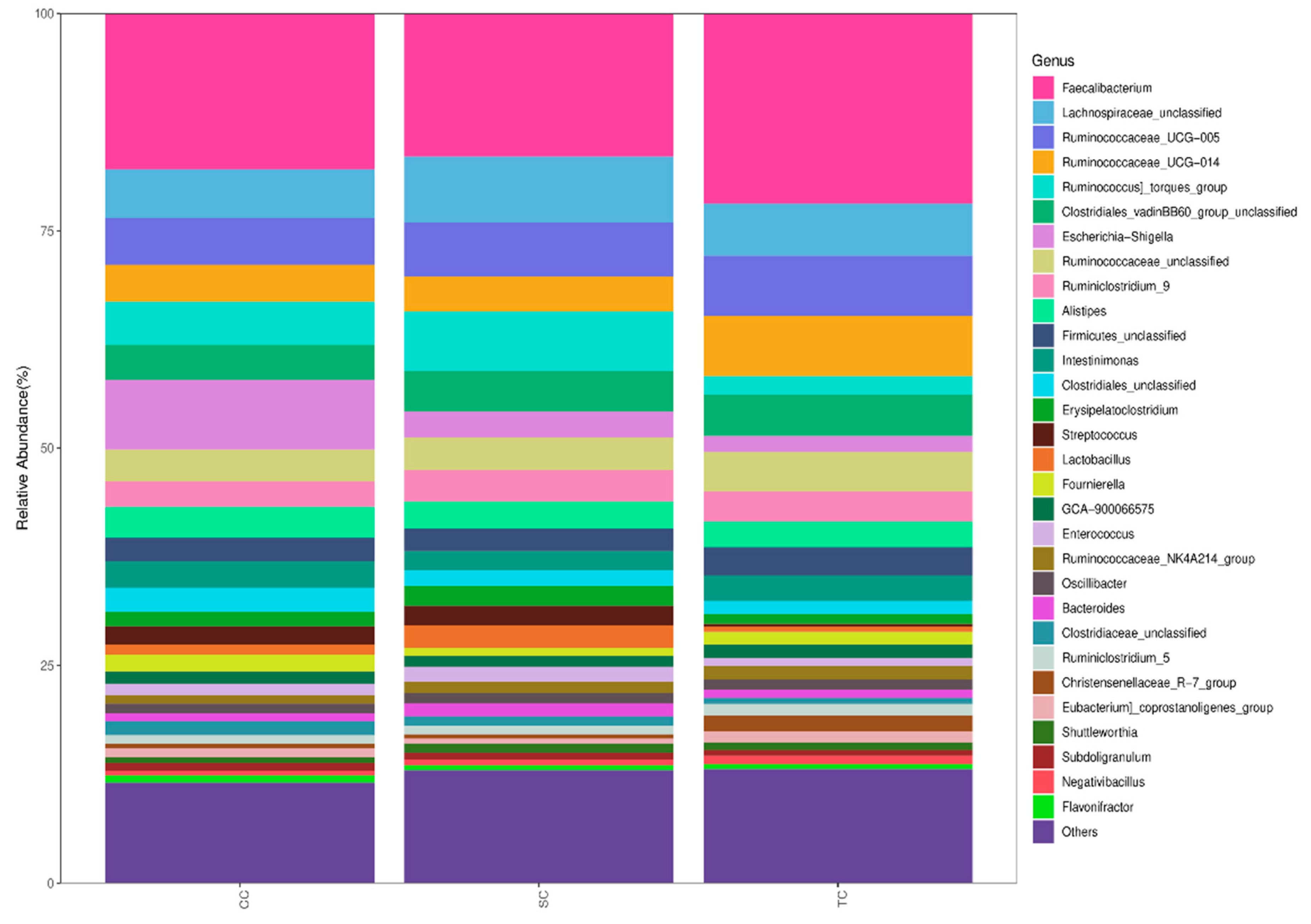
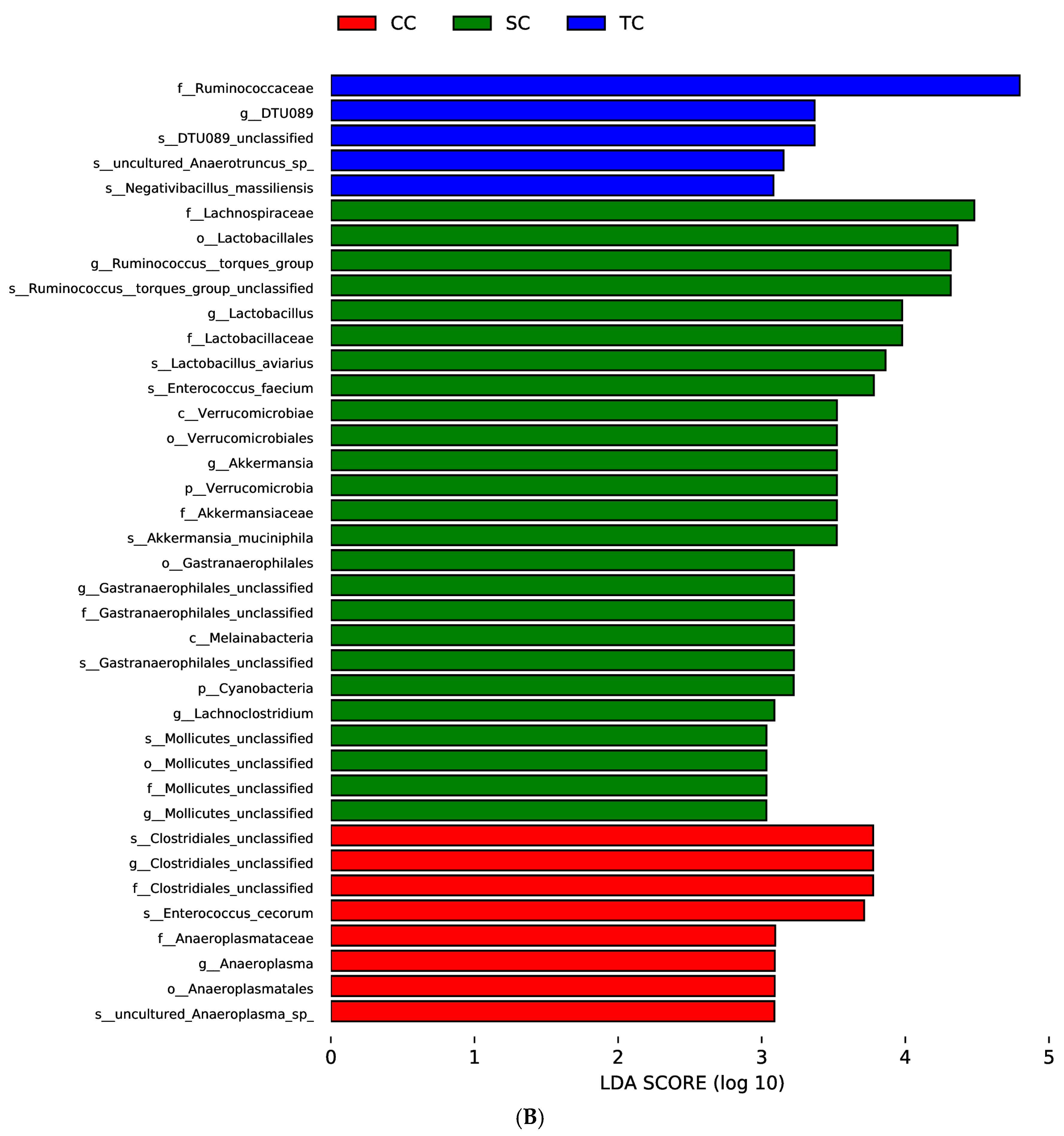
Regardless of dietary treatment,
Firmicutes,
Proteobacteria,
Tenericutes, and
Bacteroidetes were the four pre-dominant bacterial phyla in the cecal microbiota, which is similar to the results of a previous study of broilers [
42]. However, replacing SBM in the diets of broilers with FSBM modulated cecal microbiota composition at the phylum level. The relative abundance of
Firmicutes was significantly increased in the SC and TC groups.
Firmicutes are associated with the decomposition of polysaccharide and the utilization of energy in the gut owing to their genes encoding non-starch polysaccharides degrading enzymes. Moreover, the abundance of
Firmicutes and the ratio of
Firmicutes to
Bacteroides are often positively correlated with the growth performance of animals [
13,
43]. Therefore, the increase in
Firmicutes observed in the dietary treatment groups could at least partially explain the improvement in ADG observed in broilers fed FSBM. Intestinal bacteria belonging to
Proteobacteria include a wide variety of pathogens such as
Brucella spp.,
Rickettsia spp., and
Neisseria spp [
44]. In the present study, the relative abundance of
Proteobacteria was much lower in the two dietary treatment groups than in the CC group, which is consistent with previous research that showed that the addition of FSBM to diets decreased the number of harmful bacteria belonging to the phylum
Proteobacteria, such as
Escherichia coli [
9]. It has been reported that the phylum
Proteobacteria has a low abundance in the intestine of healthy humans [
44]. However, an increased abundance of
Proteobacteria has been observed in humans with enteric infections, colorectal cancer, and metabolic syndrome, and may be a microbial signature of dysbiosis in the gut microbiota [
45]. In young pigs, undigested dietary proteins in the gastrointestinal tract enter the hindgut and become a substrate of fermentation by some genera belonging to
Proteobacteria, such as
E. coli,
Klebsiella spp., and
Campylobacter spp [
46]. Conversely, these undigested dietary proteins promote the reproduction of these harmful bacteria [
47]. Therefore, we speculate that the lower abundance of the phylum
Proteobacteria that we observed may be related to the improved digestibility of the crude protein in FSBM. On the other hand, the Lactobacillus in FSBM reduces gut pH by the production of organic acids and prevents the colonization of enteropathogens through competitive exclusion, antagonistic activities and bacteriocin production, which is also contributable to decrease the abundance of the phylum
proteobacteria [
48]. Interestingly, we observed that the phylum
Verrucomicrobia was only present in the two dietary treatment groups.
Verrucomicrobia is phylogenetically closely related to
Planctomycetes and
Chlamydiae, which represent 1–3% of the total microbiota in humans and are negatively correlated with inflammatory bowel disease, Crohn’s disease, and non-obese diabetes [
49]. Therefore, the modulation of
Firmicutes,
Proteobacteria, and
Verrucomicrobia may be beneficial to maintain the normal function of the intestine and enhance the growth performance of broilers.
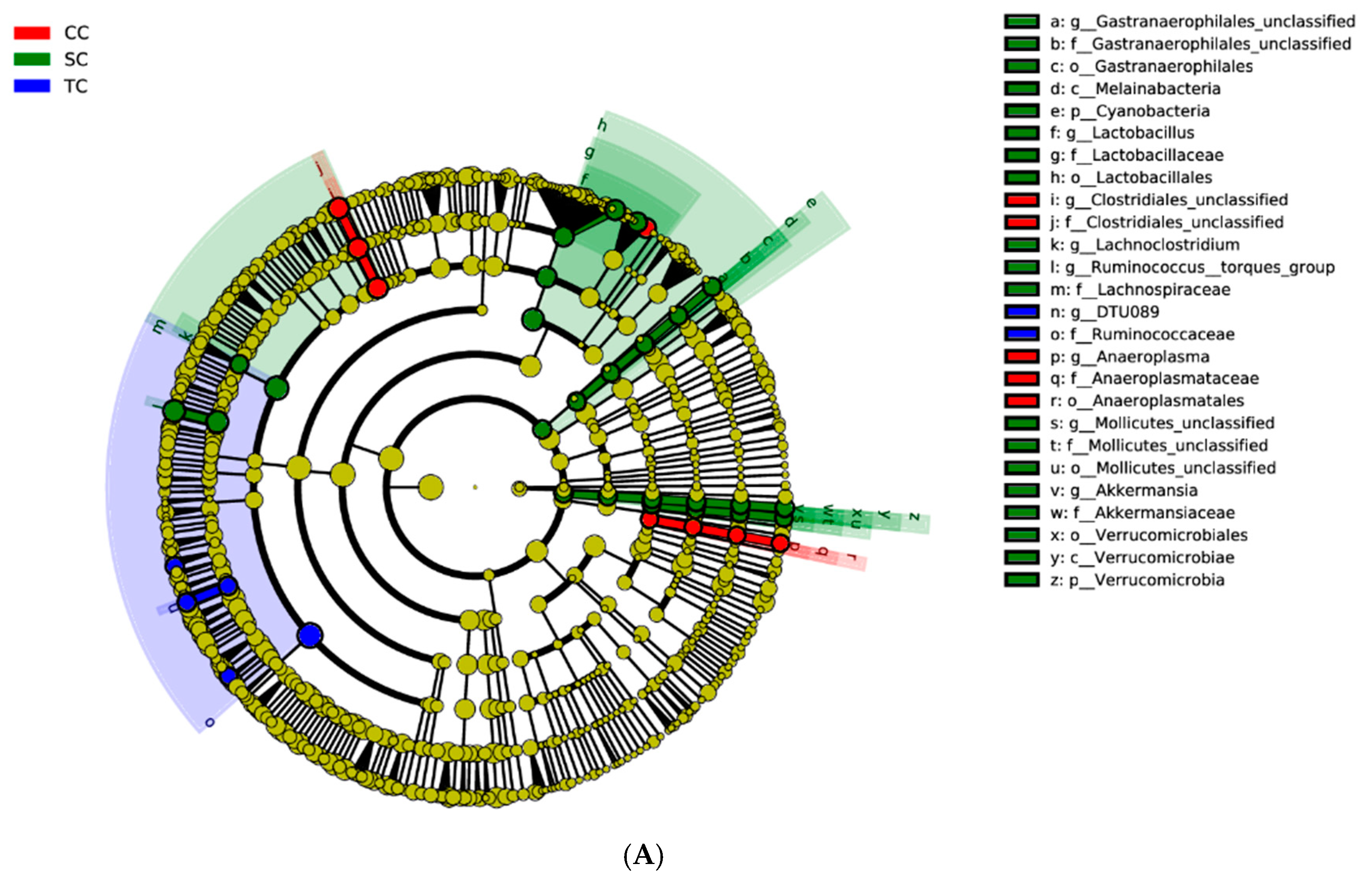
At the genus level, our results showed that the abundance of
Lachnospiraceae,
Lactobacillus, and
Lachnoclostridium increased significantly in broilers fed diets supplemented with FSBM. Studies have shown that
Lachnospiraceae constitutes one of the major taxonomic groups of the human gut microbiota and is correlated with human health [
50,
51]. The lower abundance of
Lachnospiraceae was previously reported in multiple sclerosis and ulcerative colitis patients [
51]. In animals, herbivores have a higher abundance of
Lachnospiraceae than carnivores [
50]. All members of
Lachnospiraceae are anaerobic, fermentative, and chemoorganotrophic and can degrade non-starch polysaccharides and produce acetic acid and butyrate [
51]. Butyrate is the primary energy source for host epithelial cell growth, enhances epithelial barrier integrity, and inhibits inflammatory responses [
41,
52]. In addition,
Lachnospiraceae is associated with increased FCR and bodyweight gain in broilers [
53], which was confirmed in our study. The higher relative abundance of the genus
Lachnospiraceae may be a result of decomposition of non-starch polysaccharides in SBM during fermentation, as the products of degraded non-starch polysaccharides can be more easily fermented by members of the genus
Lachnospiraceae. The genus
Lachnoclostridium—butyric-acid-producing bacteria that have been implicated in the alleviation of gut inflammation—was more abundant in the two dietary treatment groups [
54].
Lactobacillus is known to have a beneficial effect on the gastrointestinal tract and the growth of broilers and is commonly used as probiotics in animal production [
20,
42].
Lactobacillus promotes the growth performance of animals by protecting the gut from pathogens and improving nutrient and energy extraction by the host [
55]. In the present study, the relative abundance of
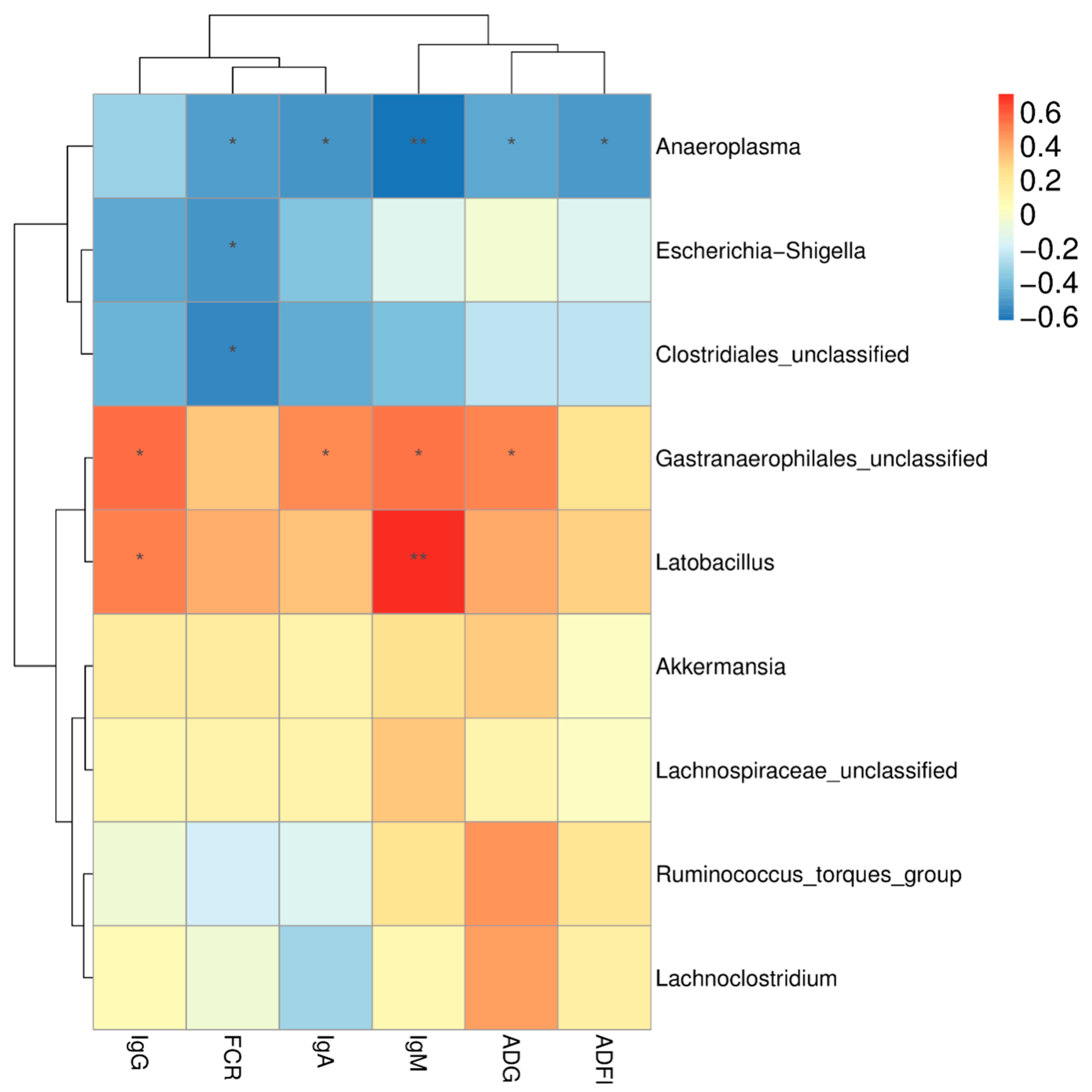
Lactobacillus was higher in the two dietary treatment groups than in the control group and was positively correlated with serum IgM and IgG levels. Wang et al. [
20] also found that feeding weaning piglets
L. plantarum PFM 105 increased the serum IgM levels. The genus
Gastranaerophilales was more abundant in the SC group and was positively correlated with ADG and serum immunity. However, little is known about this microorganism. Ma et al. [
56] reported that
Gastranaerophilales is capable of fermenting a range of sugars (e.g., glucose, starch, and hemicellulose) to produce butyrate in the gut of herbivores. Borsanelli et al. [
57] found that the abundance of
Gastranaerophilales was higher in the mouths of healthy cattle and lower in the mouths of cattle with bovine periodontitis. Notably,
Akkermansia, the only genus in the phylum
Verrucomicrobiae, was only present in the two dietary treatment groups.
Akkermansia has been reported to be a highly specialized bacterium capable of using mucin as its sole carbon and nitrogen source, and of stimulating mucin expression [
58]. An increase in
Akkermansia has been demonstrated to be protective against from inflammatory bowel diseases, metabolic diseases, and neurological disorders [
58,
59]. Yan et al. [
55] observed that
Akkermansia was more abundant in the cecum of a higher feed efficiency group, and our study similarly demonstrated that the relative abundance of
Akkermansia was weakly positively correlated with the ADG of broilers. In addition,
Clostridiales was found to be less abundant in the two dietary treatment groups and strongly negatively correlated with FCR. Another effect of dietary treatment was a significant reduction in
Escherichia-Shigella, and the decrease in
Escherichia-
Shigella was the major contributor to the lower abundance of the phylum
Proteobacteria compared with control group. The genus
Escherichia–Shigella includes opportunistic pathogenic bacteria. Previous research has demonstrated that
Escherichia–Shigella impairs intestinal structure and induces various pro-inflammatory pathways, such as the secretion of virulence factors, resulting in an increased risk of infection and diarrhea in the host [
60,
61]. The genus
Anaeroplasma is positively correlated with more severe clinical scores in the animal model of multiple sclerosis, but
Lactobacillus reuteri treatment reduced its abundance and improved the immunity of animals [
62]. Du et al. [
63] found that supplementation with
B. amyloliquefaciens in the diet decreased the abundance of
Anaeroplasma in growth-retarded beef calves. Our research shows a reduction in the abundance of
Anaeroplasma in the two dietary treatment groups, suggesting that FSBM supplementation inhibits the reproduction of the potentially harmful bacteria
Anaeroplasma. The explanation for this requires further investigation in future research.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}