1. Introduction
The gastrointestinal (GI) tract digests and absorbs nutrients while also acting as a barrier against harmful substances and pathogens ingested through the diet [
1]. Pigs are exposed to a variety of pathogenic challenges, which causes the GI immune system to become activated [
2]. Although a highly activated immune system may appear to be the best protective mechanism for animals, it can have a negative impact on animal performance [
3]. For example, the overactivation of proinflammatory cytokines such as tumor necrosis factor alpha (TNF-α) and interleukin-6 (IL-6) leads to the poor growth of pigs [
4]. Weaned pigs face problems of post-weaning diarrhea (PWD) caused by pathogenic bacteria such as
Shiga toxigenic Escherichia coli (STEC) and
enterotoxigenic Escherichia coli (ETEC) during the 4 weeks of the postweaning period [
5,
6]. The small intestines of pigs show significant structural and functional changes after weaning. These changes lead to the vicissitude of protein conversion rates, microbiota composition, digestive barrier, and immune function [
7]. In previous studies, PWD caused a lower growth performance through changes in the gut integrity, microbiome, and villus height, as well as proinflammatory cytokines [
8,
9]. Many researchers have been studying nonantibiotic biological methods, such as using essential oils, enzymes, probiotics, prebiotics, and organic acids, to prevent digestive diseases by controlling intestinal microbiota [
10,
11].
Diets in monogastric animals consist mostly of corn and non-starch polysaccharides such as arabinoxylans in Asia [
12]. Endo-β-1,4-xylanase (XYL) is a carbohydrate-active enzyme that can hydrolyze the bonds of xylans, thus improving the availability of the antinutritive factors [
13]. Xylo-oligosaccharide (XOS) produced through XYL can act as a prebiotic to increase the fermentation metabolites [
14]. The concept of stimbiotic (STB), a complex of XYL and XOS, has been recently introduced as a nondigestible and fermentable additive that can improve the fermentation of the fiber microbiome [
15,
16]. The mechanism of action of STB is that it can stimulate the intestinal microbiota responsible for fiber degradation [
17]. In a previous study, Bedford et al. [
18] reported that STB can enhance the growth performance and production of short chain fatty acids (SCFAs). Likewise, the supplementation of STB can stimulate the fermentation of dietary fiber, thereby decreasing the digesta viscosity and improving energy utilization [
19]. Petry et al. [
20] reported that the supplementation of XYL and XOS can improve the intestinal barrier integrity and reduce oxidative stress, respectively. Additionally, poor sanitary conditions, which induce bacterial infection found in industrial pig production, can negatively affect the growth performance and TNF- α, which could alleviate the use of STB [
16].
STEC is known to be a major pathogen causing diarrhea, but there is insufficient research validating what causes PWD [
21]. Therefore, we conducted experiments to verify the effects of STEC, which induce PWD. In this experiment, we hypothesized that (1) the experimental induction of STEC infection could increase the inflammatory responses and reduce the growth performance of pigs and that (2) the supplementation of STB could attenuate the extent of the performance loss and inflammatory response caused by PWD induced by STEC challenge. Therefore, the purpose of this study was to determine the effects of adding STB on the growth performance, immune response, and inflammatory response when weaned pigs were orally administered with pathogenic
E. coli.
2. Materials and Methods
2.1. Ethics
All experimental procedures received prior approval from the Animal Ethics Committee of Chungbuk National University (CBNUA-1618-21-02).
2.2. Bacterial Strains, Culture and Challenge
STEC F18 was provided in stock form. The F18
E. coli expressed heat labile toxin (LT) and
shiga toxin type 2e (stx2e). Ten microliter of thawed
E. coli stock was inoculated into 10 mL of nutrient broth and cultured at 37 °C for 24 h and then subcultured [
22]. Thereafter, the subcultured
E. coli was smeared on MacConkey agar to confirm the bacterial enumeration. A final concentration of 1.2 × 10
10 CFU/mL was used in this study.
2.3. Animals, Experimental Design and Diets
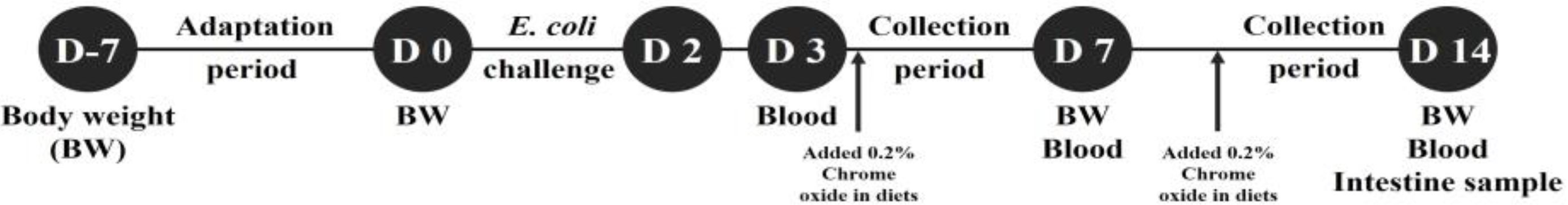
A total of 36 male pigs (Duroc × Yorkshire × Landrace), weaned at 28 d (initial body weight of 8.49 ± 0.10 kg), were assigned to 6 treatments with 6 replicates per treatment. Pigs were individually placed in 45 × 55 × 45 stainless steel metabolism cages in an environmentally controlled room. Pigs were housed in individual pens for 21 days, including 7 days before and 14 days after the first
E. coli challenge (0 dpi). The experiment was conducted in a 2 × 3 factorial arrangement of treatments consisting of two levels of challenge (challenge and non-challenge) and three levels of STB (0, 0.5, and 1 g/kg diet). Corn and soybean meal basal diets were formulated to meet or exceed the nutrient requirements for the weaned piglets by NRC (
Table 1) [
23]. The pigs were fed daily at 8:30 and 17:00 h and had ad libitum access to water. Feed residues were removed before the next meal and considered in the calculations.
Figure 1 depicts the schematic diagram of the weaned piglets experimental design used for this study.
2.4. Growth Performance
All piglets were weighed every week during the experiment period, and feed consumption was recorded to calculate the average daily gain (ADG), average daily feed intake (ADFI), and gain-to-feed ratio (G:F).
2.5. Fecal Scores
The fecal scores were individually recorded at 08:00 and 17:00 by the same person during the entire experimental period. The fecal score was scored using a method used by Zhao et al. [
24]. The fecal scores were as follows: 0, Normal feces; 1, Soft feces; 2, Mild diarrhea; and 3, Severe diarrhea.
2.6. Nutrient Digestibility
To estimate the digestibility, 0.2% chromium oxide (Cr
2O
3) was supplemented with the diets as an indigestible marker. Pigs were fed diets mixed with chromium oxide for 4 consecutive days from 4 dpi to 11, fresh excreta samples were collected in that period. At the end of the experiment, the fecal samples were stored at −20 °C and dried at 70 °C for 72 h and then ground up to pass through a 1-mm screen. All analysis items (feed and fecal) were analyzed for DM and CP. The procedures utilized for the determination of dry matter (DM) and crude protein (CP) digestibility were conducted with the methods by the AOAC [
25]. Chromium was analyzed with an ultraviolet absorption spectrophotometer (UV-1201, Shimadzu, Kyoto, Japan). The digestibility was calculated using the following formula: digestibility (%) = [1 − (Nf × Cd)/(Nd × Cf)] × 100, where Nf is the nutrient concentration in feces (% DM), Nd is the nutrient concentration in diet (% DM), Cd is the chromium concentration in diet (% DM), and Cf is the chromium concentration in feces (% DM).
2.7. Blood Profile
Blood samples were obtained from the anterior vena cava of 6 pigs per each treatment at 3, 7, and 14 dpi. At the time of collection, blood samples were collected into vacuum tubes containing K3EDTA for CBC analysis and nonheparinized tubes for serum analysis, respectively. After collection, blood samples were centrifuged (3000× g for 15 min at 4 °C). The white blood cells (WBC), basophils, neutrophils, and lymphocyte levels in the whole blood were measured using an automatic blood analyzer (ADVIA 120, Bayer, NY, USA). The immunoglobulin G (IgG) and immunoglobulin A (IgA) levels were gauged using an automatic biochemistry blood analyzer (Hitachi 747; Hitachi, Tokyo, Japan).
2.8. Morphological Analysis of Small Intestine
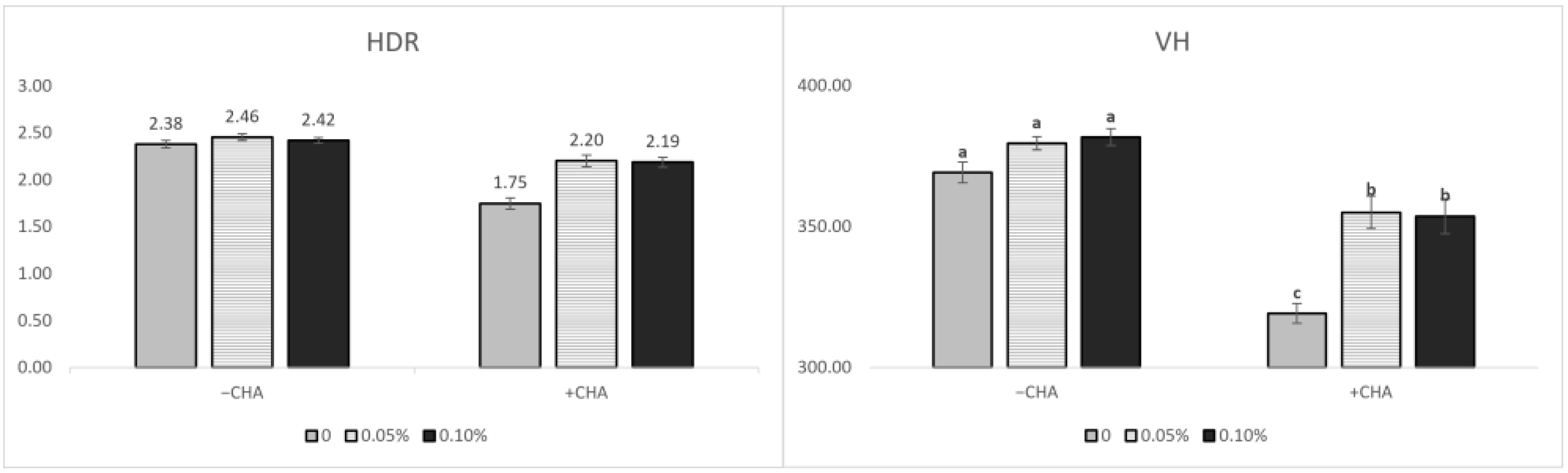
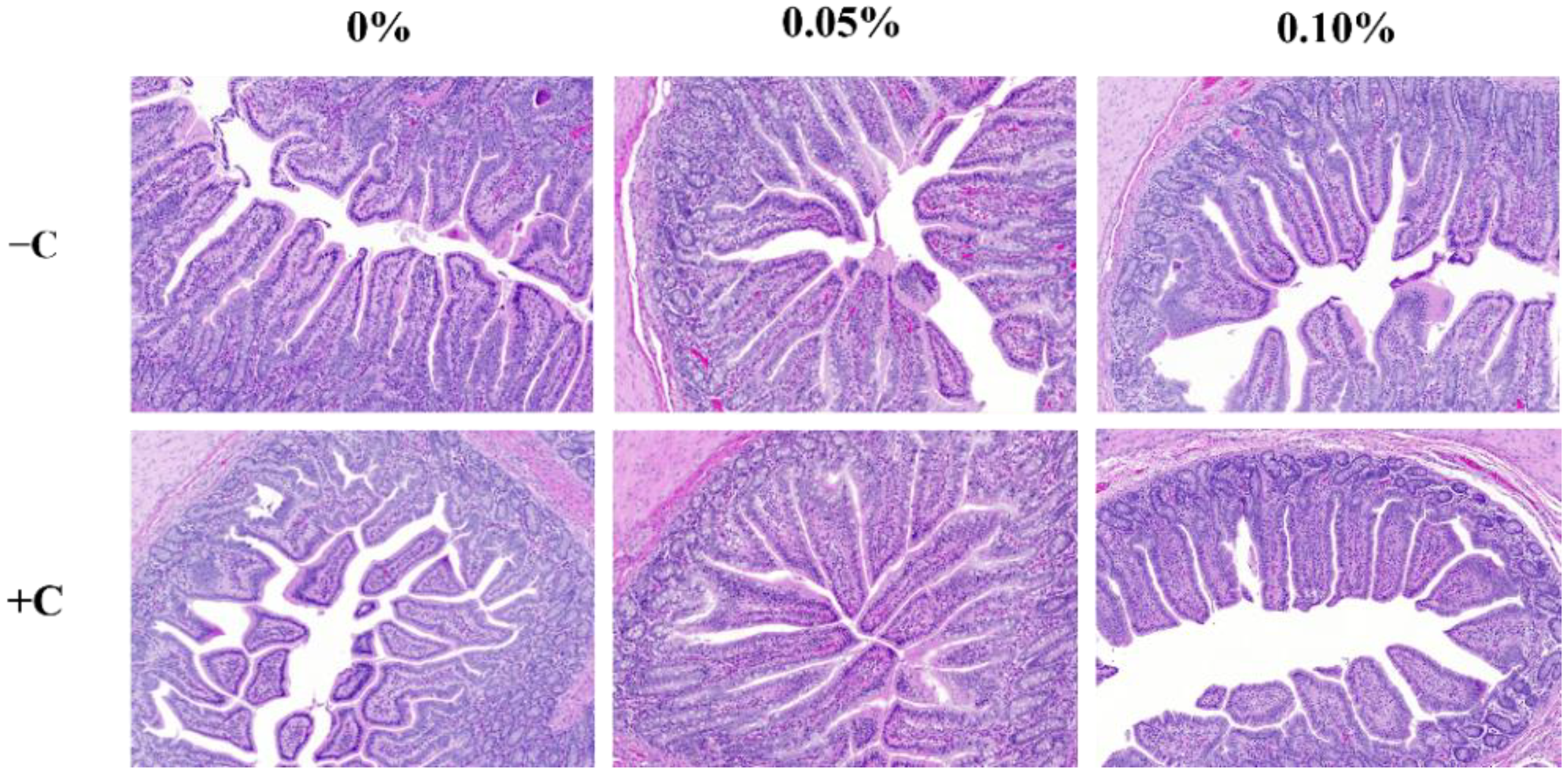
At the end of the experiment (14 dpi), the pigs were anesthetized with carbon dioxide gas after blood sampling and euthanized by exsanguination. Intestinal tissues of about 10 cm from the ileum (close to the ileocecal junction), were collected and fixed in 10% neutral buffered formalin (NBF; Sigma-Aldrich, St. Louis, MO, USA). After cutting the intestine sample, it was dehydrated and dealcoholized. The samples were then installed on slides, treated with paraffin, and stained with hematoxylin and eosin (ab245880, abcam). Villus height and crypt depth were measured under a light microscope (OLYMPUS DP71, BX50F-3, Olympus Optical Co. Ltd., Tokyo, Japan). Villus height (VH) was determined by measuring the distance between the tip of the villi to the villus crypt junction, and the crypt depth (CD) was determined by measuring the distance between adjacent villi. Mean values of 10 fields, 30 well-oriented, complete villus-crypt structures were calculated for each pig.
2.9. Measurements of Pro-Inflammatory Cytokine
The inflammatory biomarkers such as interleukin-6 (IL-6) and tumor necrosis factor α (TNF-α) were measured using commercially available ELISA kits according to manufacturer’s instructions (Quantikine, R&D Systems, Minneapolis, MN, USA).
2.10. Expression of Tight Junction Proteins
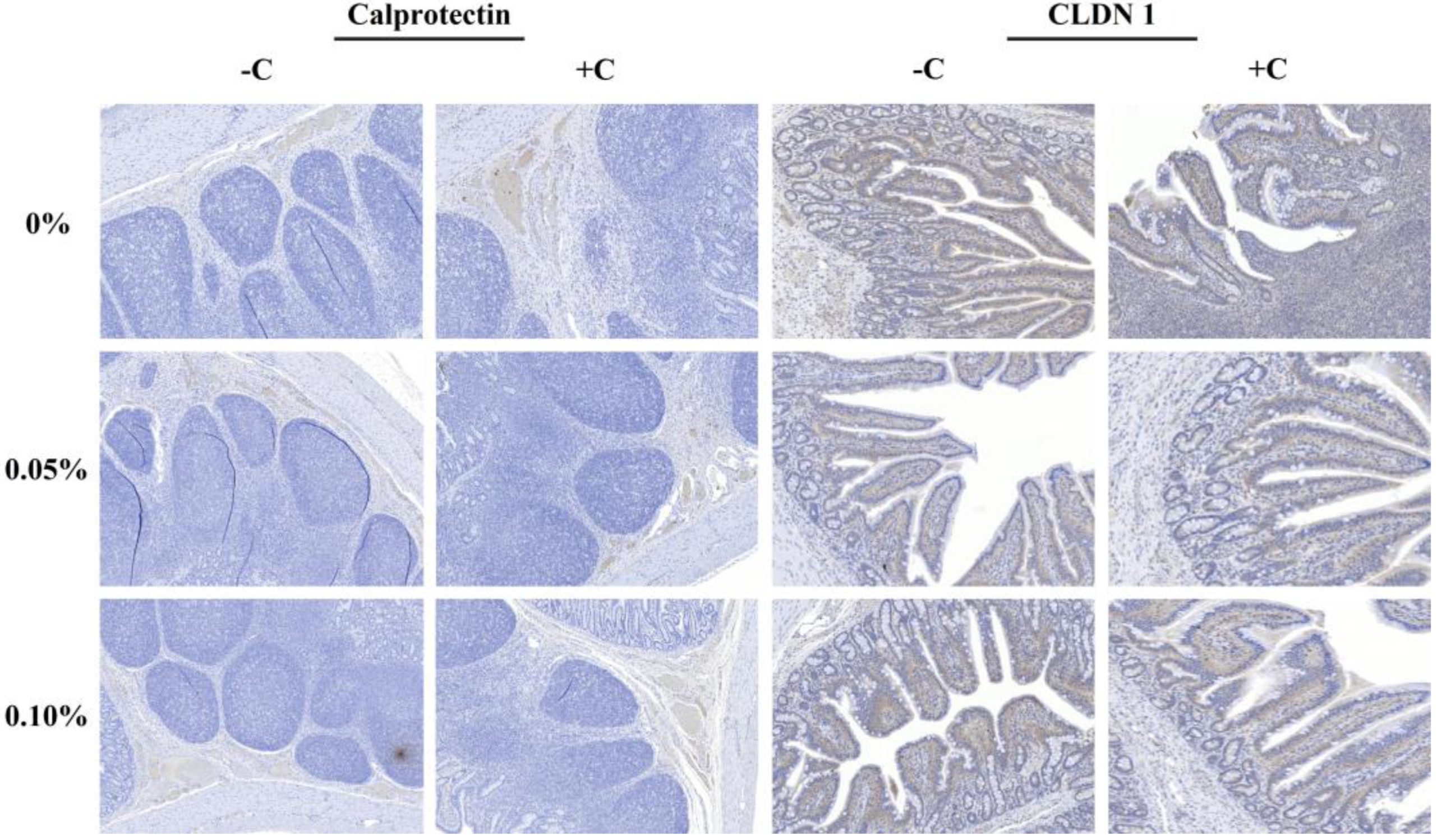
The expression of claudin-1 and calprotectin was determined via immunohistochemistry. Histologic tissues were deparaffinized, rehydrated, and rinsed using standard methods. The sections of slide were incubated with the primary antibody for claudin-1 (1:200; Novus Biologicals, Minneapolis, MN, USA) and calprotectin (1:800; Thermo Fisher Scientific, Waltham, MA, USA), followed by washing and incubation with the secondary antibody envision anti-rabbit for claudin-1 (Dako, Santa Clara, CA, USA) and calprotectin (Dako, Santa Clara, CA, USA) for 30 min. The stained samples were evaluated under a microscope (Axio Scan Z1; Carl Zeiss, Jena, Germany), and the images were analyzed via Zen 3.4 blue edition.2.11. Statistical Analysis.
Data for the effects of different levels of STB were added with a challenge or not. Data were subjected to two-way ANOVA. All data were statistically analyzed with a PROC General Linear Models (GLM) of SAS (SAS Institute, Cary, NC, USA). Differences between treatment groups were measured using Duncan’s multiple range test, with a p-value of less than 0.05 designating statistical significance.
4. Discussion
The small intestine is a major area to digest and absorb nutrients. It can serve as the first line of defense against various harmful substances or pathogens [
26]. Weaning causes numerous changes, including enzymatic, morphological, and inflammatory changes that can, damage the intestinal integrity [
27]. Harmful bacterial pathogens (i.e.,
Escherichia coli) can invade through the damaged intestine and lead to decreased nutrient digestion and absorption, consequently decreasing the growth rate [
28,
29]. In the present study, the STEC challenge decreased the growth performance. The ADG and G:F were reduced by 31% and 23%, respectively. These results were agreement with the results of He et al. [
30], which reported that the challenge with
E. coli decreased the BW, ADG, ADFI, and G:F compared to the non-challenged groups. A previous study showed that
E. coli infection can increase the frequency of diarrhea [
31,
32]. This result is similar to those obtained in prior studies using an
E. coli strain to inoculate pigs. In the present study, the VH and HDR were decreased, but the CD was increased in STEC-challenged pigs. These results are consistent with previous studies showing that pigs challenged with
E. coli had a lower VH and HDR in the small intestine [
29,
33,
34,
35]. The
E. coli challenge could cause a defect of the intestinal barrier integrity by downregulating the expression of tight junction (TJ) proteins. [
36,
37]. The STEC challenge significantly decreased the CLDN-1 and Calp expression compared to the non-challenge groups in the present, as in the previous study of Yu et al. [
34]. TJ proteins, located in the intercellular structure, are junctional adhesion molecules and multi-protein complexes composed of transmembrane proteins [
38,
39]. They are commonly considered as a strong barrier against the absorption of endotoxin [
39]. These results indicated that the oral administration of STEC successfully induced the PWD model, as we hypothesized before starting this study.
STB is a fermentable additive that can stimulate the development of a proportion of bacterial species involved in fiber degradation [
16,
20]. For example, XOS can improve the intestinal morphology and expression of TJ proteins by improving the gut microbiota communities [
40]. Thus, we hypothesized that dietary STB supplementation could mitigate pathogenic
E. coli-induced intestinal damage by improving the barrier integrity and suppressing inflammation in weaned pigs. In our study, the supplementation of STB improved the intestinal morphology and expression of TJ in pigs challenged with STEC. These results were in agreement with previous results showing that 0.01% and 0.05% XOS could increase the VH in the ileum and HDR in the jejunum [
40,
41]. Consistently, prebiotics (i.e., alginate-oligosaccharide, AOS; mannan-oligosaccharide, MOS) could mitigate the intestinal mucosa injury by improving the VH and HDR [
42,
43]. Additionally, the supplementation of MOS could elevate the expression levels of TJ proteins such as zonula occludens-1, CLDN-1, and Occludin reduced by the ETEC challenge [
43]. These results were similar to the results of the present study. Previous studies have shown that prebiotics could upregulate the expression levels of the intestinal TJ proteins in piglets [
44,
45]. Through many studies, including ours, the consistent improvement of oligosaccharides might be associated with the production of SCFAs. SCFAs are volatile fatty acids (VFA). They mainly include acetic acid, propionic acid, and butyric acid [
46]. SCFAs can promote the intestinal morphology and TJ proteins of broilers [
47,
48]. They can also be used as energy sources, leading to an improved absorption surface in the intestine via increased the proliferation of epithelial cells [
49,
50,
51,
52]. The exact mechanism of action of STB is currently unknown. It has been suggested that the improvement of the gut integrity is due to improved gut fermentation.
In the current study, STB supplementation alleviated the reduction of nutrient digestibility and growth performance caused by the STEC challenge. This effect of STB might be due to its ability to improve gut health, as mentioned above. It might also be due to its ability to improve the enzyme activities [
53]. Previous studies have reported that MOS supplementation can increase the mucosal enzyme activities, including duodenal sucrase, ileal lactase, and ileal maltase activities in the ETEC challenged pigs [
43].
The count of WBC is one of the most common diagnostic indications of infection. WBC are an important part of the immune system that fights against infections in the body [
54]. While neutrophils are the first cells to move into infected tissues during inflammatory reactions and phagocytose bacteria with their particles, lymphocytes produce specialized cellular and humoral immune responses [
55]. The neutrophils-to-lymphocytes ratio is often used as a biomarker to assess the systemic inflammation severity. According to Liu et al. [
31],
E. coli can cause inflammation in weaned piglets by boosting WBC counts and neutrophils. A previous study showed that Chito-oligosaccharides with a function similar to XOS can mitigated the increase in the value of neutrophils [
33]. In the current study, during
E. coli infection, pigs supplemented with STB had a lower neutrophils-to-lymphocytes ratio than non-supplemented with the STB groups, indicating that inflammation was decreased in piglets with STB supplementation. These results indicate that STB can reduce
E. coli-induced intestinal inflammation in pigs potentially by lowering the bacterial growth and metabolism in gut bacterial environments [
56].
When
E. coli enters the bloodstream, the general immune response is triggered, and immune cells in tissues are activated by bacterial ligands, resulting in a fast burst of proinflammatory cytokines [
57]. In a previous study, acute exposure to
E. coli can increase the blood endotoxin levels, leukocyte numbers, and proinflammatory cytokine production [
58].
Shiga toxins released by pathogenic
E. coli could cause systemic inflammation, therefore increasing inflammatory cytokines [
59]. Moreover, STEC could utilize hemoglobin as an iron source for their proliferation and virulence production [
60]. A recent study showed that
E. coli can increase the production of proinflammatory cytokines in piglets [
61]. Yu et al. [
43] reported that the supplementation of MOS can reduce the concentrations of inflammatory cytokines. Our study also showed that supplementation with STB reduced the levels of the proinflammatory cytokines. These results confirmed that the
E. coli challenge induces elevated concentrations of inflammatory cytokines and that the supplementation with STB decreases the concentrations of inflammatory cytokines in piglets.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}