
The Influence of Biological Factors on Haematological Values in Wild Marsh Harrier (Circus aeruginosus) Nestlings


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Procedures
2.3. Sample Analyses
2.4. Statistical Analyses
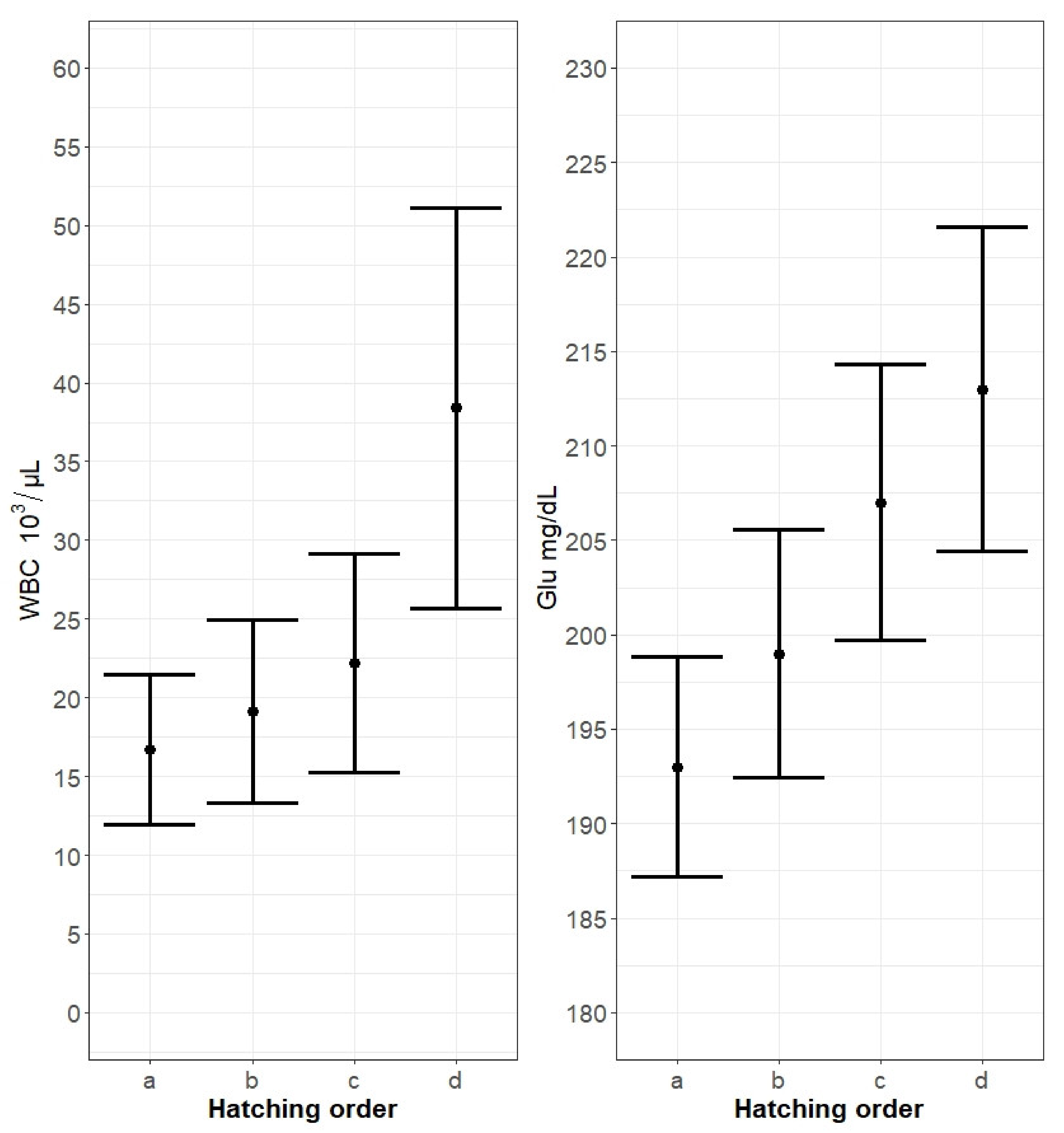
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lanzarot, M.P.; Barahona, M.V.; San Andres, M.I. Hematologic, protein electrophoresis, biochemistry, and cholinesterase values of free-living black stork nestlings (Ciconia nigra). J. Wildl. Dis. 2005, 41, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Karki, B.; Lamichhane, B.R.; Sadaula, A.; Khadka, B.B.; Bhusal, K.P. Hematological Study of Captive White-Rumped Vultures (Gyps bengalensis) to Assess Their Health Status. J. Avian Med. Surg. 2020, 34, 343–347. [Google Scholar] [CrossRef]
- Dein, F.J. Hematology. In Clinical Avian Medicine and Surgery; Harrison, G.J., Harrison, L.R., Eds.; WB Saunders Co: Philadelphia, PA, USA, 1986; pp. 174–191. [Google Scholar]
- Campbell, T.W. Hematology of birds. In Avian and Exotic Animal Hematology and Cytology, 3rd ed.; Campbell, T.W., Ellis, C.K., Eds.; Blackwell: Ames, IA, USA, 2007; pp. 3–50. [Google Scholar]
- Samour, J. Diagnostic Value of Hematology. In Clinical Avian Medicine; Harrison, G.J., Lightfoot, T., Eds.; Spix Publishing: Florida, FL, USA, 2015. [Google Scholar]
- Ferrer, M.; García-Rodríguez, T.; Carrillo, J.C.; Castroviejo, J. Hematocrit and blood chemistry values in captive raptors (Gyps fulvus, Buteo buteo, Milvus migrans, Aquila heliaca). Comp. Biochem. Physiol. A Physiol. 1987, 87, 1123–1127. [Google Scholar] [CrossRef]
- García-Montijano, M.; García, A.; Lemus, A.L.; Montesinos, A.; Canales, R.; Luaces, I.; Pereira, P. Blood chemistry, protein electrophoresis, and hematologic values of captive Spanish imperial eagles (Aquila adalberti). J. Zoo Wildl. Med. 2002, 33, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, V.; Crippa, V.; Marzia, A.; Sartorelli, P. Reference intervals for hematologic and biochemical constituents and protein electrophoretic fractions in captive common buzzards (Buteo buteo). Vet. Clin. Pathol. 2006, 35, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Black, P.; Mcruer, D.; Horne, L.A. Hematologic Parameters in Raptor Species in a Rehabilitation Setting Before Release. J. Avian Med. Surg. 2011, 25, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Villegas, A.; Sánchez, J.M.; Costillo, E.; Corbacho, C. Blood chemistry and haematocrit of the black vulture (Aegypius monachus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; Margalida, A. Hematology and blood chemistry reference values and age-related changes in wild Bearded Vultures (Gypaetus barbatus). J. Wildl. Dis. 2010, 46, 390–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujowich, M.; Mazet, K.J.; Zuba, R.J. Hematologic and biochemical reference ranges for captive California condors (Gymnogyps californianus). J. Zoo Wildl. Med. 2005, 36, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Gee, G.F.; Carpenter, J.W.; Hensler, G.L. Species differences in haematological values of captive cranes, geese, raptors and quail. J. Wildl. Manag. 1981, 45, 463–483. [Google Scholar] [CrossRef]
- Limiñana, R.; López-Olvera, J.R.; Gallardo, M.; Fordham, M.; Urios, V. Blood Chemistry and Hematologic Values in Free-Living Nestlings of Montagu’s Harriers (Circus pygargus) in a Natural Habitat. J. Zoo Wildl. Med. 2009, 40, 687–695. [Google Scholar] [CrossRef]
- Rehder, B.N.; Bird, M.D. Annual profiles of blood packed cell volumes of captive American kestrels. Can. J. Zool. 2011, 61, 2550–2555. [Google Scholar] [CrossRef]
- Dawson, D.R.; Bortolotti, R.G. Variation in Hematocrit and Total Plasma Proteins of Nestling American Kestrels (Falco sparveius) in the Wild. Comp. Biochem. Physiol. A Physiol. 1997, 117, 383–390. [Google Scholar] [CrossRef]
- Baumbusch, R.; Morandini, V.; Uríos, V.; Ferrer, M. Blood plasma biochemistry and the effects of age, sex, and captivity in Short-toed Snake Eagles (Circaetus gallicus). J. Ornithol. 2021, 1–11. [Google Scholar] [CrossRef]
- Lavin, S.; Cuenca, R.; Marco, I.; Velarde, R.; Viñas, L. Hematology and blood chemistry of the Marsh Harrier (Circus aeruginosus). Comp. Biochem. Physiol. A Physiol. 1992, 103, 493–495. [Google Scholar] [CrossRef]
- González, J.; Hiraldo, F. Some hematological data from Marsh Harriers (Circus aeruginosus) in central Spain. Comp. Biochem. Physiol. A Physiol. 1991, 100, 735–737. [Google Scholar] [CrossRef]
- Witkowski, J. Breeding biology and ecology of the Marsh Harrier Circus aeruginosus in the Barycz valley, Poland. Acta Ornithol. 1989, 25, 223–320. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 April 2020).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: Berlin, Germany, 2002; pp. 323–324. [Google Scholar]
- Muriel, R.; Schmidt, D.; Calabuig, C.; Patino-Martinez, J.; Ferrer, M. Factors affecting plasma biochemistry parameters and physical condition of Osprey (Pandion haliaetus) nestlings. J Ornithol. 2012, 154, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Bowerman, W.W.; Stickle, E.J.; Sikarskie, J.G.; Giesy, P.J. Hematology and serum chemistries of nestling bald eagles (Haliaeetus leucocephalus) in the lower peninsula of MI, USA. Chemosphere 2000, 41, 1575–1579. [Google Scholar] [CrossRef]
- Ferrer, M.; Dobado-Berrios, P. Factors affecting plasma chemistry values of the Spanish Imperial Eagle, Aquila adalberti. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 120, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Casado, E.; Balbontin, J.; Ferrer, M. Plasma chemistry in Booted eagle (Hieraaetus pennatus) during breeding season. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 233–241. [Google Scholar] [CrossRef]
- Montolio, S.A.; Valera, R.C.; González, S.L.; Cray, C.; López, R.M.; Ferron, E.S.; Francisco, O.N.; Sánchez, I.M.; Casas-Díaz, E. Plasma biochemistry RIs and age effect in European strigiformes. Vet. Clin. Pathol. 2018, 47, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Lanzarot, M.P.; Montesinos, A.; San Andrés, I.M.; Rodríguez, C.; Barahona, M.V. Hematological, protein electrophoresis and cholinesterase values of free-living nestling peregrine falcons in Spain. J. Wildl. Dis. 2001, 37, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, P.; Jerzak, L.; Sparks, T.; Johnston, A.; Bochenski, M.; Kasprzak, M.; Wiśniewska, E.; Mroczkowski, S.; Tryjanowski, P. Sex and other sources of variation in the haematological parameters of White Stork Ciconia ciconia chicks. J. Ornithol. 2013, 155, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Neb, A.; Hammouda, A.; Selmi, S. Body condition of Little Egret Egretta garzetta nestlings in relation to hatching order in a southern Tunisian breeding colony. Ostrich 2019, 90, 391–396. [Google Scholar] [CrossRef]
- Kaliński, A.; Bańbura, M.; Glądalski, M.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Zieliński, P.; Cyżewska, I.; Bańbura, J. Long-term variation in hemoglobin concentration in nestling great tits Parus major. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 185, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Nadolski, J.; Skwarska, J.; Kaliński, A.; Bańbura, M.; Śniegula, R.; Bańbura, J. Blood parameters as consistent predictors of nestling performance in great tits (Parus major) in the wild. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 143, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Bańbura, J.; Bańbura, M.; Kaliński, A.; Skwarska, J.; Słomczyński, R.; Wawrzyniak, J.; Zieliński, P. Habitat and year-to-year variation in haemoglobin concentrationin nestling blue tits Cyanistes caeruleus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 572–577. [Google Scholar] [CrossRef]
- Kaliński, A.; Wawrzyniak, J.; Bańbura, M.; Skwarska, J.; Zieliński, P.; Bańbura, J. Haemoglobin concentration and body condition of nestlings great tits Parus major: A comparison of first and second broods in two contrasting seasons. IBIS 2009, 151, 667–676. [Google Scholar] [CrossRef]
- Minias, P. The use of haemoglobin concentrations to assess physiological condition in birds: A review. Conserv. Physiol. 2015, 3, cov007. [Google Scholar] [CrossRef] [Green Version]
- Krams, I.; Suraka, V.; Rantala, M.; Sepp, T.; Mierauskas, P.; Vrublevska, J.; Krama, T. Acute infection of avian malaria impairs concentration of hemoglobin and survival in juvenile altricial birds. J. Zool. 2013, 291, 34–41. [Google Scholar] [CrossRef]
- García-Rodríguez, T.; Ferrer, M.; Carrillo, J.C.; Castroviejo, J. Metabolic responses of Buteo buteo to long term fasting and refeeding. Comp. Biochem. Physiol. A Physiol. 1987, 87, 381–386. [Google Scholar] [CrossRef]
- Montoya, B.; Briga, M.; Jimeno, B.; Moonen, S.; Verhulst, S. Baseline glucose level is an individual trait that is negatively associated with lifespan and increases due to adverse environmental conditions during development and adulthood. J. Comp. Physiol. B. 2018, 188, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Salvante, K. Techniques for Studying Integrated Immune Function in Birds. Auk 2006, 123, 575–586. [Google Scholar] [CrossRef]
- Ots, I.; Murumägi, A.; Hõrak, P. Hematological health state indices of reproducing great tits. Methodology and sources of natural variation. Funct. Ecol. 1998, 12, 700–707. [Google Scholar] [CrossRef]
- Parejo, D.; Silva, N.; Avilés, J.M. Within-brood size differences affect innate and acquired immunity in roller Coracias garrulus nestlings. J. Avian Biol. 2007, 38, 717–725. [Google Scholar] [CrossRef]
- Słomczyński, R.; Kaliński, A.; Wawrzyniak, J.; Bańbura, M.; Skwarska, J.; Zieliński, P.; Bańbura, J. Effects of experimental reduction in nest micro-parasite and macro-parasite loads on nestling hemoglobin level in blue tits Parus caeruleus. Acta Oecol. 2006, 30, 223–227. [Google Scholar] [CrossRef]
- Zaremba, U.; Kasprzykowski, Z.; Golawski, A. Variation in growth patterns of Marsh Harrier Circus aeruginosus nestlings: Effects of nestling subperiod, brood size, hatching order and weather conditions. J. Ornithol. In press.
- Kaliński, A.; Bańbura, M.; Glądalski, M.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Zieliński, P.; Bańbura, J. Physiological condition of nestling great tits (Parus major) declines with the date of brood initiation: A long term study of first clutches. Sci. Rep. 2019, 9, 9843. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.; Jenni-Eiermann, S.; Jenni, L. Heterophils/Lymphocytes-ratio and circulating corticosterone do not indicate the same stress impose on Eurasian kestrel nestlings. Funct. Ecol. 2011, 25, 566–576. [Google Scholar] [CrossRef]
- Yosef, R.; Gomboobaatar, S.; Bortolotti, G. Sibling Competition Induces Stress Independent of Nutritional Status in Broods of Upland Buzzards. J. Raptor Res. 2013, 47, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Bakaloudis, E.D.; Papakosta, A.M.; Guy-Yosef, M.; Kosicki, Z.J.; Goutner, V.; Vlachos, G.C.; Yosef, R. Sibling competition affects body condition and allostatic load in the colonial nesting lesser kestrel Falco naumanni. J. Vertebr. Biol. 2020, 69, 1–9. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Blood Parameter | Mean ± SD | Median | Minimum and Maximum Values |
|---|---|---|---|
| First blood collection (N = 58) | |||
| RBC [106/µL] | 1.70 ± 0.77 | 1.61 | 0.49–5.02 |
| WBC [103/µL] | 45.65 ± 37.41 | 31.79 | 0.33–173.80 |
| Hb [g/L] | 124.75 ± 66.42 | 115 | 30–430 |
| Glu [mg/dL] | 202.91 ± 25.15 | 205 | 152–262 |
| Second blood collection (N = 46) | |||
| RBC [106/µL] | 2.02 ± 0.66 | 2 | 0.60–3.67 |
| WBC [103/µL] | 31.31 ± 25.79 | 19.79 | 1.04–98.87 |
| Hb [g/L] | 132.53 ± 50.14 | 120 | 44–250 |
| Glu [mg/dL] | 187.68 ± 39.68 | 195 | 26–280 |
| Third blood collection (N = 14) | |||
| RBC [106/µL] | 1.91 ± 0.86 | 1.63 | 1.18–3.98 |
| WBC [103/µL] | 16.12 ± 10.46 | 13.15 | 5–39.58 |
| Hb [g/L] | 139.64 ± 77.27 | 117.5 | 25–290 |
| Glu [mg/dL] | 191.21 ± 26.82 | 184 | 161–254 |
| Hb | RBC | |||||
|---|---|---|---|---|---|---|
| Fixed Effects | Estimate | SE | p-Value | Estimate | SE | p-Value |
| Intercept | 4.843 | 0.152 | < 0.001 | 0.603 | 0.123 | < 0.001 |
| Hatching order b | –0.164 | 0.112 | 0.145 | –0.003 | 0.092 | 0.973 |
| Hatching order c | –0.196 | 0.118 | 0.095 | –0.098 | 0.096 | 0.310 |
| Hatching order d | –0.295 | 0.137 | 0.031 | –0.230 | 0.111 | 0.038 |
| Random effects | Variance | SD | Variance | SD | ||
| Nestling Nest | 0.028 0.045 | 0.167 0.212 | 0.018 0.017 | 0.134 0.131 | ||
| Blood collection | 0.002 | 0.050 | 0.007 | 0.084 | ||
| Residual | 0.102 | 0.319 | 0.083 | 0.288 | ||
| WBC | Glu | |||||
|---|---|---|---|---|---|---|
| Fixed Effects | Estimate | SE | p-Value | Estimate | SE | p-Value |
| Intercept | 2.326 | 0.460 | < 0.001 | 205.528 | 11.549 | < 0.001 |
| Hatching order b | 0.135 | 0.164 | 0.412 | 5.796 | 5.861 | 0.339 |
| Hatching order c | 0.283 | 0.179 | 0.114 | 13.435 | 6.541 | 0.048 |
| Hatching order d | 0.831 | 0.206 | < 0.001 | 19.616 | 7.775 | 0.017 |
| Brood size 2 | 0.899 | 0.489 | 0.066 | –23.003 | 13.717 | 0.099 |
| Brood size 3 | 0.829 | 0.483 | 0.086 | –7.713 | 13.308 | 0.564 |
| Brood size 4 | 0.235 | 0.452 | 0.604 | –18.998 | 12.565 | 0.137 |
| Random effects | Variance | SD | Variance | SD | ||
| Nestling | 0.043 | 0.208 | 26.04 | 5.103 | ||
| Nest | 0.171 | 0.414 | 88.98 | 9.433 | ||
| Blood collection | 0.077 | 0.277 | 27.91 | 5.283 | ||
| Residual | 0.351 | 0.593 | 538.49 | 23.205 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaremba, U.; Kasprzykowski, Z.; Kondera, E. The Influence of Biological Factors on Haematological Values in Wild Marsh Harrier (Circus aeruginosus) Nestlings. Animals 2021, 11, 2539. https://doi.org/10.3390/ani11092539
Zaremba U, Kasprzykowski Z, Kondera E. The Influence of Biological Factors on Haematological Values in Wild Marsh Harrier (Circus aeruginosus) Nestlings. Animals. 2021; 11(9):2539. https://doi.org/10.3390/ani11092539
Chicago/Turabian StyleZaremba, Urszula, Zbigniew Kasprzykowski, and Elżbieta Kondera. 2021. "The Influence of Biological Factors on Haematological Values in Wild Marsh Harrier (Circus aeruginosus) Nestlings" Animals 11, no. 9: 2539. https://doi.org/10.3390/ani11092539