Simple Summary
This study was carried out in one of the most densely populated and geographically isolated regions in the Republic of Uzbekistan, the Fergana Valley. The Fergana Valley has the highest level of endemic biodiversity in Uzbekistan (and one of the highest in Central Asia), and the habitats of these endemic species are rapidly being developed for agricultural purposes. Given this development, the remaining areas of habitat are not being adequately protected. The main goal of this study was to obtain up-to-date data on the distribution and abundance of five endemic reptile species in the remaining isolated and undeveloped habitats across the Fergana Valley. One of the most important achievements was the discovery of two unique and new micro-endemic species of gecko genus Alsophylax, which are described herein. These results elevate the number of endemic species in the Fergana Valley and further highlight the urgent need to create state-protected areas of habitat with IUCN I and II protection status for the remaining areas of suitable habitat, which is currently not available.
Abstract
The high level of endemism in Fergana Valley has been well documented in numerous studies for various groups of animals and plants. In a relatively small area, there are 45 endemic plant species, five endemic insect species, and five endemic reptile species. In surveying this area for data on distribution, abundance, acoustics, and genetic samples for species of reptiles, we discovered two new species of gecko from the genus Alsophylax. Phylogenetic analyses of mitochondrial DNA sequences indicate the relatives of these new species are the even-fingered gecko, Alsophylax pipiens, and the southern even-fingered gecko, Alsophylax laevis, located hundreds of kilometers to the northwest and southwest of the Fergana Valley. The threats to these new endemic species are significant given the amount of continued agricultural development that involves new territories previously considered “unsuitable” for any species of significance that is leading to the further reduction in, fragmentation of, and degradation of the remaining natural ecosystems in the Fergana Valley. The conservation of these rare and locally endemic species depends directly on the readiness of the state to create areas with IUCN I and II protection. The many studies documenting levels of endemism, along with the data published in this study, are the basis for the justification for state-protected areas in the Fergana Valley.
Keywords:
reptiles; conservation; taxonomy; Gekkonidae; Alsophylax; Fergana Valley; endemic species; cryptic diversity 1. Introduction
The Fergana Valley is located in the eastern region of Uzbekistan and represents an ancient, isolated ecoregion with unique flora and fauna, much of which is endemic to the valley. In a relatively small area, the Fergana Valley contains several dozen endemic plant species, five endemic species of insects, and five endemic reptile species, most of which are associated with the sandy massifs and foothill habitats in the western and southern areas [1]. With the level of aggressive agricultural development and effects associated with climate change, each of these endemic species is facing its own set of environmental threats. For example, Strauch’s toad agama (Phrynocephalus strauchi Nikolsky, 1899), Said-Aliev’s toad-head agama (Phrynocephalus saidalievi Sattorov, 1981), Fergana sand racerunner (Eremias scripta pherganensis Szczerbak et Washetko, 1973), and Strauch’s even-fingered gecko (Alsophylax loricatus Strauch, 1887) are all sandy massif specialists, and all five species fall into the “risk zone” [2]. These historically undisturbed sandy massifs, which comprise one of the key habitats for endemic species of reptiles in the Fergana Valley, have been reduced to 175.71 sq. km, and there are currently only 13 isolated plots of habitat, which now occupy an area of 0.46–58.66 sq. km [3].
1.1. The Current Status of Conservation in Fergana Valley
Agriculture and textile development has a long history in the Fergana Valley causing the gradual reduction in native habitats. The greatest reduction occurred in the early 20th century due to textiles and vegetable farming, leading to the development of the flatter areas such as sandy habitats. These sandy habitats represent the remains of a vast desert complex in the Fergana Valley originating from the Syrdarya River and are quite old, allowing for adaptation and speciation in these unique microhabitats. The rapid growth of the population of Uzbekistan [3] led to large portions of these habitats being plowed for development. Simultaneously, the rise in population also led to the construction of many agricultural fields, fish ponds, and houses, further fragmenting sandy habitats into sand islands, and preventing migration between adjacent populations. Additionally, excessive watering of adjacent agricultural developments has had a negative impact on existing habitat, resulting in a change to the hydrological regime [3]. This increase in groundwater creates more vegetation growth in these sand habitats, reducing the open cover dune habitat. This reduction in open dune habitat increases interspecific competition between the endemic sand-dune-adapted species and surrounding non-sand-dune-adapted species, making conservation efforts ineffective [4,5]. The problem of preserving sand massif habitats in the Fergana Valley is further complicated by the fact that establishing the correct status and protection regime for these unprotected areas in Uzbekistan is technically quite difficult.
Until 2023, conservation of these endemic reptile species and their habitats was hampered by the fact that, in the Fergana Valley, there were no pre-existing areas with IUCN I and II protection status. The existing natural state monument, “Yazyavan sands” [1,2], and other natural district-level monuments do not meet the requirement for higher-level biodiversity conservation [6]. However, since 2020, the project “Preservation of Key Natural Complexes in the Fergana Valley (Republic of Uzbekistan)” through the Michael Succow Foundation, with support from the World Wildlife Fund (WWF), Critical Ecosystem Partnership Fund (CEPF), and Central Asian Desert Initiative (CADI) project, has been garnering more attention for the conservation of the Fergana Valley. Additionally, the project “Maintenance and Conducting Selective Records of Rare and Endangered Species of Vertebrate Animals of the Fergana Valley”, carried out by the “Cadaster and Cadastre of Rare Vertebrate Animals” at the Institute of Zoology of the Academy of Sciences of the Republic of Uzbekistan, has been documenting new populations of species in the Fergana Valley since 2020. In 2019, our team conducted a National Geographic Explorers expedition to survey reptiles in Uzbekistan including the Fergana Valley, and our team returned for multiple expeditions in the Fergana Valley between late 2019 and late 2022. Our team assessed the current state of the remaining preserved isolated habitat sites; as a result, we were able to update distribution and population density data for many reptile species in Uzbekistan. Most of our surveys were focused on the endemic and endangered species in Fergana Valley. Moreover, during these surveys, we discovered two new micro-endemic habitat specialist species of the small body-sized gekkonid genus, Alsophylax.
1.2. Current Taxonomy of the Gecko Genus Alsophylax
Alsophylax Fitzinger, 1843 originated from the description of Lacerta pipiens by Pallas [7] and was subsequently placed in its own genus, Alsophylax, by Fitzinger [8]. Since then, Alsophylax has received little attention due to the fact that its species are very small in size and have what can be perceived as a conserved morphology across the distribution of the genus. As a result, the taxonomic history of this group has been extremely complicated and confounding. Fortunately, there have been detailed descriptions of useful characteristics for describing new species in recent works [1,9,10,11]. Currently, Alsophylax comprises six species distributed in Central Asia and southern Russia (A. laevis Nikolsky, 1907; A. loricatus Strauch, 1887; A. pipiens (Pallas, 1827); A. przewalskii Strauch, 1887; A. szczerbaki (Golubev & Sattarov, 1979); A. tadjikiensis Golubev, 1979) [1]. To date, no study has investigated the interspecific relationships within Alsophylax, and even its phylogenetic position among other gecko genera remains poorly understood, further exemplifying their understudied nature [12,13,14].
In this study, we provide the most complete phylogeny for Alsophylax on the basis of the mitochondrial barcoding gene cytochrome oxidase I, and we include expanded sampling for species complexes such as A. laevis and A. pipiens. We used this phylogeny along with a dataset of morphological characteristics to demonstrate that these new populations of Alsophylax from the Fergana Valley are distinct species and clearly demonstrate the need for further work in Uzbekistan and the Fergana Valley to find other undescribed Alsophylax species. Lastly, we provide a discussion on the history of herpetological work in the Fergana Valley, highlighting its endemic reptile species and providing updated distribution and population density assessments of these endemic reptiles.
2. Materials and Methods
2.1. Population Density and Field Observations
We sampled individuals using the line transect survey method at each sand massif habitat island. During the field observations, we observed 1288 individuals of seven species along the total route length of 210 km. Diurnal species were estimated from routes with varying width along the transect. We recorded the perpendicular distance from the transect line to each individual reptile that was recorded to calculate the transect width We carried out nocturnal sampling using headlamps at a fixed width along the transect. When surveying Teratoscincus, we also used the counting of the reddish eye reflections with some specimens detectable at quite far distances up to 100 m or more. However, we understand that using a wide census band can underestimate occurrence data. Therefore, we limited it in accordance with the terrain and vegetation cover of the area. The accounting results, average detection distance () of the species and effective width of the census band (B), were calculated. We calculated population density (D) per hectare (ha) using the following formula:
where N is the total number of individual reptiles, n is the number of individuals with measured detection distances, and L is the route length. Comparative analysis of the results obtained using various route accounting methods proved its high accuracy [15]. This method has been successfully employed in multiple studies to survey desert reptile species [16,17,18]. We calculated mean values of the population density along with estimation of their error (M ± m) according to the results at several approximately equal route sections.
We conducted daytime surveys in sunny weather with low wind and no precipitation. We started recording data coinciding with the lizards’ maximum activity, and finished when they shifted to shelters to escape the heat during the maximum daytime temperature. We also recorded substrate temperature.
2.2. Molecular Data and Phylogenetic Analyses
Our dataset included 46 samples belonging to all six Alsophylax species, including the two new species from the Fergana Valley. This also includes one Altiphylax species from Central Asia, which we hypothesize is an Alsophylax, and one sample of Mediodactylus kirmanensis (Nikolsky, 1900) from southern Iran as an outgroup. All analyzed materials are listed in Appendix A.
We extracted total genomic DNA from ethanol-preserved muscle or liver tissues using standard phenol–chloroform extraction procedures [19] followed by isopropanol precipitation. We amplified a fragment of the cytochrome oxidase I (COI) gene with maximal length of 655 bp. This mitochondrial marker is widely used for barcoding in vertebrates [20,21] and has also proven to be useful for species identification in reptiles [22,23,24]. Primers used both for PCR and sequencing were the VF1-d (5′–TTC TCA ACC AAC CAC AAR GAY ATY GG–3′), VR1-d (5′–TAG ACT TCT GGG TGG CCR AAR AAY CA–3′) [25], RepCOI-F (5′–TNT TMT CAA CNA ACC ACA AAG A–3′), and RepCOI-R (5′–ACT TCT GGR TGK CCA AAR AAT CA–3′) [24]. We sequenced fragments in both directions for each sample, and a consensus sequence was generated. We used 25 µL reactions for PCRs with approximately 50 ng of genomic DNA, 10 pmol of each primer, 15 nmol of each dNTP, 50 nmol of additional MgCl2, Taq PCR buffer (10 mM Tris-HCl, pH 8.3, 50 mM KCl, 1.1 mM MgCl2, and 0.01% gelatine), and 1 U of Taq DNA polymerase. We used the following PCR conditions: an initial denaturation step at 95 °C for 3 min; five cycles at 95 °C for 30 s, annealing at 45 °C for 1 min, and extension at 72 °C for 2 min; 35 cycles at 95 °C for 30 s, annealing at 48–51 °C for 1 min, extension at 72 °C for 2 min, final extension of 5 min at 72 °C, and storage at 4 °C. We loaded PCR products onto 1% agarose gels, stained with GelStar gel stain (Cambrex), and visualized in a Dark reader transilluminator (Clare Chemical). If results were satisfactory, products were purified using 2 µL, from a 1:4 dilution of ExoSapIt (Amersham), per 5 µL of PCR product prior to cycle sequencing. The 10 µL sequencing reaction included 2 µL of template, 2.5 µL of sequencing buffer, 0.8 µL of 10 pmol primer, 0.4 µL of BigDye Terminator version 3.1 Sequencing Standard (Applied Biosystems), and 4.2 µL of water. The sequencing reaction was 35 cycles of 10 s at 96 °C, 10 s at 50 °C, and 4 min at 60 °C. We purified cycle sequencing products using ethanol precipitation. We carried out sequence data collection and visualization on an ABI 3730xl automated sequencer (Applied Biosystems). Resulting sequences are deposited in GenBank under accession numbers (see Appendix A).
All Sequences were aligned manually using BioEdit Sequence Alignment Editor 5.0.9 [26]. The final alignment used for all phylogenetic analysis contained 654 bp of COI gene for all samples. MODELTEST v.3.06 [27] was used to estimate the optimal evolutionary models to be used for phylogenetic analysis. The preferred model was (GTR + G), as suggested by the Akaike information criterion (AIC). Confidence in tree topology was estimated with bootstraps in RAxML 8.1.1 [28]. We considered values above 85% as strong support and rather than choose an arbitrary number of bootstrap replicates to run we use internal metrics of the to allow the analysis to stop itself when sufficient replicates have been run.
2.3. Morphological Data and Statistical Analyses
We analyzed 310 specimens of all six recognized species of the genus Alsophylax for 22 morphological characteristics, including 14 metric and eight meristic characteristics. We chose these morphological characters on the basis of previous studies on Alsophylax [9], as well as additional new characteristics that were identified for this study. All specimens used were from Zoological Museum of Moscow State University (ZMMU), Zoological Institute of St. Petersburg (ZISP), Institute of Zoology National Academy of Science, Ukraine (IZ NAS), and Institute of Zoology Academy of Sciences of the Republic of Uzbekistan (IZ) (Appendix A).
Morphological measurements were taken with a digital caliper to the nearest 0.1 mm and included the following list of characteristics (14 in total): snout–vent length (SVL) measured from tip of snout to vent; tail length (TL), measured from vent to tail tip; head length (HL), distance between retroarticular process of jaw and snout tip; head width (HW), measured at the widest point of the head; head height (HeadH), measured from top of occiput to underside of jaws; orbital diameter (OrbD), diameter of orbit; snout-to-eye distance (SnEye), measured between anterior most point of eye and tip of snout; eye-to-ear distance (EyeEar), distance from anterior edge of ear opening to posterior corner of eye; humeral length (LS), measured on ventral surface of arm from base of axilla to posterior margin of elbow while forelimb bent by 90° at elbow; forearm length (ForeaL), measured on dorsal surface while forelimb bent by 90° at the elbow from the posterior margin of elbow to wrist; femur length (FemurL), measured on the ventral surface of the leg from base of femur to knee while hindlimb bent by 90°; crus length or tibia length (CrusL), measured on ventral surface of the of knee to the base of the heel while hindlimb bent by 90°; length of finger IV (LD4A), length of free distal phalanx of forth finger, without claw; length of toe IV (LD4P), length of free distal phalanx of forth toe, without claw.
The following characteristics (eight in total) were examined: number of scales around midbody including ventral and dorsal scales (Sq); number of scales along midbody from mental shield to anterior edge of cloaca (SLB); number of supralabials (SL); number of infralabials (IL); number of scales along middle of head, between eyes (I); number of subdigital lamellae under fourth finger (LF 4); number of subdigital lamellae under fourth toe (LT 4); number of postcloacal spurs on base of tail on both sides (Spurs); number of precloacal pores (PP).
We investigated morphometric differences between species of Alsophylax and the new species described herein. We log10-transformed all meristic measurements to normalize the data normality and increase the homogeneity of variance. We adjusted morphometric measurements to remove the effects of body size variation [29]. We use the allometric formula: Xadj = log(X) − b[log(BL) − log(BLmean)].We conducted a discriminant function analysis (DFA) to characterize the morphological variation between one new population of Alsophylax from the Fergana Valley and A. laevis and A. pipiens.
3. Results
3.1. Population Density and Distribution
Through literary sources, as well as the results of our surveys of the Fergana Valley from 2019 to 2022, we noted 27 species of reptiles, which accounts for 43.5% of all reptile species known from Uzbekistan. Eight (30%) of the 27 Fergana Valley species are considered rare and listed in the Red Book of the Republic of Uzbekistan, five species (18.5%) are listed in the IUCN Red List, and three (11%) are CITES-protected species. Lastly, there are seven (26%) species that are micro-endemic species. The data from our surveys were analyzed and used to make Table 1, which lists all surveyed locations with an estimated average density of each endemic reptile species.

Table 1.
Density of rare and endemic retile species in key locations of the Fergana Valley.
3.2. Molecular Analysis and Morphology
All phylogenetic analyses support the monophyly of Alsophylax (Figure 1). These analyses demonstrate that both A. pipiens and A. laevis are polyphyletic, with significant genetic structure within each of these wide-ranging species complexes. These analyses also indicate that both of these new Alsophylax populations from the Fergana Valley (Figure 2) are represent unique evolutionary lineages, sister to different clades of either A. pipiens or A. laevis. All sequences have been deposited on Genbank and their accession number can be found in Appendix B.
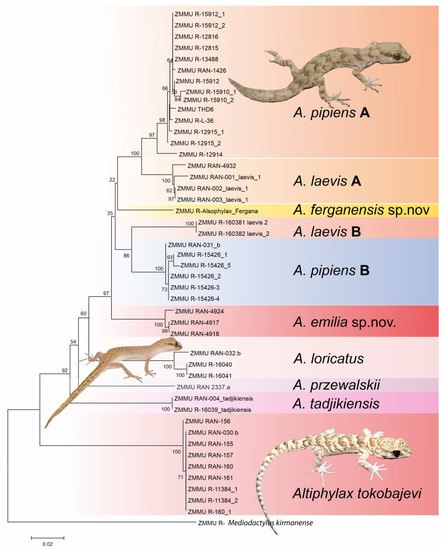
Figure 1.
The gene tree from the maximum likelihood analysis for all described Alsophylax species using cytochrome oxidase I (COI).
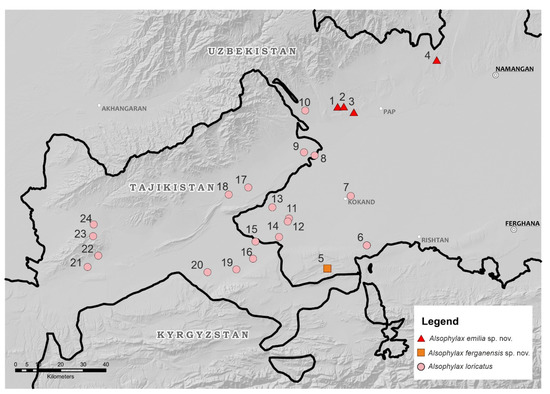
Figure 2.
Distribution of three species of Alsophylax in the Fergana Valley.
Taxonomy
Family Gekkonidae
Genus Alsophylax Fitzinger, 1843
Both new species belong to the genus Alsophylax according to the following morphological characteristics:
(1) Elongate body with relatively short limbs; (2) non-segmented tail approximately the same length as the body; (3) relatively small and roundish head with a short rostral part; (4) no femoral pores and 6–8 precloacal pores; (5) small, roundish and flattened dorsal tubercles are present in some representatives of this genus, or dorsal scales can be smooth and homogeneous in another species.
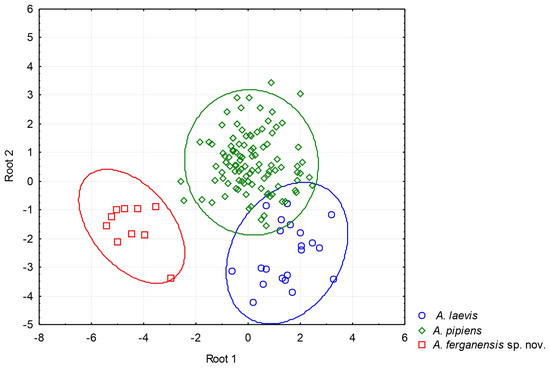
Figure 3.
A plot from the discriminant function analysis for Alsophylax laevis, A. pipiens, and A. ferganensis sp. nov. The results of our discriminant function analysis demonstrate that Alsophylax ferganensis sp. nov. is morphometrically distinct from all clades of A. laevis and A. pipiens. Given the low sample size for A. emlia (n = 3), we were not able to make meaningful statistical comparisons with other species.

Figure 4.
(a) Alsophylax ferganensis sp. nov. sp. nov. in situ and (b) its habitat.
Holotype. Adult male ZMMU Re-17532 (field ID RAN 4358) collected from urban-type settlement Shorssu (N 40.244792 E 70.821582), Uzbekistan District, Fergana Region, Uzbekistan by Timur Abduraupov and Roman Nazarov in 2021 (Figure 5a–e).
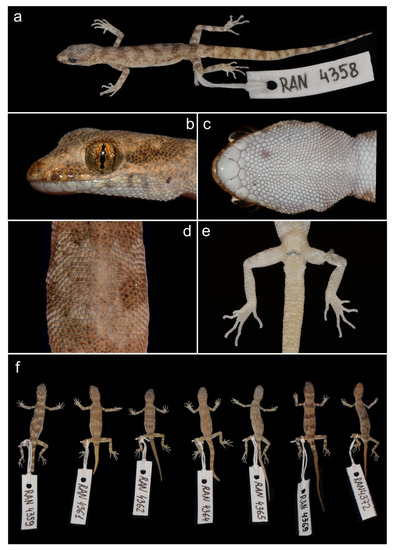
Figure 5.
Holotype of Alsophylax ferganensis sp. nov. ZMMU Re-17532: (a) general view of preserved specimens; (b) lateral view of the head; (c) gular region showing mental, postmental, and chin scale arrangement; (d) middle of the back with homogeneous dorsal scales; (e) precloacal region showing pore and spur arrangements, and plantar view of feet showing subdigital lamellae; (f) general view of part of the paratype series of A. ferganensis sp. nov.
Paratypes. Four males UZZI RE–19077, UZZI RE–19078, ZMMU Re-17537, ZMMU Re-17538 (field ID RAN 4360, RAN 4363, RAN 4370, RAN 4371) and seven females UZZI RE–19075, UZZI RE–19076, Re-17533, Re-17534, Re-17535, Re-17536, Re-17539 (field ID RAN 4359, 4361, 4362, 4364, 4365, 4369, 4372) data same as the holotype (Figure 5f, Table 2).
Table 2.
Measurements of the type series of Alsophylax ferganensis sp. nov.
Diagnosis. Alsophylax ferganensis sp. nov. is tentatively a sister to clades A of A. pipiens and A. laevis (Figure 1) and morphologically closer to A. laevis (Table 3). Alsophylax ferganensis sp. nov. sp. nov. can be distinguished from A. laevis by a smaller maximum body size (SVLmax 31.5 mm versus 38.7 mm) and relatively narrow head, as well as elongated limbs and dorsal patterns with well-defined nuchal loop, and relatively narrower dark transverse bands with approximately equal interspaces in between versus wide transverse patterns and narrow interspaces in A. laevis (Figure 6). Caudal margins have dark transverse bands that are wavy. Dorsal scales are flat, smooth, and roundish, without enlarged dorsal tubercles (Figure 7).
Table 3.
Morphological characteristics and data used in this study.
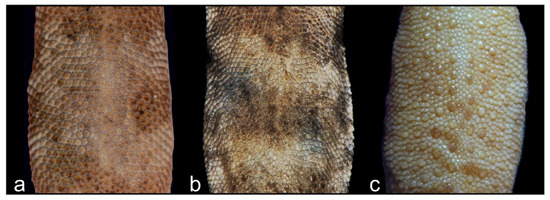
Figure 6.
Three different types of dorsal scalation in Alsophylax: (a) Alsophylax ferganensis sp. nov. with homogeneous dorsal scales; (b) Alsophylax emilia sp. nov. with slightly heterogeneous dorsal scales and irregular enlarged tubercles; (c) A. pipiens with heterogeneous dorsal scales and regular rows of longitudinal enlarged tubercles.

Figure 7.
Different type of subcaudal scalation in Alsophylax species: (a) A. ferganensis sp. nov. with slightly enlarged medial scales; (b) A. emilia sp. nov. with medial row of transverse enlarged plates; (c) A. pipiens (clade B) from type locality (Mt. Bogdo) with approximately equal subcaudal scales; (d) A. pipiens (clade A) from Mongolia ZMMU R-5381, with slightly enlarged medial scales; (e) A. pipiens (clade B) form Ustyurt Plateau, western Uzbekistan ZMMU 15426, with noticeably enlarged medial scales; (f) A. laevis RAN 4957 from central Uzbekistan, Ayagitma depression; (g) A. przewalskii Paralectotypus ZISP 7016 with small homogeneous subcaudal scales; (h) A. Tadjikiensis ZMMU R-16039.
The main features characterizing and distinguishing it from all of other species are as follows: a maximum SVL of 31.7 mm; 7–8 infralabials; 5–6 supralabials; 1–2 pairs of roundish postmentals, first pair in a broad contact; one nasal scale; 18–22 scales between the orbits of the eyes; 83–97 longitudinal ventral scales from postmentals to cloaca; 44–51 scales along the midline around the body; 13–15 subdigital lamellae on the fourth finger; 14–18 subdigital lamellae on the fourth toe; males have 6–7 precloacal pores on enlarged scales; precloacal pores on females are absent or 7–8 enlarged perforated scales can be present; 2–3 cloacal spurs on each side; with the subcaudal scales, the central line has slightly enlarged roundish plates (Table 3).
Description of holotype. This is an adult male SVL 29.6 mm with a small teardrop shaped head (HL/SVL 0.27), smoothly passing into the neck. The rostral part is roundish and short (SnEye/HL 0.37), more elongated than the occiput (SnEye/EyeEar 1.11). The ear opening is small and roundish. There is a relatively large eye (Orb D/HL 0.25). The distance between posterior margin of the eye and ear (EyeEar/HL 0.33) exceeds the diameter of the orbit of the eye. Along the midline of the rostrum, a longitudinal depression is noticeable. The nostril is surrounded by the first supralabial, rostral, and enlarged nasal scale. Seven supralabials and five large infralabials of a rectangular shape are present, which decrease in length in the caudal direction. A large pentagonal mental shield borders on the first labial and two rounded postmentals. There are 18 round–oval scales between the eyes. The upper part of the head is covered by round, small granular scales which are enlarged in the area between the orbits. Large rounded gulars gradually pass into ventral large flat ventral scales (Figure 5b–e).
The body is elongated and slightly flattened, with no lateral folds. The dorsal scales are relatively small compared to the ventral; slightly larger and smaller scales are evenly distributed among them. No enlarged dorsal tubercles. A total of 86 scales are present from postmentals to cloaca, along with 44 around the midbody. Seven elongated, rectangular, perforated precloacal scales form an inverted V-shaped row. The opening of one pore occupies half of the total area of the precloacal scale.
The forelimbs are slender, and the humeral length slightly exceeds the size of the forearm (ForeaL/LS 0.9). The hindlimb is elongated, stronger in structure than the front. The thigh and lower leg are of the same length (CrusL/FemurL 0.1). The forelegs are dorsally covered with smooth juxtaposed scales, while convex granular scales exist on the ventral side of the upper arm, overlapping each other on the forearm. The dorsal surface of the hindlimb is covered by the scales similar to the dorsal ones, in contrast to enlarged scales on the ventral side, smooth and imbricated. The fingers are slender and long. From above, the metacarpus and metatarsus are covered with scales passing from the forearm and lower leg, respectively; those from below the palm and sole are covered with small swollen rounded granules, and the lower surface of the fingers is covered with transverse subdigital lamellae. The fourth digit of the hindlimb is the longest, with 14 slightly swollen subdigital lamellae.
The tail is 36.9 mm longer than the body (SVL/TL 0.8). The base of the tail is slightly swollen; on each side, there are two pairs of postcloacal spurs. The roundish subcaudal plates on the medial line of ventral side of the tail are 1.5–2 times larger than the surrounding scales.
Coloration. The dorsum of the body, head, and limbs are light beige. The pattern on the dorsum is formed by 5–7 transversely dark brown bands with uneven edges; the interspaces between bands is approximately equal to or slightly greater than the width of the bands. There are 7–10 transverse dark stripes on the tail, about the same number as the width of the gaps. Patterns on the head are not pronounced, whereby there is a narrow nuchal loop. The dorsal surface of the limbs is covered with transverse dark bands and irregularly shaped spots. The ventral surface of the body is white.
Distribution. Alsophylax ferganensis sp. nov. is only known from the type locality on the border with Kyrgyzstan, on the southern edge of the Fergana Valley (Figure 2). The distribution of A. ferganensis sp. nov. is probably limited to this small mountain range, and a preliminary estimation of the available habitat indicates there is less than 50 km2. During field observations, another similar population was found on Pap Adyrs, which is located on the opposite side of the Fergana Valley. This isolated population is located 75 km from the type locality of A. ferganensis sp. nov., and its taxonomic status remains unclear.
Habitat and natural history. Clay-variegated canyons with sandstone outcrops represent the habitat of A. ferganensis sp. nov. (Figure 4b). The vegetation in these habitats consists of various shrubs and other plants. Most of the geckos were found on open hills with puffy cracked soil and at the bottom of the canyon along the sides.
Comparison. All representatives of the genus Alsophylax are divided into two groups—one dominant group has enlarged dorsal tubercles (A. loricatus, A. pipiens, A. przewalskii, A. szczerbaki) and the other group does not (A. laevis, A. tadjikiensis). Alsophylax ferganensis sp. nov. belongs to the second group and can be distinguished from A. loricatus, A. pipiens, A. przewalskii, and A. szczerbaki by granular homogeneous dorsal scales without enlarged tubercles (Figure 6). A. ferganensis sp. nov. sp. nov. is morphologically closest to A. laevis but differs further by smaller maximal body size, relatively elongate limbs, and a relatively smaller head that is not sharply detached from the body; dorsal patterns are relatively narrower dark transverse bands with approximately equal interspaces in between, in contrast to the well-defined nuchal loop in A. laevis with wide transverse dorsal patterns and narrow interspaces. Alsophylax ferganensis sp. nov. differs from A. tadjikiensis by the following features: 6–8 precloacal pores in new species versus 5–7 pores in A. tadjikiensis; transversal dorsal bands versus pattern-less dorsum; bright yellowish or orange coloration of tail in A. tadjikiensis.
Etymology. This species is named after the Fergana Valley where it is endemic.

Figure 8.
(a) Alsophylax emilia sp. nov. holotype and (b) its habitat.
Holotype. This is an adult male ZMMU Re-17544 (field ID RAN 4917) collected from Uzbekistan, Fergana Valley, Kokand region, in the vicinity of Jildalisoy reservoir, N 40.891349, E 70.884367, by Roman Nazarov, Evgeniya Shepelya, and Mariya Gritsina on 17 June 2022 (Figure 9a).

Figure 9.
Holotype of Alsophylax emilia sp. nov. (ZMMU Re-17544): (a) general view of preserved specimen; (b) lateral view of the head; (c) gular region showing mental, postmental, and chin scale arrangement; (d) middle of the dorsum with heterogeneous dorsal scales; (e) precloacal region showing scalation pore and spur arrangements; (e) plantar view of feet showing subdigital lamellae.
Paratypes. The adult male UZZI RE–19079 (field ID RAN 4918) collected in the same place as a holotype and ZMMU Re-17545 (field ID RAN 4924) collected from Uzbekistan, Fergana Valley, Namangan region, vicinity of Yartepa village, N 41.067757, E 71.412176, on 18 June 2022 by Roman Nazarov, Evgeniya Shepelya and Bogatova Polina (Table 4).
Table 4.
Measurements of the type series of Alsophylax emilia sp. nov.
Diagnosis. Alsophylax emilia sp. nov. is a sister to the clade containing lineages of A. pipiens, A. laevis, and A, ferganensis sp. nov. (Figure 1). Alsophylax emilia sp. nov. is morphologically closest to the A. pipiens (Figure 3; Table 3) and can be distinguished by the following features: enlarged dorsal tubercles flat roundish smooth and larger than surrounding scales by no more than 2.5 times; head shape relatively more massive and sharply delimited from the body; 7–9 precloacal pores in V-shaped line; not elongated limbs; 5–6 wide transversal dorsal bands and narrow interspaces, no contrasting nuchal loop, and wide transverse patterns. The main distinguishing features of the new species are the following: maximum SVL of 35.0 mm; maximum TL 40.2 mm; 8/8 infralabials; 6/7 supralabials; two pairs of small roundish postmentals, which contact mentals; one nasal scale; 22 scales between the orbits; 97–104 longitudinal ventral scales from postmentals to cloaca; 59–65 scales along the midline around the body; 11–12 subdigital lamellae on the fourth finger; 13–14 subdigital lamellae on the fourth toe; 2–3 cloacal spurs on each side; medial line of subcaudal scales formed by noticeably enlarged plates, 2–3 times larger the surrounding scales (Table 4).
Description of holotype. This is an adult male, SVL 35.0 mm, TL 40.2 mm, with a relatively large head shape (HL/SVL 0.28) that tapers into the neck and body. The rostrum is rounded and relatively elongate (Snyer/HL 0.31). The distance between rostral and occipital parts is approximately the same (SnEye/EyeEar 1.03). The ear opening is small and oval. The eye is relatively small (Orb D/HL 0.22). The distance between the posterior margin of eye and anterior margin of ear (EyeEar 2.9) exceeds the diameter of the orbit (Orb D 2.15).
The nostril is surrounded by the first supralabial, rostral, and one enlarged nasal scale. There are eight supralabials and six large infralabials rectangular in shape, which decrease in size posteriorly. A large mental shield with roundish caudal edge borders with the first labial and four small and rounded postmentals. There are 22 granular scales between the eyes. The upper part of the head is covered by rounded scales which are enlarged on the sides of rostrum and between the orbits. Large, rounded gulars gradually pass into ventral large flat smooth juxtaposed scales (Figure 9).
The body is elongated and slightly flattened, with no lateral folds. The dorsal scales are heterogenic. Among small roundish scales, evenly distributed enlarged dorsal tubercles do not form regular rows (Figure 9d). Enlarged dorsal tubercles are no more than 2.5 times larger than surrounding scales. In the longitudinal medial row from postmentales to cloaca, there are 97 scales, along with 65 scales around the midbody. There are eight precloacal pores, located on elongated rectangular precloacal scales in an inverted V-shaped row (Figure 9). The opening of the pore is large, occupying more than half of the total area of the precloacal scale. Between the row of precloacal pores and cloaca, there are five enlarged scales.
The forelimbs are slender, and the humeral length slightly exceeds the size of the forearm (ForeaL/LS 0.9). The hindlimbs are elongated, stronger in structure than the front. The thigh (6.5 mm) and lower leg are the same length. The dorsal surface of forelimbs and hindlimb is covered with smooth juxtaposed and imbricated scales, similar in size to the dorsal ones. The fingers are slender and not elongate (LD4A/SVL 0.07). From above, the metacarpus and metatarsus are covered with scales passing from the forearm and lower leg, respectively; those from below the palm and sole are covered with small swollen rounded granules, and the lower surface of the fingers is covered with transverse subdigital lamellae. The fourth digit of the hindlimb is the longest, with 13 slightly swollen subdigital lamellae.
The tail is 40.2 mm longer than the body (SVL/TL 0.9), cylindrical, rounded in cross-section, and not segmented. The base of the tail is slightly swollen; on the sides, there are two pairs of postcloacal spurs. The medial row features enlarged subcaudal plates.
Coloration. The background of the dorsum, head, and limbs are light beige (Figure 9d). The dorsal pattern is formed by six wide transversal dark brown bands with uneven edges; the interspaces between bands are approximately equal to or less than the width of the bands. There are 11 transverse dark bands on the tail. Patterns on the head are not pronounced, with a nuchal loop, dark brown strips on the sides of rostrum between eye, and supralabial plates. Moreover, no contrasting dark roundish spot is present on the tip of the rostrum. The dorsal surfaces of the limbs have transverse dark irregularly shaped spots (Figure 9). The color of the venter is white.
Distribution. Alsophylax emilia sp. nov. is known from a few localities on the northwestern edge of the Fergana Valley (Figure 2). Specimens were found at the three localities on pap foothills (adyrs): vicinity of Jidalisai N40.891349; E70.884367; vicinity of Uygursay (N40.95126; E71.01833); 10 km northwest of Turakurgan village (N41.067757; E71.412176); one locality in the Namangan region, vicinity of Yartepa vill. N 41.067757, E 71.412176. We hypothesize that the distribution of A. emilia sp. nov. is probably broader in the Fergana Valley, but future studies are needed to clarify the distribution of this species in the surrounding regions.
Habitat and natural history. The habitat at all known localities of Alsophylax emilia sp. nov. was similar and made of clay canyons with outcrops, with sparse vegetation consisting of various shrubs (Figure 8b).
Etymology. This species is named in honor of Soviet and then Uzbek herpetologist Emilia V. Vashetko (24.04.1940–07.11.2022) for her great contributions to the study of the herpetofauna of Uzbekistan and surrounding countries.
Comparison. Alsophylax emilia sp. nov. belongs to the group of Alsophylax with heterogeneous dorsal scales and enlarged dorsal tubercles, which includes A. loricatus, A. pipiens, A. przewalskii, and A. szczerbaki (Table 3). Alsophylax emilia sp. nov. is morphologically closest to A. pipiens, from which it can be distinguished by the smaller dorsal tubercles on new species, which is are no more than 2.5 times smaller than surrounding scales; the medial line of subcaudal scales is formed by noticeably enlarged plates, which are 2–3 times larger than surrounding scales; there are 7–9 precloacal pores versus 8–13 in A. pipiens (Figure 8 and Figure 9). Alsophylax emilia sp. nov. differs from A. loricatus, A. przewalskii, and A. szczerbaki by having smaller and smooth dorsal tubercles which are sporadically distributed across the back, versus large, strongly keeled trihedral dorsal tubercles organized in regular longitudinal rows; there are smaller scales on the head (22 scales between the eyes versus 8–15 in all three species mentioned above); dorsal patterns consist of transversal dark bands versus unclear dorsal patterns with longitudinal elements (Figure 6).
Alsophylax emilia sp. nov. further differs from A. tadjikiensis by the following characteristics: heterogeneous dorsal scalation versus homogeneous dorsal scalation; 7–9 precloacal pores in new species versus 5–7 pores in A. tadjikiensis; transversal dorsal bands versus pattern-less dorsum and bright yellowish or orange coloration of tail in A. tadjikiensis. Alsophylax emilia sp. nov. differs from A. ferganensis sp. nov. as follows: a larger body size with a maximum SVL of 35 mm versus 30.3 mm; heterogeneous dorsal scalation versus homogeneous dorsal scalation; elongate fore- and hindlimbs on A. ferganensis sp. nov.; transversal enlarged subcaudal plates versus roundish polygonal slightly enlarged subcaudal scales along the midline in A. ferganensis sp. nov. (Figure 10).

Figure 10.
A comparison of adult male Alsophylax ferganensis sp. nov. (top) and Alsophylax emilia sp. nov. (bottom).
4. Discussion
4.1. Taxonomy of Alsophylax and Conservation in the Fergana Valley
The genetic and morphological data in this study support the hypothesis that these new populations of Alsophylax represent undescribed new species, and that they currently appear to be endemic to the Fergana Valley of Uzbekistan (Figure 2 and Figure 10). Given that these new species tentatively belong to separate species groups within Alsophylax suggests that the Fergana Valley may have been colonized by at least two different Alsophylax lineages (Figure 1). These results further highlight the age and complex geological processes that have helped generate the unique biodiversity in this isolated valley. Additionally, these new species elevate the number of endemic reptiles to the Fergana Valley and Uzbekistan. Lastly, we provided data on population densities for all endemic reptile species found in the Fergana Valley (Table 1; Figures S1–S4) indicating that not all these species respond to habitat degradation in the same manner. We discuss these results in conjunction with the history of previous herpetological studies in the Fergana Valley, and the future of conservation and species discovery in the area.
4.2. The History of Herpetological Study in Central Asia and the Fergana Valley
The history of studies on reptiles inhabiting the territory of Uzbekistan, as was the case with other vertebrates in Central Asia, began with the expedition of E. A. Eversmann and K. Pander from Orenburg to Bukhara (October 1820–April 1821). The few and occasional collections of E. A. Eversmann from these areas were transferred to the University of Berlin, processed by Prof. M. Lichtenstein, and published in 1823 as an appendix to the work of E. A. Eversmann [30].
- −
- H. C. Lichtenstein published a list of 18 species of reptiles (including five new ones) and two species of amphibians [31].
- −
- A. Lehmann in July 1841 crossed the Kuvan-Darya and the Kyzylkum desert, and then visited Samarkand and Bukhara. From there, he traveled to the mountains to Iskenderun and went to Orenburg, Kazakhstan [32].
- −
- Brandt [33] and Nikolsky [34,35,36,37,38] expanded the known number of species in the area including two species of amphibians and 22 species of reptiles.
- −
- A. P. Fedchenko in 1871 worked in the Fergana Valley and visited Kokand, Andijon, Namangan, Vuadil, Uch-Kurgan, and Besh-Aryk. The works by Fedchenko [39,40,41] included only part of the materials he collected in Central Asia and the Fergana Valley. The remaining collections were mainly used by A. M. Nikolsky and A. A. Strauch in their works.
- −
- Strauch published a list of geckos from Central Asia, compiled their characteristics, and described a number of new species in Central Asia. Of greatest interest are descriptions of the Turkestan thin-toed gecko, Transcaspian bent-toed gecko, and Strauch’s even-fingered gecko from Uzbekistan and other republics of Central Asia [42].
- −
- Nikolsky published “Reptiles and amphibians of the Turkestan general governorship” and gave a complete literary summary at that time [34]. The author processed the materials of A. P. Fedchenko and others who collected in Central Asia. The paper provides information on the distribution of seven species of amphibians, three species of turtles, 42 species of lizards, and 28 species of snakes. Of these, more than half were collected in Uzbekistan [34].
- −
- Bogdanov published his work “Fauna of the Uzbek SSR. Amphibians and reptiles” [30] and provided all the data that he collected during his time doing field work in the Namangan region during 1949, 1950, 1954, and 1955.
- −
- E. V. Vashetko and Z. Kamalova studied racerunners and toad-head agamas living in the central region of the Fergana Valley [43,44,45].
- −
- T. Yadgarov studied snakes and the conservation of the endemic and unique ecosystems of the Fergana Valley [46,47]).
- −
- O.I. Tsaruk paid great attention to the problems and conservation of the unique ecosystems of the Fergana Valley, studying rare and endemic species of Phrynocephalus in the Fergana Valley [48].
- −
- Chikin (1998; 2001) assessed the status of populations of a number of species of rare and endemic reptiles in the sands of the Fergana Valley [49,50].
- −
- Bondarenko and Peregontsev (2017) described the spatial distribution of the Central Asian Tortoise, Testudo horsfieldii in Uzbekistan, including the Fergana Valley [51].
- −
- Currently, R.A. Nazarov and T.V. Abduraupov pay special attention to the study and conservation of the unique herpetofauna of the Fergana Valley, in particular, carrying out work on Papsky adyrs [4,52,53].
4.3. Conclusions
Lastly, the age of the Fergana Valley allows it to support subtle yet different habitats ranging from claystone outcroppings to massive, isolated sand massifs and xeric flats that each support their own suite of species that are locally adapted to these different micro-habitats. The discovery of possibly two new species (Figure 1 and Figure 3) that are micro-endemic habitat specialists isolated in various clay outcrops and canyons supports the hypothesis that the valley may have been colonized twice at different times. We hypothesize that this possible colonization pattern could indicate that other groups have experienced multiple colonization events and they too have levels of cryptic diversity among their populations. This study highlights the reptile diversity in the Fergana Valley and further supports the need for more surveys focusing on these habitats deemed “non-suitable”, because the data from this study indicate that they may harbor unrecognized biodiversity. If the biodiversity from these areas is not documented and protected, they could become victim to the continued textile and agricultural developments in the Fergana Valley.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13152516/s1, Figure S1. Distribution of Eremias scripta pherganensis. Figure S2. Distribution of Phrynocephalus strauchi. Figure S3. Distribution of Teratoscincus rustamowi. Figure S4. Top - Distribution of Alsophylax loricatus.
Author Contributions
Conceptualization and methodology, R.A.N., T.V.A. and J.L.G.; sample preparation, E.Y.S., M.A.G., R.A.N., D.A.M., J.D.L., M.D.B. and N.A.P.; molecular analysis, N.A.P.; data analysis, E.Y.S. and R.A.N.; manuscript preparation, R.A.N., E.Y.S., J.L.G. and M.D.B. All authors have read and agreed to the published version of the manuscript.
Funding
The initial 2019 expedition was funded by National Geographic Grant NGS 9535-14. This work was financially supported by the project of WWF01434/9E070711/GLO “Conservation of key natural complexes in the Fergana Valley (Republic of Uzbekistan)” of the Michael Succow Foundation, implemented with the support of WWF and CEPF (Critical Ecosystem Partnership Fund) and implementation of the research topic of the Institute of Zoology “Creating a scientific basis for maintaining cadaster of rare and endangered species of animals (on the example of the Ferghana Valley)”. This study was supported by the Russian Foundation of Basic Research project RFBR 20-54-56033 and under an international Iran-Russian project INSF 99003440. The molecular study was supported by the Russian Science Foundation (RSF grant No. 22-14-00037) with support of Russian Foundation for Basic Research grant No. 20-54-56033 (specimen collection, specimen examination) and frames of the State Themes of ZMMU 121032300105-0. Lastly, this research was partially funded by the State Themes of ZISP 122031100282-2 (awarded to author D.M.).
Institutional Review Board Statement
Surveys were conducted under the supervision of the State Committee of the Republic of Uzbekistan for Ecology and Environmental Protection.
Data Availability Statement
All data were deposited online.
Acknowledgments
The authors thank Ahmedov Askar, Soldatov Valentin, Adylova Dinara, Khasanov Mirpulat, Mun Yuriy, Bogatova Polina, and Chebotaryov Sergey for their assistance for collecting data in the field. They also thank the State Committee of the Republic of Uzbekistan for Ecology and Environmental Protection for their assistance in preparing permits for the collection of genetic samples.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Table A1.
A list of specimens which were used in morphological and phylogenetic analyses.
Table A1.
A list of specimens which were used in morphological and phylogenetic analyses.
| Species | Collection Number | N | E | H (m) | Locality |
|---|---|---|---|---|---|
| A. laevis | ZMMU R-6643 | 37.4139 | 65.0347 | 244 | Turkmenistan, Chardzhou region, eastern part of the Obruchev Steppe, vicinity of the Yusun well |
| ZMMU R-7280 1–2 | 40.697583 | 63.659622 | Uzbekistan, Bukhara region, foothills of Kuldzhuk-tau, 10 km west. Shuruk vill. | ||
| ZMMU R-4125 | 37.271453 | 60.245347 | Turkmenistan, Takhir RW station | ||
| ZMMU R-7281 1–2 | 40.546495 | 63.790836 | Uzbekistan, Bukhara region, 15 km south Shuruk vill. | ||
| ZMMU R-5895 | 38.046054 | 58.035237 | Turkmenistan, Gekdere vill. | ||
| ZMMU R-5940 | 39.213124 | 55.709714 | Turkmenistan, 10 km from Uzun-Su RW station | ||
| ZMMU R-16038 | 39.508916 | 54.362538 | Turkmenistan, 18 km northwest of Kyzyl Arvat (Sedar city), env. Qishlaq Aji | ||
| ZISP—18936 1–13 | 37.271453 | 60.245347 | Turkmenistan, station Takyr | ||
| ZISP—19177 1–15 | 37.371190 | 60.077820 | Turkmenistan, 33 km west of Tejen on the highway to Ashgabat | ||
| ZISP—16612 | 37.261158 | 67.307407 | Uzbekistan, Kara-su near Termez | ||
| ZISP—18925 1–27 | 39.015630 | 56.253282 | Turkmenistan, 4 km northwest of Kizyl-Arvat | ||
| IZUAS—1473 1–6 | 40.682777 | 64.010022 | Uzbekistan, 40 km to the west of Ayagitma (Kyzylkum) | ||
| IZUAS—1474 1–13 | 40.813600 | 64.336812 | Uzbekistan, 20 km to the northwest of Ayagitma | ||
| IZUAS—1574 1–2 | 40.682777 | 64.010022 | Uzbekistan, 20 km to the west of Ayagitma | ||
| A. loricatus | ZMMU R-10679 | 40.283611 | 69.623962 | Tajikistan, Leninabad, old fortress. | |
| ZMMU R-16041 | 40.289225 | 70.426283 | Tajikistan, Ferghana Valley, env. Kanibadam | ||
| ZMMU R-11424 1–5 | 40.319699 | 71.024406 | Uzbekistan, Ferghana Valley, env. Kokand city, Sarykurgan village on the Sokh river. Uzbek-Kyrgyz border. | ||
| ZMMU R-16105 1–6 | 40.535786 | 70.940439 | Uzbekistan, Ferghana Valley, environs of Kokand | ||
| ZMMU R-16106 1–3 | 40.331407 | 71.021404 | Uzbekistan, Fergana Valley, the bank of the Sokh River. Sarykurgan. | ||
| ZMMU R-16104 1–8 | 40.535786 | 70.940439 | Uzbekistan, Ferghana Valley, environs of Kokand | ||
| A. pipiens | ZMMU R-12915 | 44.05 | 69 | 337 | Kazakhstan, western part of the Moinkum Sands |
| ZMMU R-5878 1–9 | 42.757384 | 107.550913 | Mongolia, Galbyn-Gobi (southern), Tavan-Aldyn-Khudun | ||
| ZMMU R-5490 1–2 | 45.659435 | 91.154084 | Mongolia, Kobdos aimag, 15 km NW of the Ushig outpost | ||
| ZMMU R-6088 1–4 | 45.467223 | 106.260296 | Mongolia, Middle Gobi aimag, Bayan-Dov | ||
| ZMMU R-12602 | 44.140570 | 103.715861 | Mongolia, South Gobi aimag, Bayan-zag | ||
| ZMMU R-5881 1–3 | - | - | Mongolia | ||
| ZMMU R-13053 | 45.351402 | 96.647005 | Mongolia, Gobi-Altai aimag, Tsagan-Tolgoin-Bulak, about 30 km on the road from Tsogt to Dzakhoy | ||
| ZMMU R-4253 1–2 | 43.151441 | 109.145179 | Mongolia, Dornogob, 30 km south of Khuvsgul somon, on the road to Khatan-Bulak | ||
| ZMMU R-4924 1–3 | 43.229632 | 101.047568 | Mongolia, South Gobi aimag, vicinity of Gurvan-Tes | ||
| ZMMU R-5491 1–3 | 48.212680 | 102.217947 | Mongolia, Gobi-Altai aimag, Khatsovmiin-bulag, about 50 km S-SE from Khair-Khan | ||
| ZMMU R-5381 1–5 | 42.333333 | 103.25 | 1200 | Mongolia, South Gobi aimag, southern slope of the Mt. Baga-Ula | |
| ZMMU R-5489 | 43.128934 | 97.975124 | Mongolia, Bayan Khongor aimag, Shara-Khulsny-Bulak | ||
| ZMMU R-4920 1–4 | 43.251052 | 100.004418 | Mongolia, South Gobi aimag, Naryn-Bulak | ||
| ZMMU R-6090 | 42.533333 | 106.85 | Mongolia, 5 km to the west from Ganz-Mod-Khuduk | ||
| ZMMU R-5492 1–2 | 45.659435 | 91.154084 | Mongolia, Kobdos aimag, 10 km NW of the Ushig outpost | ||
| ZMMU R-5880 1–2 | 43.716667 | 108.633333 | Mongolia, Dornogovi aimag, in the ruins of a monastery | ||
| ZMMU R-6089 1–2 | 44.25 | 109.283333 | Mongolia, Dornogovi aimag | ||
| ZMMU R-5488 | 46.699222 | 91.069969 | Mongolia, Kobdos aimag, 22 km southwest of the Gurvan Khukhet outpost | ||
| ZMMU R-5254 | 43.667070 | 102.933058 | Mongolia, 150 km NW from Bayan-Leg (northern slope of the Nem-Get ridge) | ||
| ZMMU R-4921 | 43.321384 | 99.329913 | Mongolia, vicinity of the Ekhiin-Gol station | ||
| ZMMU R-5385 | 42.066667 | 104.416667 | 1250 | Mongolia, South Gobi aimag, 5 km NW from the Jal-Shand-Khuduk well | |
| ZMMU R-6056 | 43 | 107.666667 | Mongolia, Aimak Umnegovi | ||
| ZMMU R-7172 | 49.615853 | 116.993728 | China/Mongolia, South Gobi aimag, eastern tip of Bordzingiin-Gobi, 12 km west of Genghis Khan Wall | ||
| ZMMU R-3908 | 43.836195 | 106.549735 | Mongolia, South Gobi aimag, 10 km north of the southeastern tip of the Khurkhe-Ula ridge (an extension of the Gobi Altai) | ||
| ZMMU R-5829 25 | 48.967206 | 89.963372 | Mongolia, the ruins of the monastery Ulgii | ||
| ZMMU R-5382 1–3 | 41.833333 | 104.666667 | 1050 | Mongolia, South Gobi aimag, northern tip of the Dzamyn-Khuren-Eds sands, 10 km SE from the Bichit-Usny-Khuduk well | |
| ZMMU R-4922 | 43.251052 | 100.004418 | Mongolia, Naryn-Dats-Bulak | ||
| ZMMU R-4925 1–2 | 43.229632 | 101.047568 | Mongolia, South Gobi aimag, vicinity of Gurvan-Tes | ||
| ZMMU R-4923 317–318 | 43.909167 | 9.912222 | Mongolia, Haichin-2 | ||
| ZMMU R-12914 | 43.73 | 79.46 | Kazakhstan | ||
| ZMMU R-9571 | 44.052888 | 62.711996 | Kazakhstan, Kzyl-Orda region, 80 km to the northeast from Chaban-Kazgan, northern Kyzyl-Kum | ||
| ZMMU R-9570 1–6 | 43.535957 | 58.688738 | Uzbekistan, at the foot of the Chink near Sudachey Lake, in the lower reaches of the Amu-Darya | ||
| ZMMU R-14716 | 44.94747 | 68.57365 | Kazakhstan, the edge of the floodplain of the Chu River | ||
| ZMMU R-5920 | 43.423373 | 78.993863 | Kazakhstan, Alma-Ata region, 40 km W-SW Chundzha village | ||
| ZMMU R-336 | 45.657446 | 58.602648 | Kazakhstan, NW shore of the Aral Sea, Ust-Urt, Kaska-dzhol tract | ||
| ZMMU R-11423 1 | 43.870681 | 77.051736 | Kazakhstan, surroundings of Kapchagan | ||
| ZMMU R-115 1–3 | 45.915034 | 73.613855 | Kazakhstan, Cape Korzhun-Tubek, western shore of Lake Balkhash | ||
| ZMMU R-118 1–3 | 45.474858 | 73.612777 | Kazakhstan, Zhambyl region, Mynaral tract, western shore of the lake. Balkhash | ||
| ZMMU R-9569 1–2 | 5.252309 | 63.423455 | Kazakhstan, Northern Kyzyl-Kum, 50–70 km SW-W of Dzhusaly station | ||
| ZMMU R-9566 | 44.481389 | 64.168056 | Kazakhstan, Kum-Kala, Karmakchinsky district, Kzyl-Orda region | ||
| ZMMU R-9565 | 45.505092 | 61.612670 | Kazakhstan, Erembet-Tunbugut, Karmakchinsky district, Kzyl-Orda region | ||
| ZMMU R-9567 | 45.597886 | 63.072142 | Kazakhstan, 21 km W from Tunbugut, Karmakchinsky district, Kzyl-Orda region. | ||
| ZMMU R-12917 | 46.733333 | 75.366667 | Kazakhstan, north of Lake Balkhash, kishlak Ortaderessin | ||
| ZMMU R-13488 | 44.066667 | 76.983333 | Kazakhstan, Almaty region, about 18 km northwest of Kapchagay, at the foot of the rocks on the banks of the Ili River | ||
| ZMMU R-10682 1–2 | 50.430058 | 79.896721 | Kazakhstan, 25 km west of Semipalatinsk | ||
| ZMMU R-4293 1–9 | 50.423652 | 80.254928 | Kazakhstan, Semipalatinsk | ||
| ZMMU R-5896 | 40.45 | 54.5667 | Turkmenistan, Tersakkan tract, about 200 km NE from Krasnovodsk | ||
| ZMMU R-9564 1–2 | 43.616389 | 61.926667 | Kazakhstan, Western Kyzyl-Kum, Chaban Kazgan well | ||
| ZMMU R-10472 | 48.144205 | 46.856669 | Russia, Astrakhan region, Big Bogdo mountain | ||
| ZMMU R-9214 | 48.144205 | 46.856669 | Russia, Astrakhan region, Big Bogdo mountain | ||
| ZMMU R-10849 1–2 | 48.206567 | 46.970051 | Russia, Astrakhan region, Bogdinsky-Baskungaksky state reserve, steppe between Surikovskaya and Kordonnaya beams | ||
| ZMMU R-4621 | 48.144205 | 46.856669 | Russia, lower Volga, surroundings of Lake Baskunchak, Mount Bogdo. | ||
| ZMMU R-117 1–3 | 44.759161 | 74.432668 | Kazakhstan, Tai-Aral island in the south of Lake Balkhash. | ||
| ZMMU R-113 1–5 | 48.144205 | 46.856669 | Russia, Kyrgyz steppe (Big Bogdo) | ||
| ZMMU R-12568 | 44.95 | 63.616667 | Kazakhstan | ||
| ZMMU R-12569 | 45.605099 | 59.110612 | Kazakhstan, Aral Sea, Barsakelmes island | ||
| ZMMU R-6835 1–3 | 47.616760 | 80.420510 | Kazakhstan, Semipalatinsk region, right bank of the Ayaguz river, 40 km downstream from the city of Ayaguz | ||
| ZMMU R-4903 1–2 | 48.144205 | 46.856669 | Russia, lower Volga, surroundings of Lake Baskunchak, Mount Bogdo. | ||
| ZMMU R-111 | Expedition to the Persian Gulf | ||||
| ZMMU R-15912 | 43.681 | 108.441 | Mongolia, Dornogob aimag, Ungetiin uul | ||
| ZMMU R-15910 1–2 | 43.608 | 108.237 | Mongolia, Dornogob aimag, southeast of Suykhent | ||
| ZMMU R-15426 1–8 | 42.381003 | 57.370581 | Uzbekistan, Ustyurt plateau, northern shore of Sarykamysh lake | ||
| IZASU 120754—120759 | 42.997157 | 56.080082 | Uzbekistan, Ustyurt | ||
| A. przewalskii | ZMMU R-16103 | - | - | China | |
| ZISP 8271 | 37.354466 | 82.945839 | China, Niyadarya | ||
| ZISP 8250 | 37.110833 | 79.920833 | China, Chotan | ||
| ZISP 8267 | 35.810567 | 83.832288 | China, Kara-Sai | ||
| ZISP 15690 1–3 | 42.112944 | 92.112376 | China, coast from Bugas to Lukchun | ||
| ZISP 8258 1–3 | 41.531399 | 79.496704 | China, Chilan-su | ||
| A. szczerbaki | ZMMU R-12087 | 42.323907 | 59.180661 | Turkmenistan, Tashauz region, Kunya-Urgench | |
| ZISP 18885 7779–7784 | 41.650710 | 59.905444 | Turkmenistan, Takhta village on the Amur Darya | ||
| ZISP 18908 1870–1874 | 41.650710 | 59.905444 | Turkmenistan, Tatauzhskaya region, Takhta village | ||
| ZISP 18519 1–6 | 42.323393 | 59.162816 | Turkmenistan, the vicinity of the village of Kunya-Urgench | ||
| A. tadjikiensis | ZMMU R-16039 | - | - | Turkmenistan | |
| ZISP 18884 1824 | 37.595005 | 68.655830 | Tajikistan, Kurchantyubinsk region, 7 km from the village of Kyzyl-kakha down the Vakhsh river. | ||
| ZISP 18675 | 38.219290 | 69.126832 | Tajikistan, Valley of the Vakhsh River, near Kolkhozabad |
Appendix B
The Genbank numbers for the 47 samples used in the phylogenetic analyses:
| ZMMU_RAN_032_b_A_loricatus_COI_mtDNA | OR298052 |
| ZMMU_RAN_030_b_A_tokobajevi_COI_mtDNA | OR298053 |
| ZMMU_RAN_155_A_tokobajevi_COI_mtDNA | OR298054 |
| ZMMU_RAN_156_A_tokobajevi_COI_mtDNA | OR298055 |
| ZMMU_RAN_157_A_tokobajevi_COI_mtDNA | OR298056 |
| ZMMU_RAN_160_A_tokobajevi_COI_mtDNA | OR298057 |
| ZMMU_RAN_161_A_tokobajevi_COI_mtDNA | OR298058 |
| ZMMU_R_11384_1_A_tokobajevi_COI_mtDNA | OR298059 |
| ZMMU_R_11384_2_A_tokobajevi_COI_mtDNA | OR298060 |
| ZMMU_R_15426_1_A_pipiens_COI_mtDNA | OR298061 |
| ZMMU_R_15426_5_A_pipiens_COI_mtDNA | OR298062 |
| ZMMU_RAN_4932_laevis_COI_mtDNA | OR298063 |
| ZMMU_R_A_Fergana_COI_mtDNA | OR298064 |
| ZMMU_RAN_4917_emilia_COI_mtDNA | OR298065 |
| ZMMU_RAN_4918_emilia_COI_mtDNA | OR298066 |
| ZMMU_RAN_4924_emilia_COI_mtDNA | OR298067 |
| ZMMU_R_15426_2_A_pipiens_COI_mtDNA | OR298068 |
| ZMMU_R_15426_3_A_pipiens_COI_mtDNA | OR298069 |
| ZMMU_R_15426_4_A_pipiens_COI_mtDNA | OR298070 |
| ZMMU_RAN_2337a_A_przewalskii_COI_mtDNA | OR298071 |
| ZMMU_R_13488_A_pipiens_COI_mtDNA | OR298072 |
| ZMMU_RAN_031_b_A_pipiens_COI_mtDNA | OR298073 |
| ZMMU_R_12914_A_pipiens_COI_mtDNA | OR298074 |
| ZMMU_R_L_36_A_pipiens_COI_mtDNA | OR298075 |
| ZMMU_THD6_A_pipiens_COI_mtDNA | OR298076 |
| ZMMU_R_12915_1_A_pipiens_COI_mtDNA | OR298077 |
| ZMMU_R_12915_2_A_pipiens_COI_mtDNA | OR298078 |
| ZMMU_R_12815_A_pipiens_COI_mtDNA | OR298079 |
| ZMMU_RAN_1426_A_pipiens_COI_mtDNA | OR298080 |
| ZMMU_R_06_Mediodactylus | OR298081 |
| ZMMU_R_160381_laevis_2_COI_mtDNA | OR298082 |
| ZMMU_R_160382_laevis_2_COI_mtDNA | OR298083 |
| ZMMU_RAN_001_laevis_1_COI_mtDNA | OR298084 |
| ZMMU_RAN_002_laevis_1_COI_mtDNA | OR298085 |
| ZMMU_RAN_003_laevis_1_COI_mtDNA | OR298086 |
| ZMMU_R_160_tokobajevi_COI_mtDNA | OR298087 |
| ZMMU_RAN_004_tadjikiensis_COI_mtDNA | OR298088 |
| ZMMU_R_16039_tadjikiensis_COI_mtDNA | OR298089 |
| ZMMU_R_16040_loricatus_COI_mtDNA | OR298090 |
| ZMMU_R_16041_loricatus_COI_mtDNA | OR298091 |
| ZMMU_R_12816_pipiens_COI_mtDNA | OR298092 |
| ZMMU_R_15912_pipiens_COI_mtDNA | OR298093 |
| ZMMU_R_15910_1_pipiens_COI_mtDNA | OR298094 |
| ZMMU_R_15910_2_pipiens_COI_mtDNA | OR298095 |
References
- Sindaco, R.; Jeremcenko, V.K. The Reptiles of the Western Palearctic. In Annotated Checklist and Distributional Atlas of the Turtles, Crocodiles, Amphisbaenians and Lizards of Europe, North Africa, Middle East and Central Asia; Edizioni Belvedere: Latina, Italy, 2008; Volume 1, p. 580. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. 2022. Available online: https://www.iucnredlist.org (accessed on 2 July 2023).
- Bondarenko, K.A. Transformation of Socio-Cultural Factors Impacting on the external labour migration in Uzbekistan. Spat. Econ. Pros-Transtvennaya Ekon. 2020, 3, 76–108. [Google Scholar] [CrossRef]
- Nazarov, R.A.; Nuridjanov, D.A.; Vashetko, E.V. The current state of endemic reptilian species of the Fergana Valley and possible measures for their conservation. Contemporary Problems of Conservation of Rare, Disappearing Lowland Animals of Uzbekistan; Tashkent, Uzbekistan. 2016, pp. 141–149. Available online: https://istina.msu.ru/publications/article/32600194/ (accessed on 2 July 2023).
- Nazarov, R.A.; Radjabizadeh, M.; Poyarkov, N.A.; Ananjeva, N.B.; Melnikov, D.A.; Pouyani, E.R. A new species of frog-eyed gecko, genus Teratoscincus strauch, 1863 (Squamata: Sauria: Sphaerodactylidae), from central Iran. Russ. J. Herpetol. 2017, 24, 4. [Google Scholar] [CrossRef]
- Bykova, E.A.; Abduraupov, T.V.; Gritsina, M.A.; Esipov, A.V.; Dehkonov, S.I. Current status of rare and endangered species of vertebrates in Fergana region of Uzbekistan. In Actual Problems of Mathematics and Natural Sciences: Materials of the International Scientific-Practical Conference, Petropavlovsk-Barnaul-Surgut-Novosibirsk; Surgut State Pedagogical University: Surgut, Russia, 2022; pp. 257–263. [Google Scholar]
- Pallas, P. Zoographia Russo-Asiatics, Sistens Omnium Animalium in Extenso Imperio Rossico et Adjacentibus Maribus Observatorum Recensionem, Domicilia, Mores et Descriptiones, Anatomen Atque Icones Plurimorum; Caes. Academiae Scientarum: Petropoli, Russia, 1827; Volume 3, p. 426. [Google Scholar]
- Fitzinger, L.J.F.J. Systema Reptilium; Fasciculus Primus; Braumüller et Seidel: Wien, Austria, 1843. [Google Scholar]
- Szczerbak, N.N.; Golubev, M.L. The Gecko Fauna of the USSR and Adjacent Regions; Nauka Dymka: Kiev, Ukraine, 1986; p. 232. [Google Scholar]
- Anderson, S.C. The Lizards of Iran. Society for the Study of Amphibians and Reptiles. In Contributions to Herpetology; Ithaca: New York, NY, USA, 1999; Volume 15, p. 450. [Google Scholar]
- Khan, M.Z.; Mahmood, N. Current population status of diurnal lizards of Karachi, Pakistan. Russ. J. Herpetol. 2003, 10, 241–244. [Google Scholar]
- Bauer, A.M.; Masroor, R.; Titus-Mcquillan, J.; Heinicke, M.P.; Daza, J.D.; Jackman, T.R. A preliminary phylogeny of the Palearctic naked-toed geckos (Reptilia: Squamata: Gekkonidae) with taxonomic implications. Zootaxa 2013, 3599, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, I.; Bauer, A.M.; Jackman, T.R.; Karanth, P. Cryptic species and Miocene diversification of palaearctic naked-toed geckos (Squamata: Gekkonidae) in the Indian dry zone. Zool. Scr. 2014, 43, 455–471. [Google Scholar] [CrossRef]
- Gamble, T.; Greenbaum, E.; Jackman, T.R.; Bauer, A.M. Into the light: Diurnality has evolved multiple times in geckos. Biol. J. Linn. Soc. 2015, 115, 896–910. [Google Scholar] [CrossRef]
- Bondarenko, D.A.; Chelintsev, N.G. A comparative estimation of different methods of the line transect census of desert reptiles. Bulletin of Moscow Society of Naturalists. Biol. Ser. 1996, 101, 26–35. [Google Scholar]
- Bondarenko, D.A. “Raspredelenie i plotnost naseleniya sredneaziatskoy cherepahiv Tsentralnyh Kyzylkumah” [The Distribution and Population Density of the Central Asian Tortoise in Central Kizil-Kum (Uzbekistan)]. The Bulletin of Moscow Society of Naturalists. Biol. Ser. 1994, 99, 22–27. [Google Scholar]
- Bondarenko, D.A.; Peregontsev, E.A.; Mukhtar, G.B. Current state of steppe tortoise (Agrionemys horsfieldii Gray, 1844) populations in land-scapes of southern Kazakhstan. Russ. J. Ecol. 2008, 39, 210. [Google Scholar] [CrossRef]
- Bondarenko, D.A.; Peregontsev, E.A. Reptile communities of the Karakalpakian Ustyurt (Uzbekistan). Curr. Stud. Herpetol. 2018, 18, 13–26. [Google Scholar] [CrossRef]
- Hillis, D.M.; Moritz, C.; Mable, B.K. Molecular Systematics; Sinauer: Sunderland, MA, USA, 1996; Volume 2. [Google Scholar]
- Hebert, P.D.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Rodriguez, J.J.; Whitfield, J.B.; Deans, A.R.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D. Extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural history, DNA barcoding, morphology, and collections. Proc. Natl. Acad. Sci. USA 2008, 105, 12359–12364. [Google Scholar] [CrossRef] [PubMed]
- Solovyeva, E.N.; Poyarkov, N.A.; Dunaev, E.A.; Duysebayeva, T.N.; Bannikova, A.A. Molecular differentiation and taxonomy of the sunwatcher toad-headed agama species complex Phrynocephalus superspecies helioscopus (Pallas 1771) (Reptilia: Agamidae). Russ. J. Genet. 2011, 47, 842–856. [Google Scholar] [CrossRef]
- Nazarov, R.A.; Poyarkov, N.A.; Orlov, N.L.; Phung, T.M.; Nguyen, T.T.; Hoang, D.M.; Ziegler, T. Two new cryptic species of the Cyrtodactylus irregularis complex (Squamata: Gekkonidae) from southern Vietnam. Zootaxa 2012, 3302, 1–24. [Google Scholar] [CrossRef]
- Nagy, Z.T.; Sonet, G.; Glaw, F.; Vences, M. First large-scale DNA barcoding assessment of reptiles in the biodiversity hotspot of Madagascar, based on newly designed COI primers. PLoS ONE 2012, 7, e34506. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post–analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Chan, K.O.; Grismer, L.L. A standardized and statistically defensible framework for quantitative morphological analyses in taxonomic studies. Zootaxa 2021, 5023, 293–300. [Google Scholar] [CrossRef]
- Bogdanov, O.P. Fauna Uzbekskoj SSR. T. 1. Zemnovodnye i Presmykajuschiesya 1. [The Fauna of the Uzbek SSR. T. 1. Amphibians and Rep-Tiles]; Izdatelstvo Akademii Nauk Uzbekskoy SSR [Uzbek SSR Academy Sciences Publishing House]: Tashkent, Uzbekistan, 1960; p. 260. [Google Scholar]
- Lichtenstein, M.H.C. Verzeichniss der Doubletten des zoologischen Museums der Königl. In Universität zu Berlin nebst Beschreibung vieler bisher unbekannter Arten von Säugethieren, Vögeln, Amphibien und Fischen; Königl. Preuss. Akad. Wiss. T. Trautwein: Berlin, Germany, 1823; p. 118. [Google Scholar]
- Lehmann, A. Alexander Lehmann’s Reise nach Buchara und Samarkand in den Jahren 1841 und 1842 (Nach den hinterlas-senen Schriften desselben bearbeitet, und mit Anmerkungen versehen von G. v. Helmersen); Nebst einem zoologischen An-haenge von J.F.Brandt.; Kaiserlichen Akademie der Wissenschaften: Wien, Austria, 1852. [Google Scholar]
- Brandt, J.F. Zoologischer Anhang. In Alexander Lehmann’s Reise Nach Buchara und Samarkand in den Jahren 1841 und 1842; Beitr. z. Kenntn. d. Russ. Reiches; Kaiserlichen Akademie der Wissenschaften: Wien, Austria, 1852; p. 299. [Google Scholar]
- Nikolsky, A.M. Reptiles and amphibians of the Turkestan general-governorship (Herpetologia Turanica). AP Fedchenko, Travel to the Turkestan of the member-founder of the AP Fedchenko Society, accomplished on behalf of the Imperial Society of Amateurs of natural history, anthropology, and ethnography. Zoogeographical Investig. 1899, 23, 21–84. [Google Scholar]
- Nikolsky, A.M. Reptiles and amphibians of Russian Empire (Herpetologica Russica). Mem. De L’academie Imp. Des Sci. De St.-Petersbour 1905, 17, 518. [Google Scholar]
- Nikolsky, A.M. The Fauna of Russia and Adjacent Countries. Reptiles (Reptilia), Vol. 1. Chelonia and Sauria; Academia of Science: Petrograd, Russia, 1915; p. 534. [Google Scholar]
- Nikolsky, A.M. Fauna of Russia and Adjacent Countries. Reptiles (Reptilia) Vol. 2. Ophidia; Academia of Science: Petrograd, Russia, 1916; p. 247. [Google Scholar]
- Nikolsky, A.M. Fauna of Russia and Adjacent Countries. Amphibia; Academia of Science: Petrograd, Russia, 1918; p. 344. [Google Scholar]
- Fedchenko, A.P. Puteshestvie v Turkestan [Travels in Turkestan]. Turkestan Vedemosti 1870, 15, 91–92. [Google Scholar]
- Fedchenko, A.P. Report of the Turkestan scientific expedition of the society (from 1869 to April 15, 1870). Izv. Imp. Obs. Lubit. Estestvozn. Antropol. I Etnogr. 1871, 8, 135–190. [Google Scholar]
- Fedchenko, A.P. Voyage to Turkistan. CH. II. In Kokan Khanate; Izvestia Imperatorskogo O-va Lyubitelei Estestvoznania, Antropologii, i Etnografii: Moscow, Russia, 1875; Volume 7, p. 176. [Google Scholar]
- Strauch, A.A. Bemerkungen über die Geckoniden-Sammlung im zoologischen Museum der Kaiserlichen Akademie der Wissenschaften zu St. Petersburg. Mémoires De L’académie Impériale Des Sci. De St. Pétersbourg Ser. 1887, 7, 1–72. [Google Scholar]
- Vashetko, E.V. Ecology of rapid racerunner in Ferghana Valley. Zool. Zhurnal 1972, 51, 153–155. [Google Scholar]
- Vashetko, E.V.; Kamalova, Z.Y. Turtles, Lizards. In Pozvonochnye Zhivotnye Ferganskoi Doliny [Vertebrate Animals of the Ferghana Valley]; FAN Publ.: Tashkent, Uzbekistan, 1974; pp. 60–74. [Google Scholar]
- Vashetko, E.V.; Kamalova, Z.Y. On the Problem Creating a Reserve for the Conservation Endemic Reptile Species in the Ferghana Valley. In Okhrana Zhivotnogo Mira i Rastenii Uzbekistana: Tezisy Konferentsii [Conservation of the Flora and Fauna of Uzbekistan]; Fan Publ.: Tashkent, Uzbekistan, 1978; pp. 16–17. [Google Scholar]
- Yadgarov, T. Snakes. In Vertebrate Animals of Fergana Valley; FAN: Tashkent, Uzbekistan, 1974; pp. 75–84. [Google Scholar]
- Yadgarov, T.; Allabergenov, K. On conservation of gene pools of the vertebrate animals fauna of the Yazyavan Desert. Uzb. Biol. J. 1990, 2, 59–61. [Google Scholar]
- Tsaruk, O.I. Toad-heads agamas (Phrynocephalus) of Uzbekistan. The state of populations and prospects for conservation. In Reserves of Uzbekistan; Chinor ENK: Tashkent, Uzbekistan, 2001; Issue 3; pp. 72–77. [Google Scholar]
- Chikin, Y.A. Sohranenie Bioraznoobraziya Central’noy Azii, [Conservation of the Biodiversity in the Central Asia]; Zoohobbi: Alma-Ata, Kazakhstan, 1998; Volume 3. [Google Scholar]
- Chikin, Y.A. Protected reptile species from sands of Fergana. In Proceedings of the 1st meeting of the Nikolsky Herpetological Sociality, Puschino, Russia, 4–7 December 2000; pp. 327–328. [Google Scholar]
- Bondarenko, D.A.; Peregontsev, E.A. Distribution of the Central Asian Tortoise Agrionemys horsfieldii (Gray, 1844) in Uzbekistan (range, re-gional and landscape distribution, populations density). Curr. Stud. Herpetol. 2017, 17, 3–4, 124–146. [Google Scholar]
- Nazarov, R.A.; Abduraupov, T.V. New data on distribution and conservation of endemic reptile species in the Fergana Valley. In Proceedings of the Problems of Herpetology: Program and Abstracts of the VIII Congress of the A. M. Nikolsky Herpetological Society (NHS) of the Russian Academy of Sciences «Current Herpetological Research in Eurasia», Moscow, Russia, 3–9 October 2021; pp. 188–189. [Google Scholar]
- Abdulakimovna, M.M.; Narkuziyevich, A.O. Review of literature on Helmintofauna of Rodenta in the territory of the Republic of Uzbekistan. Gospod. I Innow. 2022, 21, 245–249. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).