
Spatiotemporal and Individual Patterns of Domestic Cat (Felis catus) Hunting Behaviour in France
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
- Following Thomas et al. [28] in the UK, we predict that the number of prey brought home by domestic cats will be higher during spring and summer coinciding with prey breeding seasons in temperate areas than during the rest of the year.
- Weather conditions strongly influence small mammals [29,30,31], birds [32,33], and lacertid [34] activity in Europe; rainfall positively influences small mammal activity but negatively influences the activity of birds and lacertids. Thus, we predict a higher number of small mammals brought home by cats in regions with a higher relative rainfall, while the number of birds and lacertids brought home by cats in such localities will be lower.
- Biogeographic factors determine prey species’ ecological range [35,36]. Accordingly, we predict that the number of individuals of endothermic prey (i.e., mammals and birds) brought home by cats will increase from the southeast to the northwest while the opposite pattern will be true for ectothermic prey (i.e., lacertids) brought home by cats.
2. Materials and Methods
2.1. Preys Brought Home
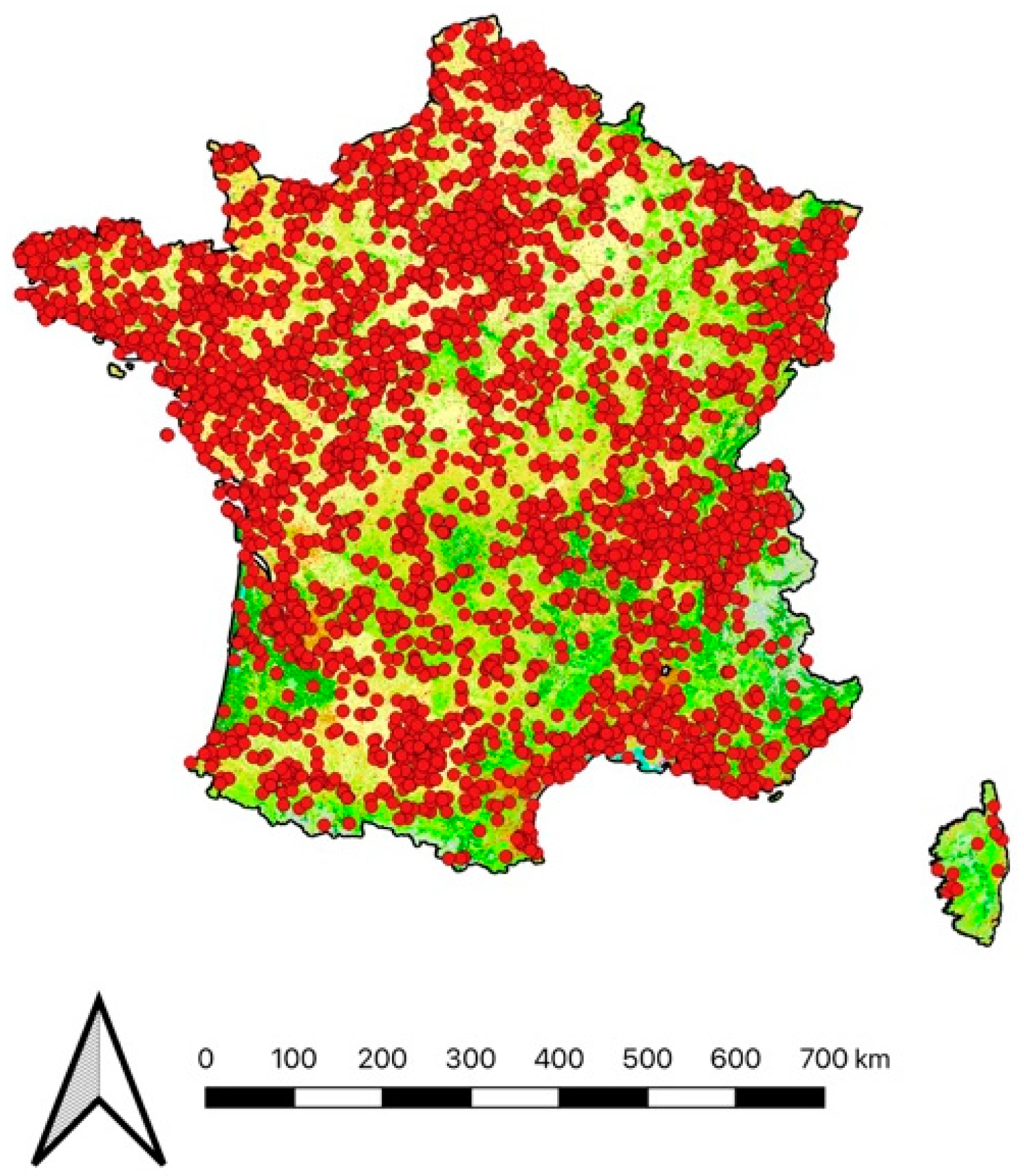
2.2. Spatial Variables
2.3. Statistical Analysis
3. Results
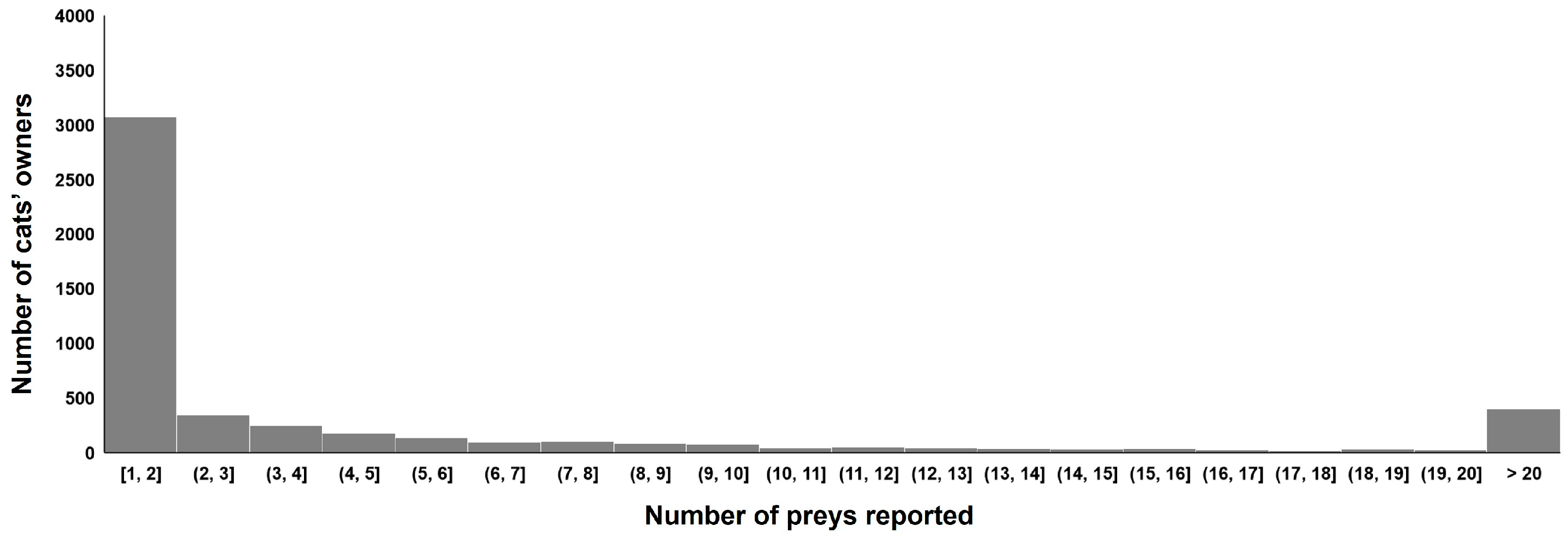
3.1. Prey Species Brought Home by Cats
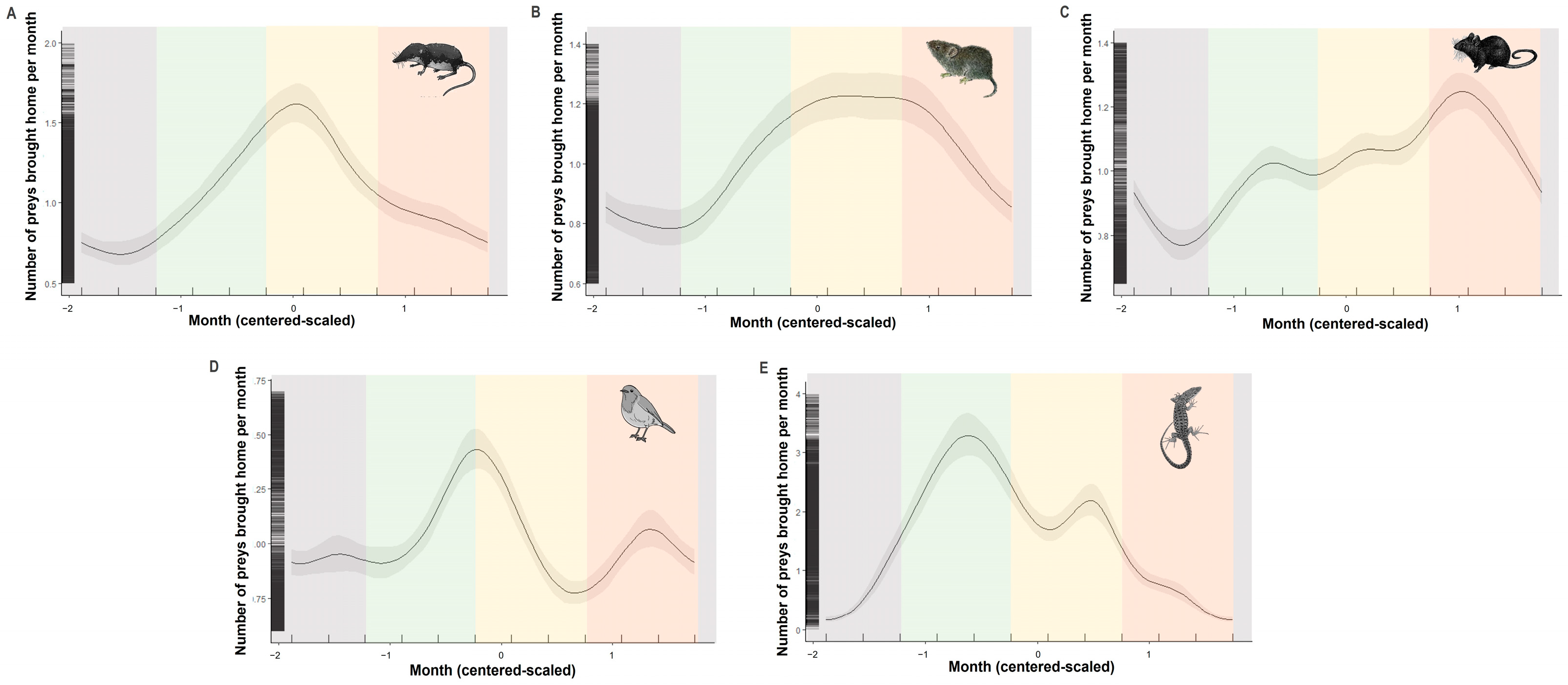
3.2. Seasonality of Prey Brought Home by Cats
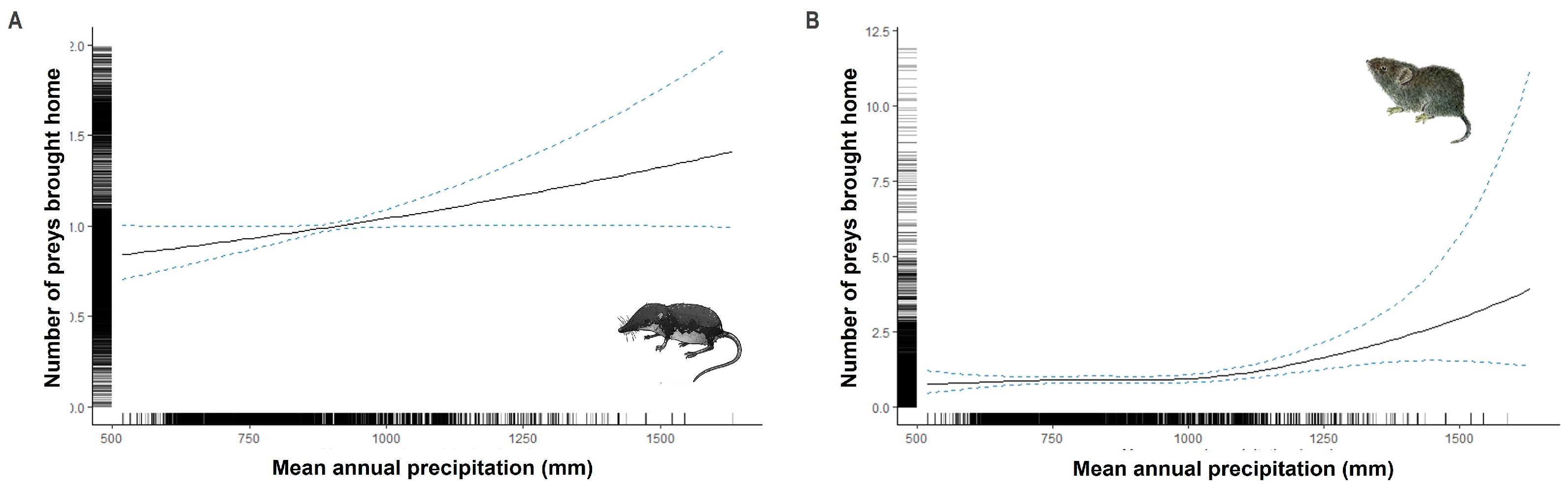
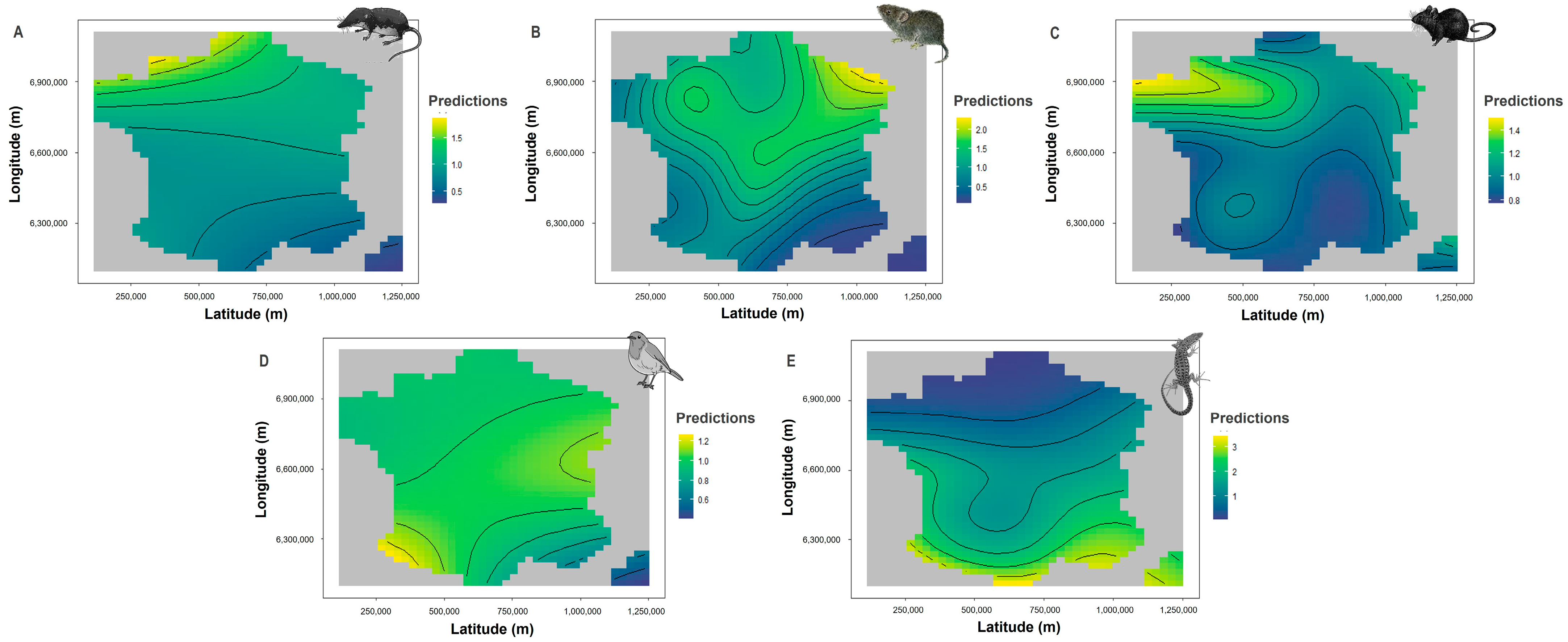
3.3. Climatic and Geographic Effects on Prey Brought Home by Cats
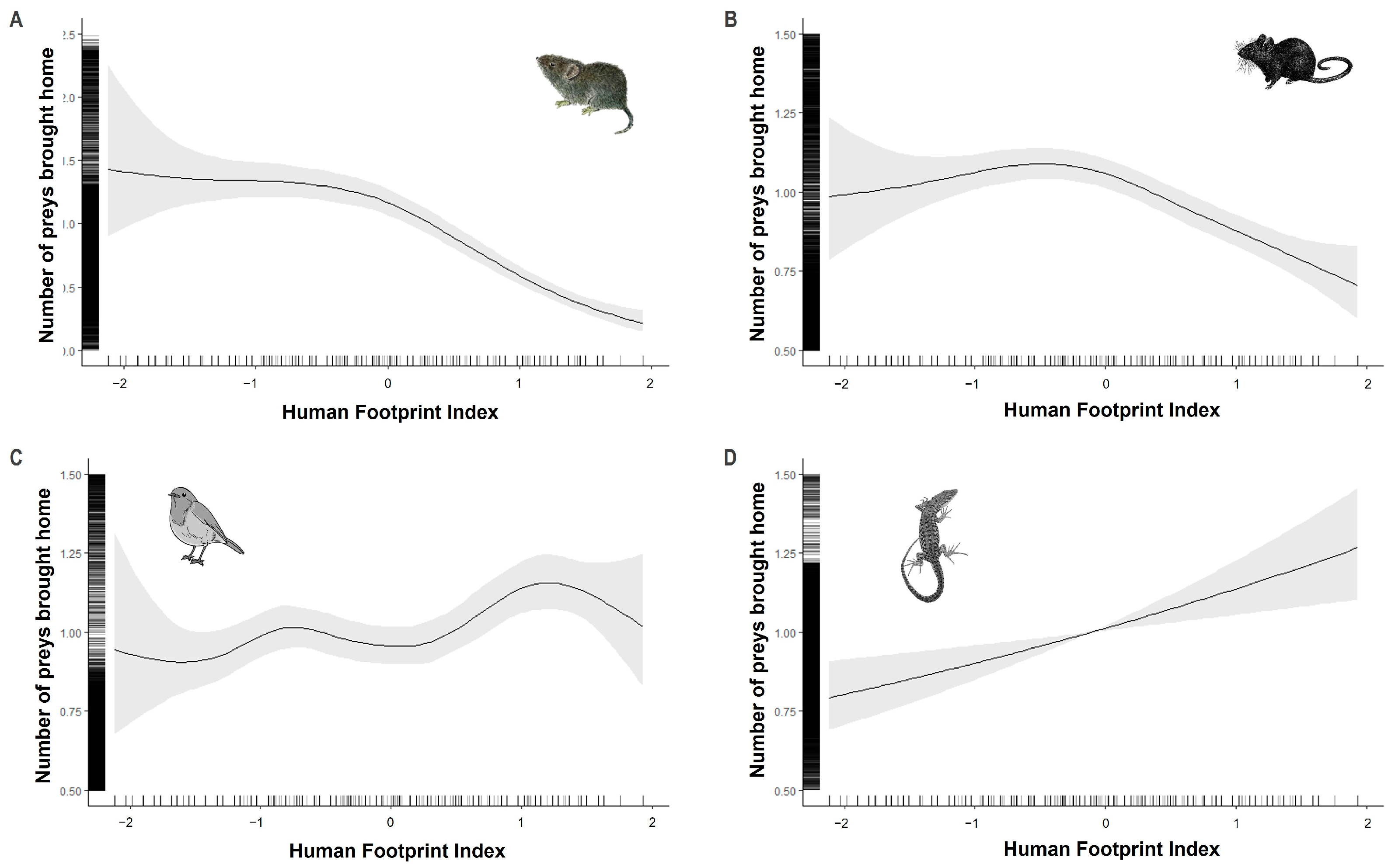
3.4. Anthropogenic Impacts on Prey Brought Home by Cats
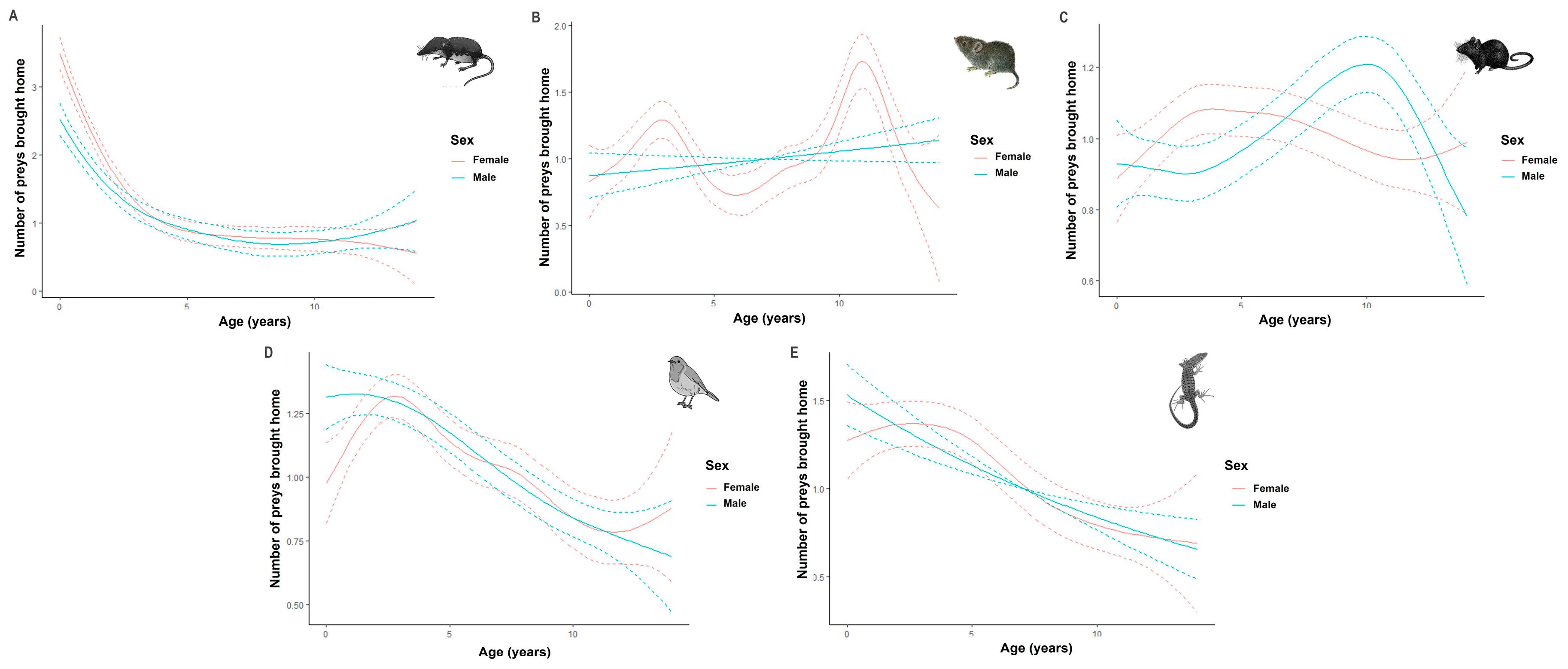
3.5. Individual Factors Influencing Prey Brought Home by Cats
4. Discussion
4.1. Prey Species Brought Home by Cats
4.2. Seasonality of Prey Brought Home by Cats
4.3. Climatic and Geographic Effects on Prey Brought Home by Cats
4.4. Anthropogenic Pressure Influence on Prey Returned Home
4.5. Individual Factors Influencing Prey Brought Home by Cats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Driscoll, C.A.; Clutton-Brock, J.; Kitchener, A.C.; O’Brien, S.J. The Taming of the Cat. Sci. Am. 2009, 300, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Genovesi, P.; Jeschke, J.M. Global Patterns in Threats to Vertebrates by Biological Invasions. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152454. [Google Scholar] [CrossRef] [PubMed]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group Auckland: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Anonymous Annual Facts & Figures. Available online: https://europeanpetfood.org/about/statistics/ (accessed on 28 June 2023).
- Mori, E.; Menchetti, M.; Camporesi, A.; Cavigioli, L.; Tabarelli de Fatis, K.; Girardello, M. License to Kill? Domestic Cats Affect a Wide Range of Native Fauna in a Highly Biodiverse Mediterranean Country. Front. Ecol. Evol. 2019, 7, 477. [Google Scholar] [CrossRef]
- Crowley, S.L.; Cecchetti, M.; McDonald, R.A. Hunting Behaviour in Domestic Cats: An Exploratory Study of Risk and Responsibility among Cat Owners. People Nat. 2019, 1, 18–30. [Google Scholar] [CrossRef]
- Forin-Wiart, M.-A. Identification des Facteurs de Variation de la Prédation Exercée par les Chats Domestiques (Felis silvestris catus) En Milieu Rural. Ph.D. Thesis, Ecole Doctorale Sciences, Technologies, Santé, Reims, France, 2014. [Google Scholar]
- Trouwborst, A.; McCormack, P.C.; Martínez Camacho, E. Domestic Cats and Their Impacts on Biodiversity: A Blind Spot in the Application of Nature Conservation Law. People Nat. 2020, 2, 235–250. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. The Impact of Free-Ranging Domestic Cats on Wildlife of the United States. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Loss, S.R.; Marra, P.P. Population Impacts of Free-Ranging Domestic Cats on Mainland Vertebrates. Front. Ecol. Environ. 2017, 15, 502–509. [Google Scholar] [CrossRef]
- Murphy, B.P.; Woolley, L.-A.; Geyle, H.M.; Legge, S.M.; Palmer, R.; Dickman, C.R.; Augusteyn, J.; Brown, S.C.; Comer, S.; Doherty, T.S. Introduced Cats (Felis catus) Eating a Continental Fauna: The Number of Mammals Killed in Australia. Biol. Conserv. 2019, 237, 28–40. [Google Scholar] [CrossRef]
- Li, Y.; Wan, Y.; Shen, H.; Loss, S.R.; Marra, P.P.; Li, Z. Estimates of Wildlife Killed by Free-Ranging Cats in China. Biol. Conserv. 2021, 253, 108929. [Google Scholar] [CrossRef]
- Blancher, P. Estimated Number of Birds Killed by House Cats (Felis catus) in Canada Estimation du Nombre d’oiseaux Tués par les Chats Domestiques (Felis catus) Au Canada. Avian Conserv. Ecol. 2013, 8, 3. [Google Scholar]
- Eisenhauer, N. Impacts of Free-Ranging Cats on Invertebrates. Front. Ecol. Environ. 2018, 16, 262–263. [Google Scholar] [CrossRef]
- Calvert, A.; Bishop, C.; Elliot, R.; Krebs, E.; Kydd, T.; Machtans, C.; Robertson, G. A Synthesis of Human-Related Avian Mortality in Canada. Avian Conserv. Ecol. 2013, 8, 11. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. Direct Mortality of Birds from Anthropogenic Causes. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 99–120. [Google Scholar] [CrossRef]
- Woods, M.; McDonald, R.A.; Harris, S. Predation of Wildlife by Domestic Cats Felis catus in Great Britain. Mammal Rev. 2003, 33, 174–188. [Google Scholar] [CrossRef]
- Tschanz, B.; Hegglin, D.; Gloor, S.; Bontadina, F. Hunters and Non-Hunters: Skewed Predation Rate by Domestic Cats in a Rural Village. Eur. J. Wildl. Res. 2011, 57, 597–602. [Google Scholar] [CrossRef]
- Meek, P.D. Food Items Brought Home by Domestic Cats Felis catus (L.) Living in Booderee National Park, Jervis Bay. Proc. Linn. Soc. N. S. W. 1998, 120, 43–48. [Google Scholar]
- Morgan, S.A.; Hansen, C.M.; Ross, J.G.; Hickling, G.J.; Ogilvie, S.C.; Paterson, A.M. Urban Cat (Felis catus) Movement and Predation Activity Associated with a Wetland Reserve in New Zealand. Wildl. Res. 2009, 36, 574–580. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Żmihorski, M.; Gryz, J. Annual Variation in Prey Composition of Domestic Cats in Rural and Urban Environment. Urban Ecosyst. 2017, 20, 945–952. [Google Scholar] [CrossRef]
- Gillies, C.; Clout, M. The Prey of Domestic Cats (Felis catus) in Two Suburbs of Auckland City, New Zealand. J. Zool. 2003, 259, 309–315. [Google Scholar] [CrossRef]
- Baker, P.J.; Bentley, A.J.; Ansell, R.J.; Harris, S. Impact of Predation by Domestic Cats Felis catus in an Urban Area. Mammal Rev. 2005, 35, 302–312. [Google Scholar] [CrossRef]
- Dickman, C.R.; Newsome, T.M. Individual Hunting Behaviour and Prey Specialisation in the House Cat Felis catus: Implications for Conservation and Management. Appl. Anim. Behav. Sci. 2015, 173, 76–87. [Google Scholar] [CrossRef]
- Loyd, K.A.T.; Hernandez, S.M.; Carroll, J.P.; Abernathy, K.J.; Marshall, G.J. Quantifying Free-Roaming Domestic Cat Predation Using Animal-Borne Video Cameras. Biol. Conserv. 2013, 160, 183–189. [Google Scholar] [CrossRef]
- Seymour, C.L.; Simmons, R.E.; Morling, F.; George, S.T.; Peters, K.; O’riain, M.J. Caught on Camera: The Impacts of Urban Domestic Cats on Wild Prey in an African City and Neighbouring Protected Areas. Glob. Ecol. Conserv. 2020, 23, e01198. [Google Scholar] [CrossRef]
- Brown, E.D.; Williams, B.K. The Potential for Citizen Science to Produce Reliable and Useful Information in Ecology. Conserv. Biol. 2019, 33, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.L.; Fellowes, M.D.; Baker, P.J. Spatio-Temporal Variation in Predation by Urban Domestic Cats (Felis catus) and the Acceptability of Possible Management Actions in the UK. PLoS ONE 2012, 7, e49369. [Google Scholar] [CrossRef]
- Wróbel, A.; Bogdziewicz, M. It Is Raining Mice and Voles: Which Weather Conditions Influence the Activity of Apodemus flavicollis and Myodes glareolus? Eur. J. Wildl. Res. 2015, 61, 475–478. [Google Scholar] [CrossRef]
- Sidorowicz, J. Influence of the Weather on Capture of Micromammalia. I. Rodents (Rodentia). Acta Theriol. 1960, 4, 139–158. [Google Scholar] [CrossRef]
- Vickery, W.L.; Bider, J.R. The Influence of Weather on Rodent Activity. J. Mammal. 1981, 62, 140–145. [Google Scholar] [CrossRef]
- O’Connor, R.J.; Hicks, R.K. The Influence of Weather Conditions on the Detection of Birds during Common Birds Census Fieldwork. Bird Study 1980, 27, 137–151. [Google Scholar] [CrossRef]
- Robbins, C.S. Bird Activity Levels Related to Weather. Stud. Avian Biol. 1981, 6, 301–310. [Google Scholar]
- Ortega, Z.; Pérez-Mellado, V. Seasonal Patterns of Body Temperature and Microhabitat Selection in a Lacertid Lizard. Acta Oecologica 2016, 77, 201–206. [Google Scholar] [CrossRef]
- Fitzgerald, M. Hunting Behaviour of Domestic Cats and Their Impact on Prey Populations. In The Domestic Cats: The Biology of Its Behaviour; Turner, D.C., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 151–175. [Google Scholar]
- Lozano, J.; Moleón, M.; Virgós, E. Biogeographical Patterns in the Diet of the Wildcat, Felis silvestris Schreber, in Eurasia: Factors Affecting the Trophic Diversity. J. Biogeogr. 2006, 33, 1076–1085. [Google Scholar] [CrossRef]
- Baker, P.J.; Molony, S.E.; Stone, E.; Cuthill, I.C.; Harris, S. Cats about Town: Is Predation by Free-Ranging Pet Cats Felis catus Likely to Affect Urban Bird Populations? Ibis 2008, 150, 86–99. [Google Scholar] [CrossRef]
- Cordonnier, M.; Ferry, N.; Renaud, E.; Maurice, A.-C.; Bonnaud, E.; Baudry, E. Drivers of Predation by Pet Cats: Environment Overcomes Predator’s Intrinsic Characteristics. Urban Ecosyst. 2022, 25, 1327–1337. [Google Scholar] [CrossRef]
- Kauhala, K.; Talvitie, K.; Vuorisalo, T. Free-Ranging House Cats in Urban and Rural Areas in the North: Useful Rodent Killers or Harmful Bird Predators? Folia Zool. 2015, 64, 45–55. [Google Scholar] [CrossRef]
- Kays, R.; Dunn, R.R.; Parsons, A.W.; Mcdonald, B.; Perkins, T.; Powers, S.A.; Shell, L.; McDonald, J.L.; Cole, H.; Kikillus, H. The Small Home Ranges and Large Local Ecological Impacts of Pet Cats. Anim. Conserv. 2020, 23, 516–523. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin/Heidelberg, Germany, 2009; Volume 574. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A Protocol for Data Exploration to Avoid Common Statistical Problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Wood, S.N. Mgcv: GAMs with GCV Smoothness Estimation and GAMMs by REML/PQL, R Package Version 1.1-8; R Foundation for Statistical Computing: Vienna, Austria, 2004; Available online: https://www.r-project.org/ (accessed on 6 February 2023).
- Fasiolo, M.; Nedellec, R.; Goude, Y.; Wood, S.N. Scalable Visualization Methods for Modern Generalized Additive Models. J. Comput. Graph. Stat. 2020, 29, 78–86. [Google Scholar] [CrossRef]
- Barratt, D.G. Predation and Movement by House-Based Domestic Cats Felis catus (L.) in Suburban and Rural Habitats—Preliminary Findings. In People and Nature Conservation: Perspectives on Private Land Use and Endangered Species Recovery; Royal Zoological Society of New South Wales: Mosman, NSW, Australia, 1995; pp. 181–187. [Google Scholar]
- Barratt, D.G. Predation by House Cats, Felis catus (L.), in Canberra, Australia. I. Prey Composition and Preference. Wildl. Res. 1997, 24, 263–277. [Google Scholar] [CrossRef]
- Driscoll, C.A.; Macdonald, D.W.; O’Brien, S.J. From Wild Animals to Domestic Pets, an Evolutionary View of Domestication. Proc. Natl. Acad. Sci. USA 2009, 106, 9971–9978. [Google Scholar] [CrossRef] [PubMed]
- van Heezik, Y.; Smyth, A.; Adams, A.; Gordon, J. Do Domestic Cats Impose an Unsustainable Harvest on Urban Bird Populations? Biol. Conserv. 2010, 143, 121–130. [Google Scholar] [CrossRef]
- Ancillotto, L.; Serangeli, M.T.; Russo, D. Curiosity Killed the Bat: Domestic Cats as Bat Predators. Mamm. Biol. 2013, 78, 369–373. [Google Scholar] [CrossRef]
- Oedin, M.; Brescia, F.; Millon, A.; Murphy, B.P.; Palmas, P.; Woinarski, J.C.; Vidal, E. Cats Felis catus as a Threat to Bats Worldwide: A Review of the Evidence. Mammal Rev. 2021, 51, 323–337. [Google Scholar] [CrossRef]
- Pisanu, B.; Pavisse, R.; Clergeau, P. GPS-Based Seasonal Home Ranges of Neutered Pet Cats Felis catus along a Habitat Gradient. Hystrix Ital. J. Mammal. 2020, 31, 105–109. [Google Scholar]
- Ferreira, J.P.; Leitão, I.; Santos-Reis, M.; Revilla, E. Human-Related Factors Regulate the Spatial Ecology of Domestic Cats in Sensitiveareas for Conservation. PLoS ONE 2011, 6, e25970. [Google Scholar] [CrossRef]
- Germain, E.; Benhamou, S.; Poulle, M.-L. Spatio-Temporal Sharing between the European Wildcat, the Domestic Cat and Their Hybrids. J. Zool. 2008, 276, 195–203. [Google Scholar] [CrossRef]
- Goszczyński, J.; Krauze, D.; Gryz, J. Activity and Exploration Range of House Cats in Rural Areas of Central Poland. Folia Zool. 2009, 58, 363–371. [Google Scholar]
- Churchfield, S. Population Dynamics and the Seasonal Fluctuations in Numbers of the Common Shrew in Britain. Acta Theriol. 1980, 25, 415–424. [Google Scholar] [CrossRef]
- Churchfield, S.; Hollier, J.; Brown, V.K. Population Dynamics and Survivorship Patterns in the Common Shrew Sorex araneus in Southern England. Acta Theriol. 1995, 40, 53–68. [Google Scholar] [CrossRef]
- Henttonen, H.; Haukisalmi, V.; Kaikusalo, A.; Korpimäki, E.; Norrdahl, K.; Skarén, U.A. Long-Term Population Dynamics of the Common Shrew Sorex araneus in Finland. Ann. Zool. Fenn. 1989, 26, 349–355. [Google Scholar]
- Torre, I.; Bastardas-Llabot, J.; Arrizabalaga, A.; Díaz, M. Population Dynamics of Small Endotherms under Global Change: Greater White-Toothed Shrews Crocidura russula in Mediterranean Habitats. Sci. Total Environ. 2020, 705, 135799. [Google Scholar] [CrossRef] [PubMed]
- Pucek, Z.; Jędrzejewski, W.; Jędrzejewska, B.; Pucek, M. Rodent Population Dynamics in a Primeval Deciduous Forest (Białowieża National Park) in Relation to Weather, Seed Crop, and Predation. Acta Theriol. 1993, 38, 199–232. [Google Scholar] [CrossRef]
- Ylönen, H.; Altner, H.-J.; Stubbe, M. Seasonal Dynamics of Small Mammals in an Isolated Woodlot and Its Agricultural Surroundings. Ann. Zool. Fenn. 1991, 28, 7–14. [Google Scholar]
- Maher, C.R.; Lott, D.F. A Review of Ecological Determinants of Territoriality within Vertebrate Species. Am. Midl. Nat. 2000, 143, 1–29. [Google Scholar] [CrossRef]
- Meek, R. Patterns of Reptile Road-Kills in the Vendée Region of Western France. Herpetol. J. 2009, 19, 135–142. [Google Scholar]
- Ferreiro, R.; Galán, P. Reproductive Ecology of the Slow Worm (Anguis Fragilis) in the Northwest Iberian Peninsula. Anim. Biol. 2004, 54, 353–371. [Google Scholar] [CrossRef]
- Pilorge, T. Density, Size Structure, and Reproductive Characteristics of Three Populations of Lacerta vivipara (Sauria: Lacertidae). Herpetologica 1987, 43, 345–356. [Google Scholar]
- Koenig, J.; Shine, R.; Shea, G. The Dangers of Life in the City: Patterns of Activity, Injury and Mortality in Suburban Lizards (Tiliqua scincoides). J. Herpetol. 2002, 36, 62–68. [Google Scholar] [CrossRef]
- Esther, A.; Imholt, C.; Perner, J.; Schumacher, J.; Jacob, J. Correlations between Weather Conditions and Common Vole (Microtus Arvalis) Densities Identified by Regression Tree Analysis. Basic Appl. Ecol. 2014, 15, 75–84. [Google Scholar] [CrossRef]
- Bretagnolle, V.; Gillis, H. Predator–Prey Interactions and Climate Change. In Effects of Climate Change on Birds; Oxford University Press: Oxford, UK, 2010; pp. 227–248. [Google Scholar]
- Gilg, O.; Sittler, B.; Hanski, I. Climate Change and Cyclic Predator–Prey Population Dynamics in the High Arctic. Glob. Chang. Biol. 2009, 15, 2634–2652. [Google Scholar] [CrossRef]
- Gilg, O.; Kovacs, K.M.; Aars, J.; Fort, J.; Gauthier, G.; Grémillet, D.; Ims, R.A.; Meltofte, H.; Moreau, J.; Post, E. Climate Change and the Ecology and Evolution of Arctic Vertebrates. Ann. N. Y. Acad. Sci. 2012, 1249, 166–190. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, G.; Giraudoux, P.; Delattre, P. Numerical and Dietary Responses of a Predator Community in a Temperate Zone of Europe. Ecography 2009, 32, 277–290. [Google Scholar] [CrossRef]
- Weber, J.-M.; Dailly, L. Food Habits and Ranging Behaviour of a Group of Farm Cats (Felis catus) in a Swiss Mountainous Area. J. Zool. Lond. 1998, 245, 234–237. [Google Scholar] [CrossRef]
- Liberg, O. Food Habits and Prey Impact by Feral and House-Based Domestic Cats in a Rural Area in Southern Sweden. J. Mammal. 1984, 65, 424–432. [Google Scholar] [CrossRef]
- Delattre, P.; Giraudoux, P.; Baudry, J.; Quéré, J.P.; Fichet, E. Effect of Landscape Structure on Common Vole (Microtus arvalis) Distribution and Abundance at Several Space Scales. Landsc. Ecol. 1996, 11, 279–288. [Google Scholar] [CrossRef]
- Bonnaud, E.; Bourgeois, K.; Vidal, E.; Legrand, J.; Le Corre, M. How Can the Yelkouan Shearwater Survive Feral Cat Predation? A Meta-Population Structure as a Solution? Popul. Ecol. 2009, 51, 261–270. [Google Scholar] [CrossRef]
- Neves, T.; Tapisso, J.T.; Porto, M.; Pereira, H.M.; Mathias, M.L.; Borda-de-Água, L. The Role of Competition in Driving Species Global Distributions: Soricid Shrews as a Case Study. J. Biogeogr. 2019, 46, 134–144. [Google Scholar] [CrossRef]
- Carretero, M.A. From Set Menu to a La Carte. Linking Issues in Trophic Ecology of Mediterranean Lacertids. Ital. J. Zool. 2004, 71, 121–133. [Google Scholar] [CrossRef]
- Graux, A.-I.; Resmond, R.; Casellas, E.; Delaby, L.; Faverdin, P.; Le Bas, C.; Ripoche, D.; Ruget, F.; Therond, O.; Vertès, F. High-Resolution Assessment of French Grassland Dry Matter and Nitrogen Yields. Eur. J. Agron. 2020, 112, 125952. [Google Scholar] [CrossRef]
- Guislain, M.-H.; Raoul, F.; Giraudoux, P.; Terrier, M.-E.; Froment, G.; Ferté, H.; Poulle, M.-L. Ecological and Biological Factors Involved in the Transmission of Echinococcus multilocularis in the French Ardennes. J. Helminthol. 2008, 82, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Montemayor, E.; Ferryman, M.; Watts, K.; Macgregor, N.A.; Hambly, N.; Brennan, S.; Coxon, R.; Langridge, H.; Park, K.J. Small Mammal Responses to Long-Term Large-Scale Woodland Creation: The Influence of Local and Landscape-Level Attributes. Ecol. Appl. 2020, 30, e02028. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewski, R.; Babińska-Werka, J.; Gliwicz, J.; Goszczyński, J. Synurbization Processes in Population of Apodemus agrarius. I. Characteristics of Populations in an Urbanization Gradient. Acta Theriol. 1978, 23, 341–358. [Google Scholar] [CrossRef]
- Cavia, R.; Cueto, G.R.; Suárez, O.V. Changes in Rodent Communities According to the Landscape Structure in an Urban Ecosystem. Landsc. Urban Plan. 2009, 90, 11–19. [Google Scholar] [CrossRef]
- Gomes, V.; Ribeiro, R.; Carretero, M.A. Effects of Urban Habitat Fragmentation on Common Small Mammals: Species versus Communities. Biodivers. Conserv. 2011, 20, 3577–3590. [Google Scholar] [CrossRef]
- Gortat, T.; Barkowska, M.; Gryczyńska-Siemią Tkowska, A.; Pieniążek, A.; Kozakiewicz, A.; Kozakiewicz, M. The Effects of Urbanization—Small Mammal Communities in a Gradient of Human Pressure in Warsaw City, Poland. Pol. J. Ecol. 2014, 62, 163–172. [Google Scholar] [CrossRef]
- Klimant, P.; Klimantová, A.; Baláž, I.; Jakab, I.; Tulis, F.; Rybanský, Ľ.; Vadel, Ľ.; Krumpálová, Z. Small Mammals in an Urban Area: Habitat Preferences and Urban-Rural Gradient in Nitra City, Slovakia. Pol. J. Ecol. 2017, 65, 144–157. [Google Scholar] [CrossRef]
- Łopucki, R.; Kitowski, I. How Small Cities Affect the Biodiversity of Ground-Dwelling Mammals and the Relevance of This Knowledge in Planning Urban Land Expansion in Terms of Urban Wildlife. Urban Ecosyst. 2017, 20, 933–943. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of Urbanization on Species Richness: A Review of Plants and Animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Caldwell, A.J.; While, G.M.; Wapstra, E. Plasticity of Thermoregulatory Behaviour in Response to the Thermal Environment by Widespread and Alpine Reptile Species. Anim. Behav. 2017, 132, 217–227. [Google Scholar] [CrossRef]
- Brandt, E.E.; Kelley, J.P.; Elias, D.O. Temperature Alters Multimodal Signaling and Mating Success in an Ectotherm. Behav. Ecol. Sociobiol. 2018, 72, 191. [Google Scholar] [CrossRef]
- Jara, F.G.; Thurman, L.L.; Montiglio, P.-O.; Sih, A.; Garcia, T.S. Warming-Induced Shifts in Amphibian Phenology and Behavior Lead to Altered Predator–Prey Dynamics. Oecologia 2019, 189, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Battles, A.C.; Kolbe, J.J. Miami Heat: Urban Heat Islands Influence the Thermal Suitability of Habitats for Ectotherms. Glob. Chang. Biol. 2019, 25, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Ackley, J.W.; Angilletta, M.J.; DeNardo, D.; Sullivan, B.; Wu, J. Urban Heat Island Mitigation Strategies and Lizard Thermal Ecology: Landscaping Can Quadruple Potential Activity Time in an Arid City. Urban Ecosyst. 2015, 18, 1447–1459. [Google Scholar] [CrossRef]
- French, S.S.; Webb, A.C.; Hudson, S.B.; Virgin, E.E. Town and Country Reptiles: A Review of Reptilian Responses to Urbanization. Integr. Comp. Biol. 2018, 58, 948–966. [Google Scholar] [CrossRef]
- Brum, P.H.R.; Gonçalves, S.R.A.; Strüssmann, C.; Teixido, A.L. A Global Assessment of Research on Urban Ecology of Reptiles: Patterns, Gaps and Future Directions. Anim. Conserv. 2023, 26, 1–13. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Mertig, A.G.; Liu, J. Landowners and Cat Predation across Rural-to-Urban Landscapes. Biol. Conserv. 2003, 115, 191–201. [Google Scholar] [CrossRef]
- Dickman, C.R. House Cats as Predators in the Australian Environment: Impacts and Management. Hum.-Wildl. Confl. 2009, 3, 41–48. [Google Scholar]
- Moseby, K.E.; Peacock, D.E.; Read, J.L. Catastrophic Cat Predation: A Call for Predator Profiling in Wildlife Protection Programs. Biol. Conserv. 2015, 191, 331–340. [Google Scholar] [CrossRef]
- Cordonnier, M.; Perrot, A.; Ferry, N.; Bonnaud, E.; Baudry, E. Pet Cat Personality Linked to Owner-reported Predation Frequency. Ecol. Evol. 2023, 13, e9651. [Google Scholar] [CrossRef]
- Cecchetti, M.; Crowley, S.L.; McDonald, J.; McDonald, R.A. Owner-Ascribed Personality Profiles Distinguish Domestic Cats That Capture and Bring Home Wild Animal Prey. Appl. Anim. Behav. Sci. 2022, 256, 105774. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Gryz, J.; Goszczyński, J. Predation by Domestic Cats in Rural Areas of Central Poland: An Assessment Based on Two Methods. J. Zool. 2012, 288, 260–266. [Google Scholar] [CrossRef]
- Folinsbee, K.E. Evolution of Venom across Extant and Extinct Eulipotyphlans. C. R. Palevol 2013, 12, 531–542. [Google Scholar] [CrossRef]
- Tomasi, T.E. Function of Venom in the Short-Tailed Shrew, Blarina brevicauda. J. Mammal. 1978, 59, 852–854. [Google Scholar] [CrossRef] [PubMed]
- Nader, I.A.; Martin, R.L. The Shrew as Prey of the Domestic Cat. J. Mammal. 1962, 43, 417. [Google Scholar] [CrossRef]
- Tidemann, C.R.; Yorkston, H.D.; Russack, A.J. The Diet of Cats, Felis catus, on Christmas Island, Indian Ocean. Wildl. Res. 1994, 21, 279–285. [Google Scholar] [CrossRef]
- Caro, T.M. Effects of the Mother, Object Play, and Adult Experience on Predation in Cats. Behav. Neural Biol. 1980, 29, 29–51. [Google Scholar] [CrossRef]
- Zwerts, J.A.; Stephenson, P.J.; Maisels, F.; Rowcliffe, M.; Astaras, C.; Jansen, P.A.; van Der Waarde, J.; Sterck, L.E.; Verweij, P.A.; Bruce, T. Methods for Wildlife Monitoring in Tropical Forests: Comparing Human Observations, Camera Traps, and Passive Acoustic Sensors. Conserv. Sci. Pract. 2021, 3, e568. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Predictor | Estimate (SE) | Statistic | p-Value * |
|---|---|---|---|---|
| Soricids R2adj = 0.48 Pearson estimate = 0.66 Deviance explained = 53.6% | Intercept | −2.78 (0.24) | z = −11.52 | 0.0001 |
| Month | Edf = 5.33 | X2 = 492.31 | 0.01 | |
| Rainfall | Edf = 1.05 | X2 = 3.97 | 0.05 | |
| Latitude × Longitude | Edf = 5.37 | X2 = 45.65 | 0.0001 | |
| HFI | Edf = 3.12 | X2 = 4.94 | 0.14 | |
| Age (Female) | Edf = 4.21 | X2 = 172.49 | 0.0001 | |
| Age (Male) | Edf = 3.45 | X2 = 86.96 | 0.0001 | |
| CatID | Edf = 1154.40 | X2 = 8888.47 | 0.001 | |
| Town | Edf = 0.28 | X2 = 2.53 | 0.74 | |
| Year | Edf = 6.67 | X2 = 827.85 | 0.01 | |
| Cricetids R2adj = 0.41 Pearson estimate = 0.76 Deviance explained = 57.1% | Intercept | −2.16 (0.13) | z = −17.19 | 0.0001 |
| Month | Edf = 4.56 | X2 = 571.14 | 0.05 | |
| Rainfall | Edf = 3.12 | X2 = 13.15 | 0.01 | |
| Latitude × Longitude | Edf = 12.41 | X2 = 82.82 | 0.0001 | |
| HFI | Edf = 3.40 | X2 = 163.78 | 0.0001 | |
| Age (Female) | Edf = 7.50 | X2 = 72.13 | 0.0001 | |
| Age (Male) | Edf = 1.00 | X2 = 2.38 | 0.12 | |
| CatID | Edf = 1309.41 | X2 = 16,494.29 | 0.01 | |
| Town | Edf = 0.60 | X2 = 90.78 | 0.85 | |
| Year | Edf = 5.79 | X2 = 544.12 | 0.82 | |
| Murids R2adj = 0.37 Pearson estimate = 1.16 Deviance explained = 47.2% | Intercept | −0.65 (0.05) | z = −14.14 | 0.0001 |
| Month | Edf = 6.63 | X2 = 209.70 | 0.0001 | |
| Rainfall | Edf = 1.83 | X2 = 1.63 | 0.52 | |
| Latitude × Longitude | Edf = 1.30 | X2 = 49.32 | 0.0001 | |
| HFI | Edf = 2.84 | X2 = 39.99 | 0.0001 | |
| Age (Female) | Edf = 3.72 | X2 = 10.44 | 0.1 | |
| Age (Male) | Edf = 4.73 | X2 = 30.14 | 0.0001 | |
| CatID | Edf = 1828 | X2 = 51,789.30 | 0.79 | |
| Town | Edf = 0.00 | X2 = 0.00 | 0.99 | |
| Year | Edf = 4.97 | X2 = 41.94 | 0.05 | |
| Passeriformes R2adj = 0.18 Pearson estimate = 0.88 Deviance explained = 24.7% | Intercept | −1.16 (0.04) | z = −31.89 | 0.0001 |
| Month | Edf = 6.91 | X2 = 251.62 | 0.0001 | |
| Rainfall | Edf = 1.00 | X2 = 1.69 | 0.19 | |
| Latitude × Longitude | Edf = 5.62 | X2 = 11.61 | 0.1 | |
| HFI | Edf = 4.52 | X2 = 18.14 | 0.01 | |
| Age (Female) | Edf = 4.90 | X2 = 46.01 | 0.0001 | |
| Age (Male) | Edf = 2.71 | X2 = 56.38 | 0.0001 | |
| CatID | Edf = 1045.30 | X2 = 6135.86 | 0.96 | |
| Town | Edf = 0.20 | X2 = 0.57 | 0.77 | |
| Year | Edf = 0.80 | X2 = 1.10 | 0.91 | |
| Lacertilians R2adj = 0.46 Pearson estimate = 0.58 Deviance explained = 55.7% | Intercept | −3.06 (0.08) | z = −37.57 | 0.0001 |
| Month | Edf = 7.61 | X2 = 785.08 | 0.0001 | |
| Rainfall | Edf = 2.79 | X2 = 3.59 | 0.31 | |
| Latitude × Longitude | Edf = 1.86 | X2 = 297.10 | 0.0001 | |
| HFI | Edf = 1.00 | X2 = 11.18 | 0.0001 | |
| Age (Female) | Edf = 3.14 | X2 = 29.88 | 0.0001 | |
| Age (Male) | Edf = 1.00 | X2 = 23.71 | 0.0001 | |
| CatID | Edf = 949.90 | X2 = 4953.17 | 0.001 | |
| Town | Edf = 0.00 | X2 = 0.00 | 0.95 | |
| Year | Edf = 3.56 | X2 = 8.90 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castañeda, I.; Forin-Wiart, M.-A.; Pisanu, B.; de Bouillane de Lacoste, N. Spatiotemporal and Individual Patterns of Domestic Cat (Felis catus) Hunting Behaviour in France. Animals 2023, 13, 3507. https://doi.org/10.3390/ani13223507
Castañeda I, Forin-Wiart M-A, Pisanu B, de Bouillane de Lacoste N. Spatiotemporal and Individual Patterns of Domestic Cat (Felis catus) Hunting Behaviour in France. Animals. 2023; 13(22):3507. https://doi.org/10.3390/ani13223507
Chicago/Turabian StyleCastañeda, Irene, Marie-Amélie Forin-Wiart, Benoît Pisanu, and Nathalie de Bouillane de Lacoste. 2023. "Spatiotemporal and Individual Patterns of Domestic Cat (Felis catus) Hunting Behaviour in France" Animals 13, no. 22: 3507. https://doi.org/10.3390/ani13223507
APA StyleCastañeda, I., Forin-Wiart, M.-A., Pisanu, B., & de Bouillane de Lacoste, N. (2023). Spatiotemporal and Individual Patterns of Domestic Cat (Felis catus) Hunting Behaviour in France. Animals, 13(22), 3507. https://doi.org/10.3390/ani13223507