
The Relation between Migratory Activity of Pipistrellus Bats at Sea and Weather Conditions Offers Possibilities to Reduce Offshore Wind Farm Effects


Abstract
:Simple Summary
Abstract
1. Introduction
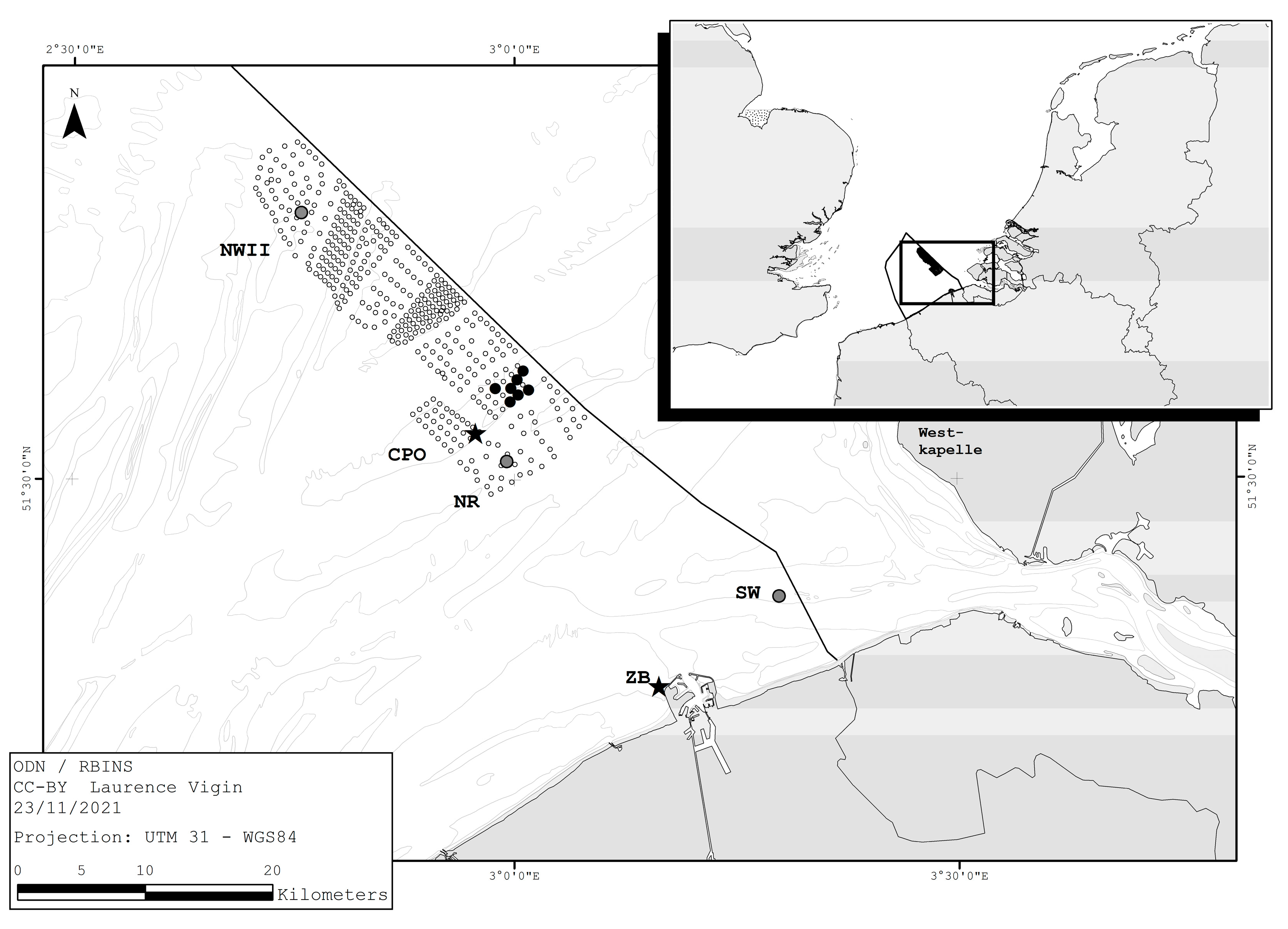
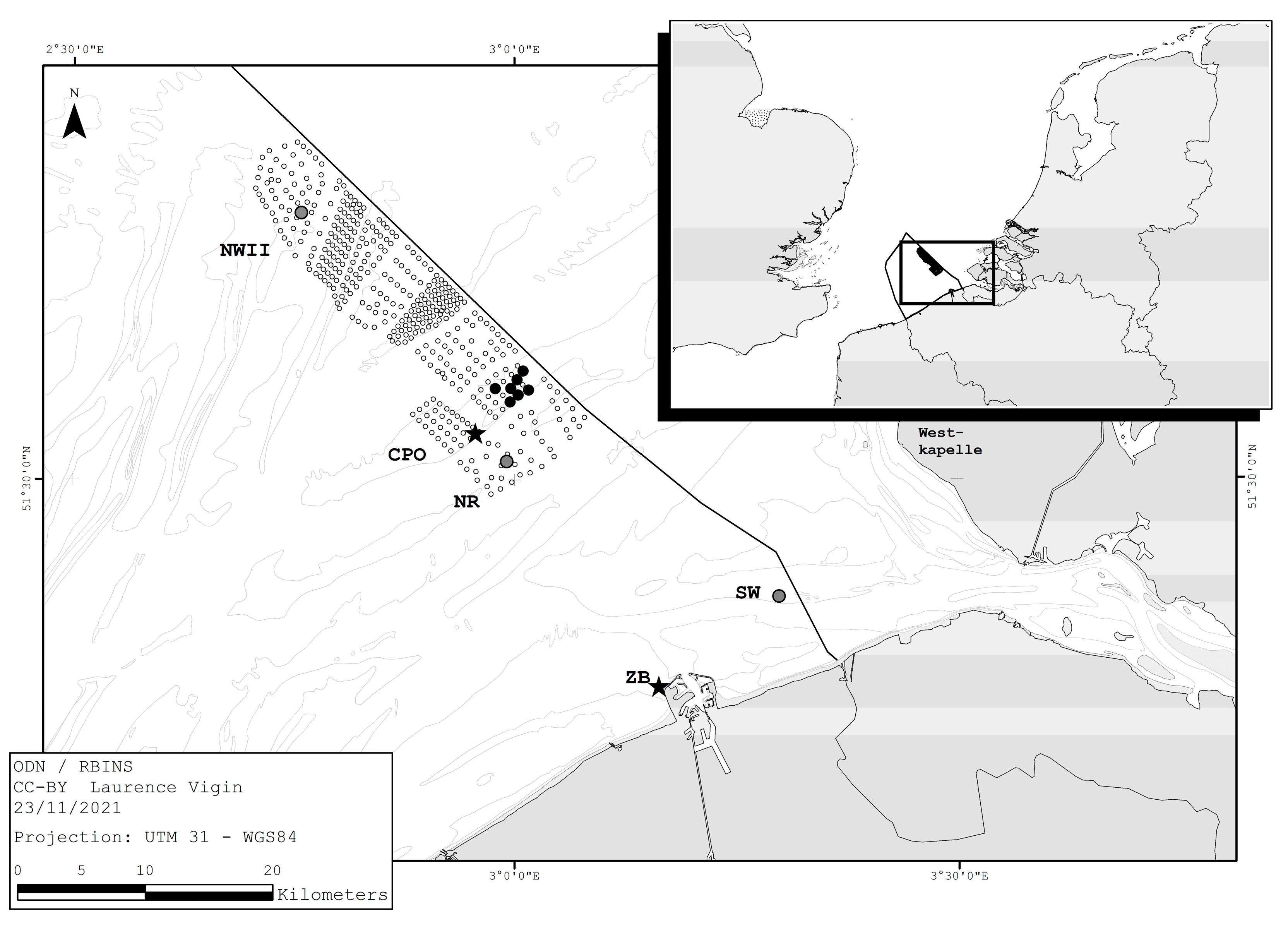
2. Materials and Methods
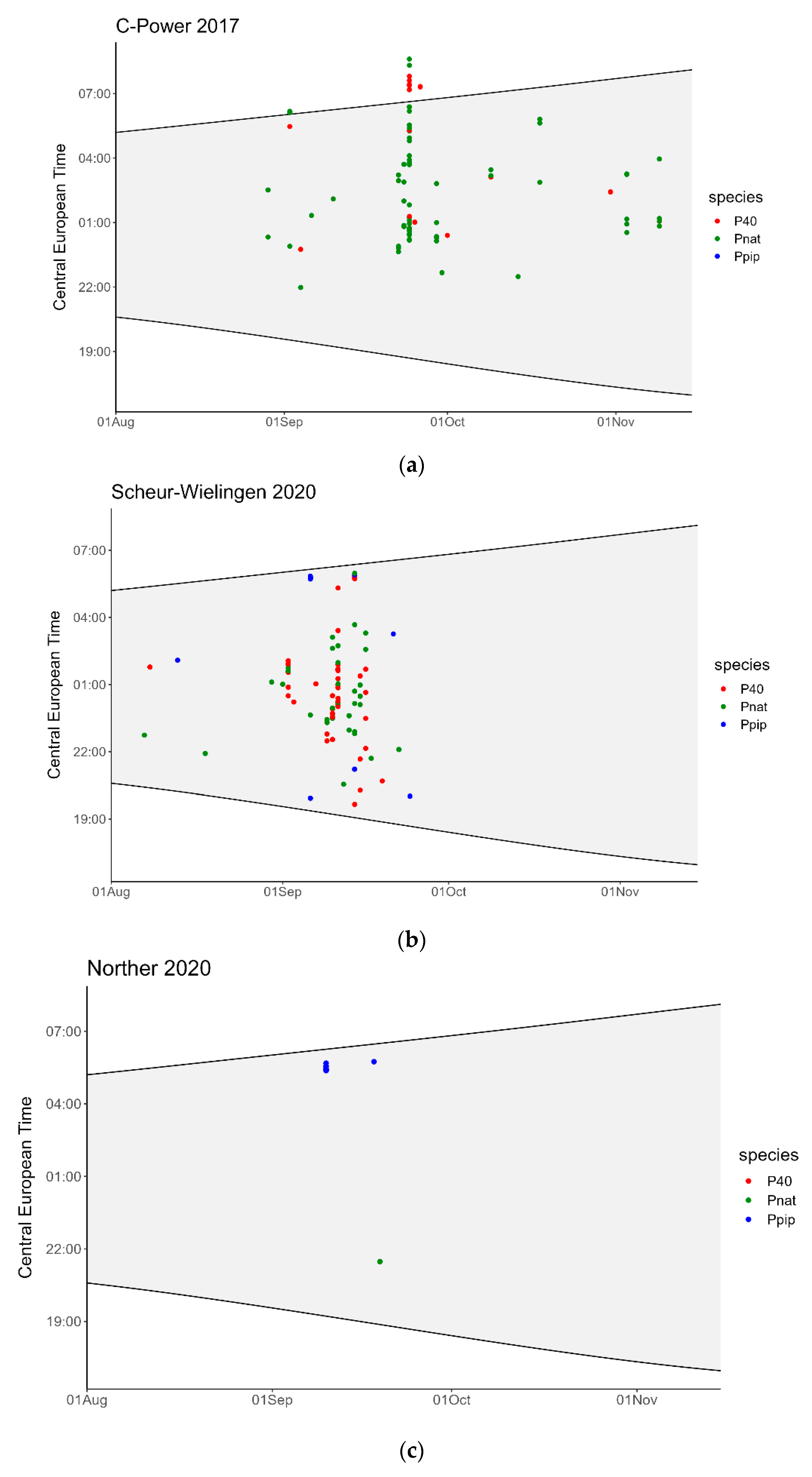
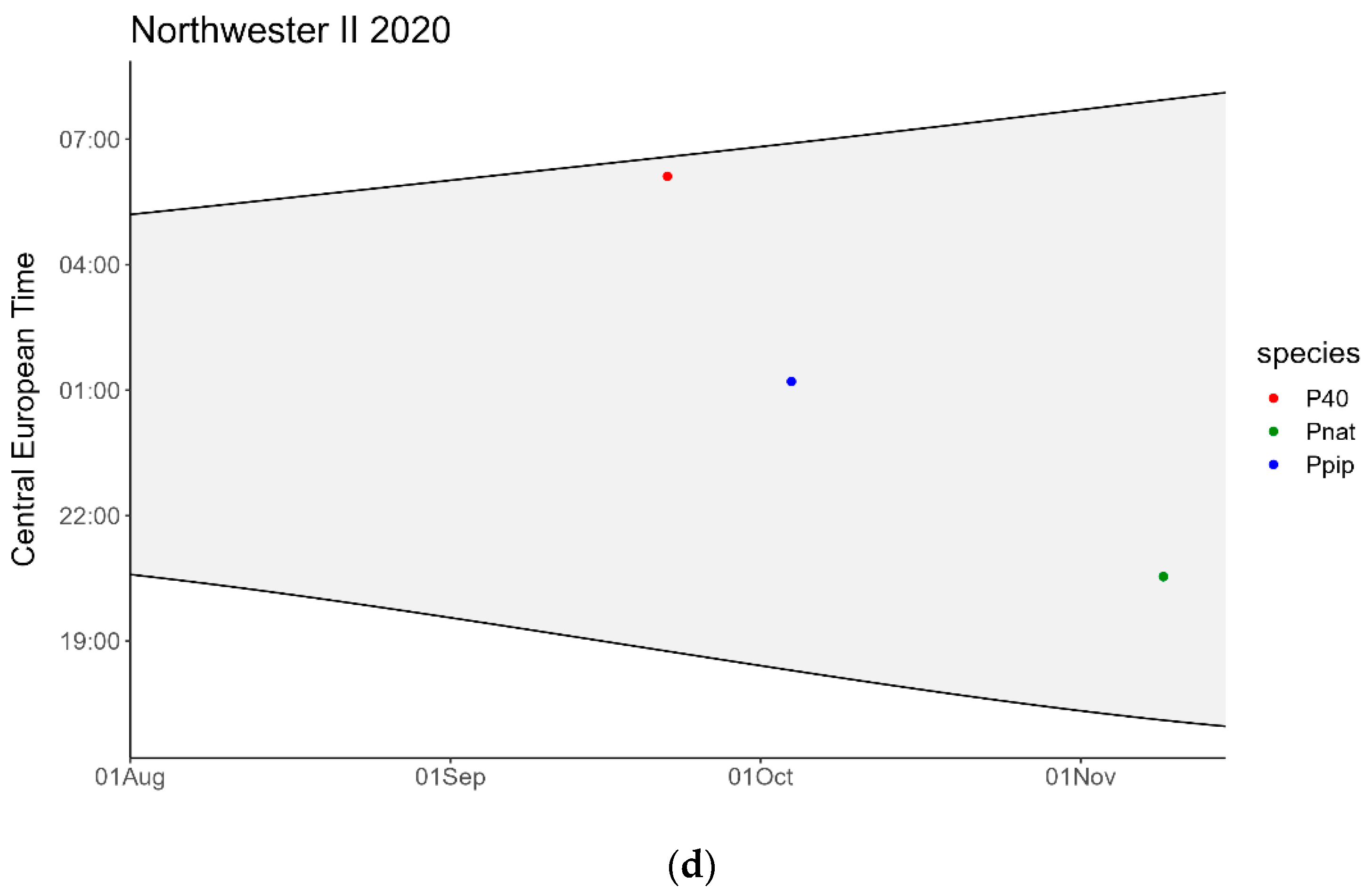
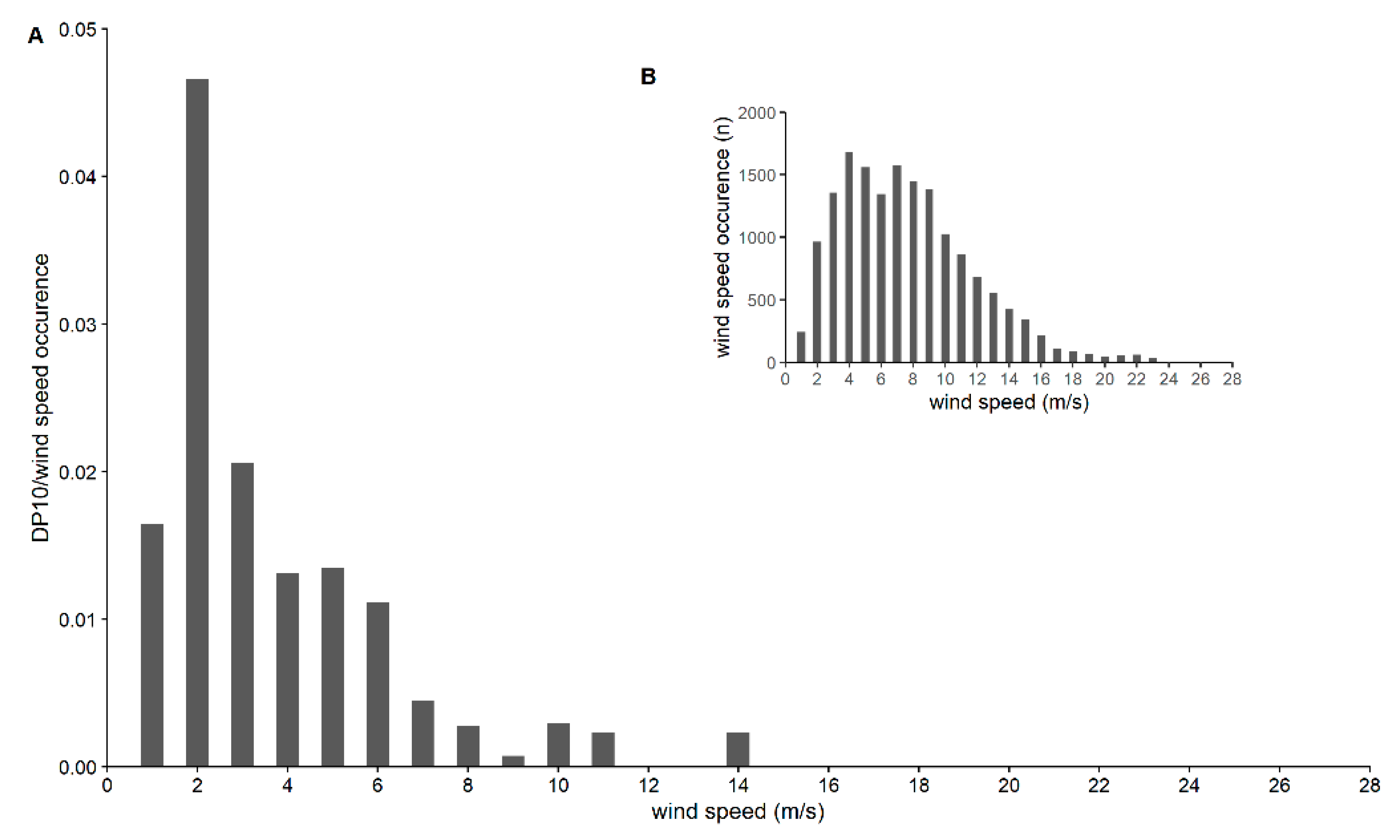
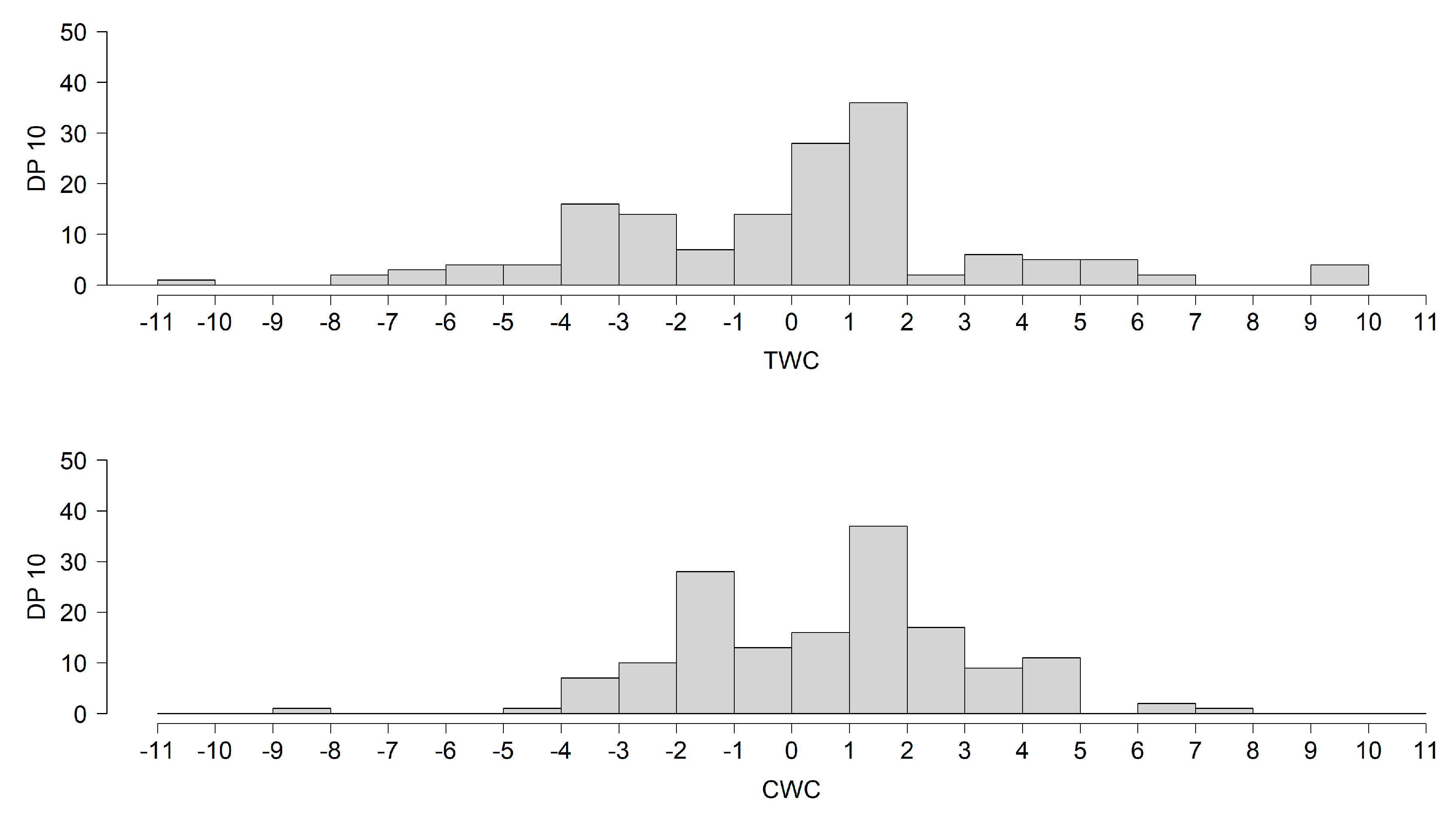
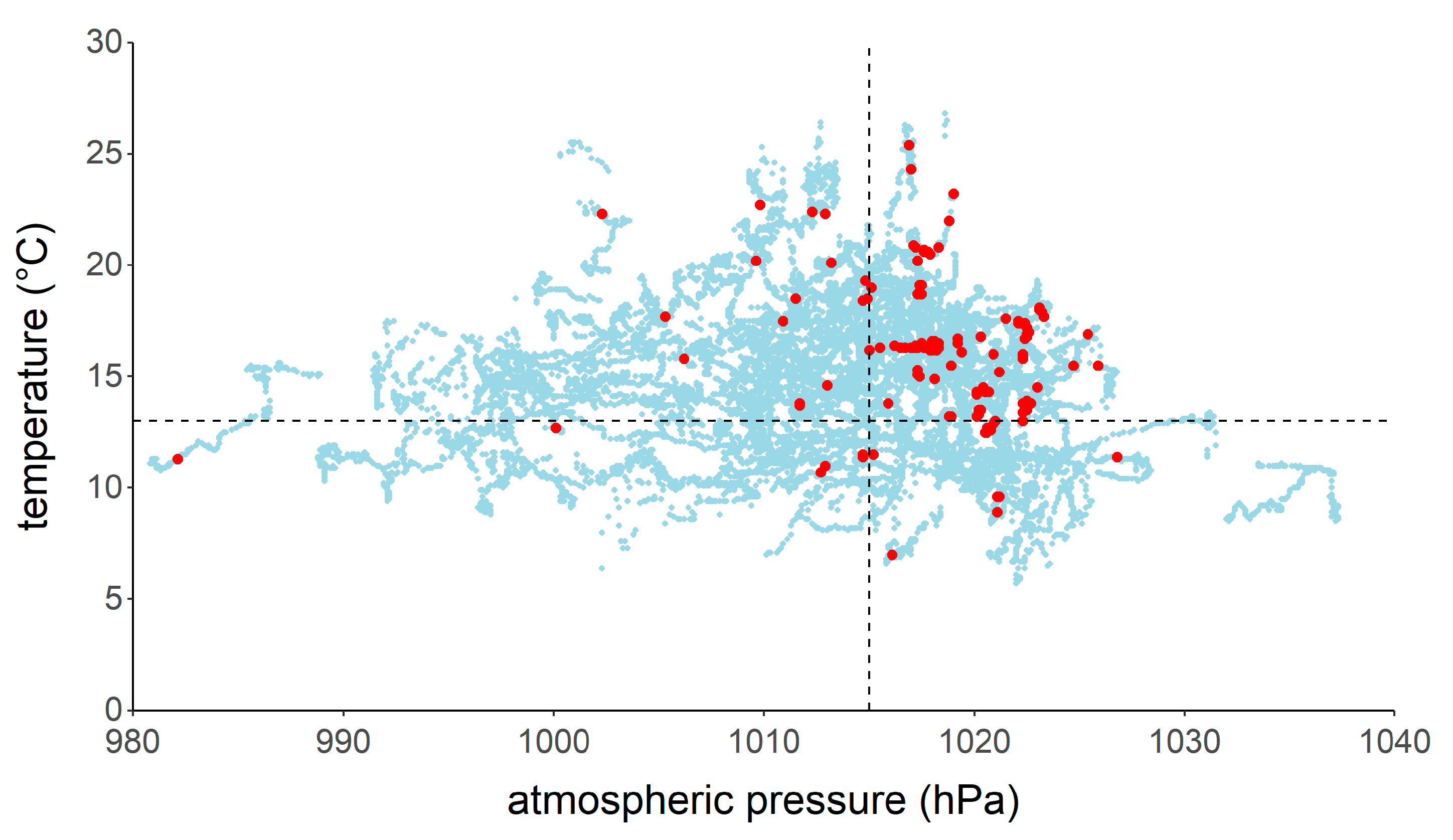
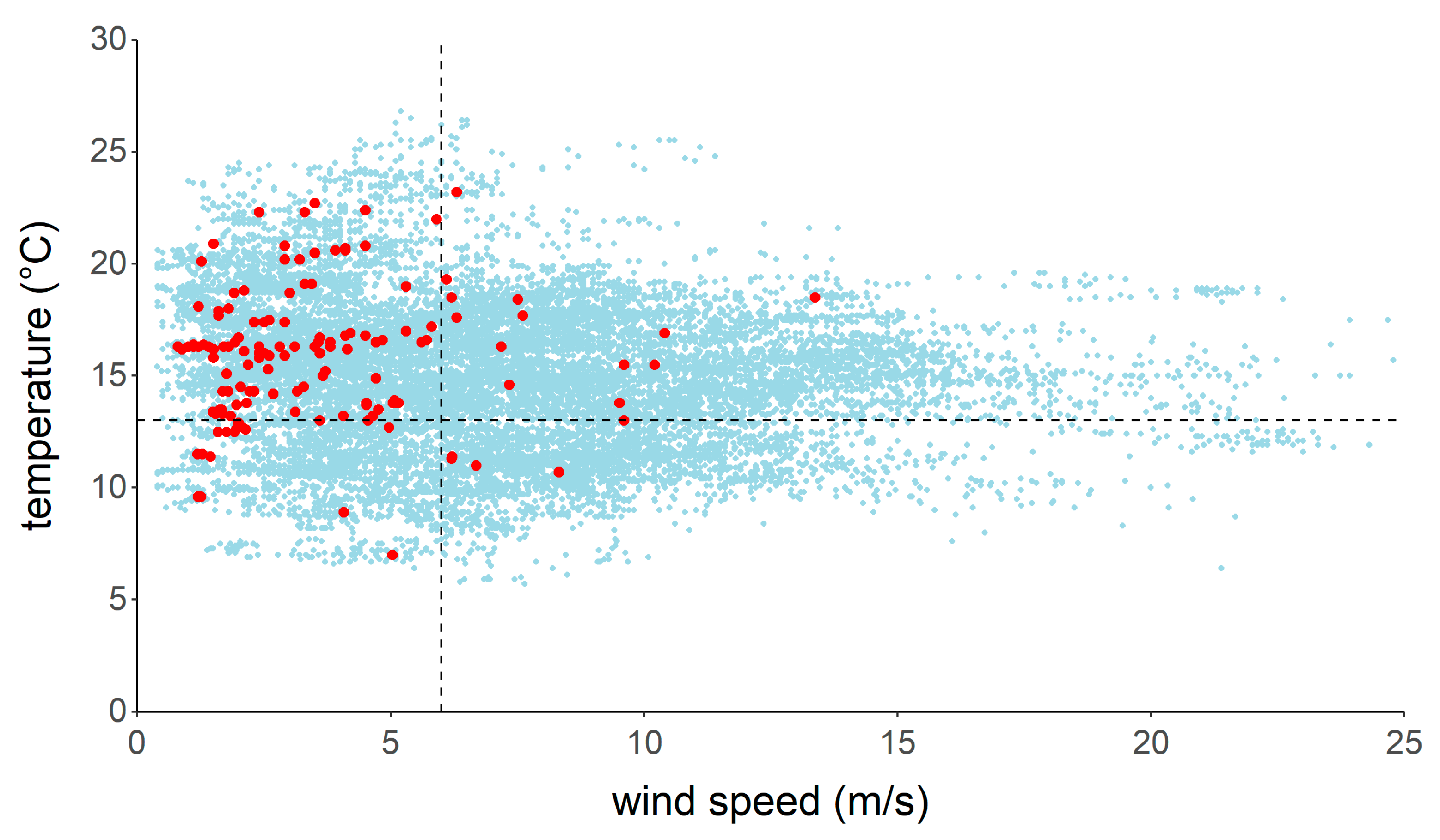
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.J.; Karapandža, B.; Kovač, D.; Kervyn, T.; Dekker, J.; Kepel, A.; Bach, P.; Collins, J.; et al. Guidelines for Consideration of Bats in Wind Farm Projects; EUROBATS Publication Series; UNEP/EUROBATS Secretariat: Bonn, Germany, 2014; 133p, ISBN 978-92-95058-30-9. [Google Scholar]
- Walter, G.; Mathes, H.; Joost, M. Fledermauszug uber Nord- und Ostsee—Ergebnisse aus Offshore-Untersuchungen und deren Einordnung in das bisher bekannte Bild zum Zuggeschehen. Nyctalus 2007, 12, 221–233. [Google Scholar]
- Boshamer, J.P.C.; Bekker, J.P. Nathusius’ Pipistrelles (Pipistrellus Nathusii) and Other Species of Bats on Offshore Platforms in the Dutch Sector of the North Sea. Lutra 2008, 51, 17. [Google Scholar]
- Skiba, R. Europäische Fledermäuse, 2nd ed.; Hohenwarsleben: Westarp Wissenschaften, Germany, 2009. [Google Scholar]
- Leopold, M.F.; Boonman, M.; Collier, M.P.; Davaasuren, N.; Fijn, R.C.; Gyimesi, A.; de Jong, J.; Jongbloed, R.H.; Jonge Poerink, B.; Kleyheeg-Hartman, J.C.; et al. A First Approach to Deal with Cumulative Effects on Birds and Bats of Offshore Wind Farms and Other Human Activities in the Southern North Sea; Report C166/14; IMARES: Wageningen, The Netherlands, 2014; 188p. [Google Scholar]
- Brabant, R.; Laurent, Y.; Jonge Poerink, B.; Degraer, S. Activity and Behaviour of Nathusius’ Pipistrelle Pipistrellus Nathusii at Low and High Altitude in a North Sea Offshore Wind Farm. Acta Chiropt. 2019, 21, 341–348. [Google Scholar] [CrossRef]
- Hüppop, O.; Hill, R. Migration Phenology and Behaviour of Bats at a Research Platform in the South-Eastern North Sea. Lutra 2016, 59, 5–22. [Google Scholar]
- Lagerveld, S.; Jonge Poerink, B.; Haselager, R.; Verdaat, H. Bats in Dutch Offshore Wind Farms in Autumn 2012. Lutra 2014, 57, 61–69. [Google Scholar]
- Brabant, R.; Laurent, Y.; Lafontaine, R.-M.; Vandendriessche, B.; Degraer, S. First Offshore Observation of Parti-Coloured Bat Vespertilio Murinus in the Belgian Part of the North Sea. Belg. J. Zool. 2016, 146, 62–65. [Google Scholar] [CrossRef]
- Baerwald, E.F.; Barclay, R. Sciencebased strategies can save bats at wind farms. Bats 2014, 32, 2–4. [Google Scholar]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.-J.; Green, M.; Rodrigues, L.; Hedenström, A. Bat Mortality at Wind Turbines in Northwestern Europe. Acta Chiropt. 2010, 12, 261–274. [Google Scholar] [CrossRef]
- Voigt, C.C.; Popa-Lisseanu, A.G.; Niermann, I.; Kramer-Schadt, S. The Catchment Area of Wind Farms for European Bats: A Plea for International Regulations. Biol. Conserv. 2012, 153, 80–86. [Google Scholar] [CrossRef]
- Lehnert, L.S.; Kramer-Schadt, S.; Schönborn, S.; Lindecke, O.; Niermann, I.; Voigt, C.C. Wind Farm Facilities in Germany Kill Noctule Bats from Near and Far. PLoS ONE 2014, 9, e103106. [Google Scholar] [CrossRef] [Green Version]
- Lagerveld, S.; van der Wal, J.T.; de Vries, P.; Verdaat, H.; Sonneveld, C.; van der Meer, J.; Brabant, R.; Noort, B. Bats at the Southern North Sea in 2017 & 2018; Wageningen Marine Research Report C062/19; Wageningen Marine Research: Den Helder, The Netherlands, 2019; p. 36. [Google Scholar]
- Simon, R.; Hochradel, K.; Mages, J.; Nagy, M.; Naucke, A.; Niermann, I.; Weber, N.; Behr, O. Methoden akustischer Erfassung der Fledermausaktivität an Windenergieanlagen. In Reduktion des Kollisionsrisikos von Fledermäusen an Onshore-Windenergieanlagen (RENEBAT II); Behr, O., Brinkmann, R., Korner-Nievergelt, F., Nagy, M., Niermann, I., Reich, M., Simon, R., Eds.; Institut für Umweltplanung: Umwelt und Raum: Hannover, Germany, 2015; pp. 39–80. [Google Scholar]
- Voigt, C.C.; Russo, D.; Runkel, V.; Goerlitz, H.R. Limitations of Acoustic Monitoring at Wind Turbines to Evaluate Fatality Risk of Bats. Mamm. Rev. 2021, 51, 559–570. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic. Ecology of European Bats. Species Identification and Studies of Their Habitats and Foraging Behaviour; Biotope Editions, Mèze; Muséum National d’Histoire Naturelle: Paris, France, 2015. [Google Scholar]
- Hüppop, O.; Hilgerloh, G. Flight Call Rates of Migrating Thrushes: Effects of Wind Conditions, Humidity and Time of Day at an Illuminated Offshore Platform. J. Avian Biol. 2012, 43, 85–90. [Google Scholar] [CrossRef]
- Petersons, G. Seasonal Migrations of North-Eastern Populations of Nathusius’ Bat Pipistrellus Nathusii (Chiroptera). Myotis 2004, 41, 29–56. [Google Scholar]
- Vierhaus, H. Pipistrellus Nathusii (Keyserling Und Blasius, 1839)–Rauhhautfledermaus. Handbuch der Säugetiere Europas; Aula: Wiebelsheim, Germany, 2004; pp. 825–873. [Google Scholar]
- Dietz, C.; Kiefer, A. Bats of Britain and Europe; Bloomsbury Publishing: London, UK, 2016; ISBN 1-4729-3576-4. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Modelling for Nested Data. In Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; pp. 101–142. [Google Scholar]
- Rydell, J.; Bach, L.; Bach, P.; Diaz, L.G.; Furmankiewicz, J.; Hagner-Wahlsten, N.; Kyheröinen, E.-M.; Lilley, T.; Masing, M.; Meyer, M.M.; et al. Phenology of Migratory Bat Activity Across the Baltic Sea and the South-Eastern North Sea. Acta Chiropt. 2014, 16, 139–147. [Google Scholar] [CrossRef]
- Flanders Marine Institute (VLIZ), Belgium. LifeWatch Observatory Data: Sensor Network for Bat-Detection. Available online: https://rshiny.lifewatch.be/bats-data/ (accessed on 15 July 2021).
- Eklöf, J.; Šuba, J.; Petersons, G.; Rydell, J. Visual Acuity and Eye Size in Five European Bat Species in Relation to Foraging and Migration Strategies. Environ. Exp. Biol. 2014, 12, 01–06. [Google Scholar]
- Bender, M.J.; Hartman, G.D. Bat Activity Increases with Barometric Pressure and Temperature during Autumn in Central Georgia. Southeast. Nat. 2015, 14, 231–242, 12. [Google Scholar] [CrossRef]
- Muthersbaugh, M.S.; Ford, W.M.; Powers, K.E.; Silvis, A. Activity Patterns of Bats During the Fall and Spring Along Ridgelines in the Central Appalachians. J. Fish Wildl. Manag. 2019, 10, 180–195. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.D.; McWilliams, S.R. Bat Activity during Autumn Relates to Atmospheric Conditions: Implications for Coastal Wind Energy Development. J. Mammal. 2016, 97, 1565–1577. [Google Scholar] [CrossRef]
- Dechmann, D.K.; Wikelski, M.; Ellis-Soto, D.; Safi, K.; O’Mara, M.T. Determinants of Spring Migration Departure Decision in a Bat. Biol. Lett. 2017, 13, 20170395. [Google Scholar] [CrossRef]
- Baerwald, E.F.; D’Amours, G.H.; Klug, B.J.; Barclay, R.M.R. Barotrauma Is a Significant Cause of Bat Fatalities at Wind Turbines. Curr. Biol. 2008, 18, R695–R696. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Behr, M.J.; Gendler, A.; Drake, D.; Dieterle, B.D.; Rudd, R.J.; Walrath, N.L. Investigating the Causes of Death for Wind Turbine-Associated Bat Fatalities. J. Mammal. 2011, 92, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Rollins, K.E.; Meyerholz, D.K.; Johnson, G.D.; Capparella, A.P.; Loew, S.S. A Forensic Investigation Into the Etiology of Bat Mortality at a Wind Farm: Barotrauma or Traumatic Injury? Vet. Pathol. 2012, 49, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Skiba, R. Die Fledermäuse Im Bereich Der Deutschen Nordsee Unter Berücksichtigung Der Gefährdungen Durch Windenergieanlagen (WEA). Nyctalus (Neue Folge) 2007, 12, 199–220. [Google Scholar]
- Ahlén, I.; Baagøe, H.J.; Bach, L. Behavior of Scandinavian Bats during Migration and Foraging at Sea. J. Mammal. 2009, 90, 1318–1323. [Google Scholar] [CrossRef] [Green Version]
- Hatch, S.K.; Connelly, E.E.; Divoll, T.J.; Stenhouse, I.J.; Williams, K.A. Offshore Observations of Eastern Red Bats (Lasiurus Borealis) in the Mid-Atlantic United States Using Multiple Survey Methods. PLoS ONE 2013, 8, e83803. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.M.; Lintott, P.R.; Hosken, D.J.; Economou, T.; Mathews, F. Peaks in Bat Activity at Turbines and the Implications for Mitigating the Impact of Wind Energy Developments on Bats. Sci. Rep. 2021, 11, 3636. [Google Scholar] [CrossRef] [PubMed]
- Rijkswaterstaat. Kavelbesluit V windenergiegebied Hollandse Kust (noord); Rijkswaterstaat: Utrecht, The Netherlands, 2019; p. 96. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Location | N Records | DP10 | Species |
|---|---|---|---|---|
| 2017 | C-Power G01 | 19 | 9 | Pnat |
| 6 | 3 | P40 | ||
| C-Power G03 | 30 | 9 | Pnat | |
| 5 | 3 | P40 | ||
| C-Power H01 | 7 | 3 | Pnat | |
| 4 | 1 | P40 | ||
| C-Power H02 | 25 | 18 | Pnat | |
| 7 | 2 | P40 | ||
| C-Power I01 | 17 | 9 | Pnat | |
| 1 | 1 | P40 | ||
| C-Power I03 | 7 | 4 | Pnat | |
| C-Power J01 | 16 | 8 | Pnat | |
| 7 | 5 | P40 | ||
| 2020 | Norther | 11 | 4 | Ppip |
| 1 | 1 | Pnat | ||
| Northwester II | 1 | 1 | Ppip | |
| 2 | 1 | Pnat | ||
| 1 | 1 | P40 | ||
| Scheur-Wielingen | 31 | 7 | Ppip | |
| 111 | 32 | Pnat | ||
| 128 | 33 | P40 |
| Year | Location | Distance from the Coast (km) | DP10/Night |
|---|---|---|---|
| 2017 | C-Power G01 | 29.3 | 0.12 |
| 2017 | C-Power G03 | 30.6 | 0.12 |
| 2017 | C-Power H01 | 29.5 | 0.04 |
| 2017 | C-Power H02 | 30.4 | 0.20 |
| 2017 | C-Power I01 | 29.4 | 0.10 |
| 2017 | C-Power I03 | 30.8 | 0.04 |
| 2017 | C-Power J01 | 31.2 | 0.13 |
| 2020 | Norther | 23.4 | 0.05 |
| 2020 | Northwester II | 48.7 | 0.03 |
| 2020 | Scheur-Wielingen | 6.4 | 0.67 |
| Wind Direction | Adjusted DP10 (%) Offshore | Adjusted DP10 (%) Nearshore |
|---|---|---|
| E | 41.9 | 29.6 |
| N | 3.6 | 28.3 |
| NE | 16.4 | 15.7 |
| NW | 1.0 | 7.2 |
| S | 12.2 | 1.2 |
| SE | 17.3 | 8.1 |
| SW | 2.6 | 3.6 |
| W | 5.1 | 6.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brabant, R.; Laurent, Y.; Jonge Poerink, B.; Degraer, S. The Relation between Migratory Activity of Pipistrellus Bats at Sea and Weather Conditions Offers Possibilities to Reduce Offshore Wind Farm Effects. Animals 2021, 11, 3457. https://doi.org/10.3390/ani11123457
Brabant R, Laurent Y, Jonge Poerink B, Degraer S. The Relation between Migratory Activity of Pipistrellus Bats at Sea and Weather Conditions Offers Possibilities to Reduce Offshore Wind Farm Effects. Animals. 2021; 11(12):3457. https://doi.org/10.3390/ani11123457
Chicago/Turabian StyleBrabant, Robin, Yves Laurent, Bob Jonge Poerink, and Steven Degraer. 2021. "The Relation between Migratory Activity of Pipistrellus Bats at Sea and Weather Conditions Offers Possibilities to Reduce Offshore Wind Farm Effects" Animals 11, no. 12: 3457. https://doi.org/10.3390/ani11123457
APA StyleBrabant, R., Laurent, Y., Jonge Poerink, B., & Degraer, S. (2021). The Relation between Migratory Activity of Pipistrellus Bats at Sea and Weather Conditions Offers Possibilities to Reduce Offshore Wind Farm Effects. Animals, 11(12), 3457. https://doi.org/10.3390/ani11123457