

Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata
 , , ,
, , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Experimental Diets
2.3. Fish and Feeding Trial
2.4. Growth Performance
2.5. Fulton’s Condition Factor and Somatic Indices
2.6. Stress Parameters
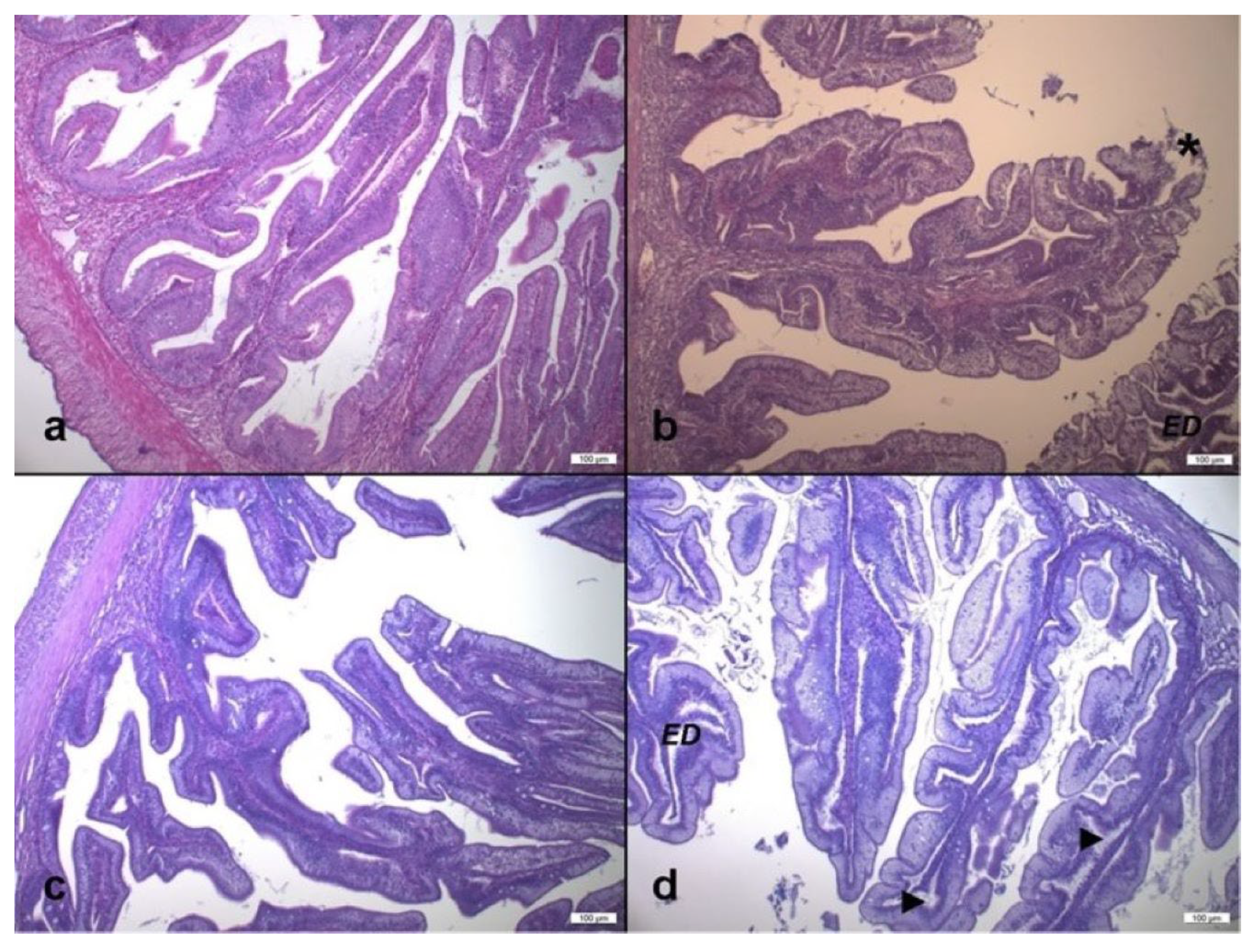
2.7. Gut Histological Analyses
- Epithelium detachment from the lamina propria (ED);
- Microvilli condition (mVC);
- Loss of enterocytes nuclei position (NP);
- Loss of normal supranuclear vacuolation (SNV).
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Somatic Indices and Fulton’s Condition Factor
3.3. Stress Parameters
3.4. Gut Histological Investigations
3.4.1. Gut Morphometric Investigations
3.4.2. Gut Semi-Quantitative Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardinaletti, G.; Randazzo, B.; Messina, M.; Zarantoniello, M.; Giorgini, E.; Zimbelli, A.; Tulli, F. Effects of graded dietary inclusion level of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Capucchio, M.T.; Caldaroni, B.; Magara, G.; Dörr, A.J.M.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; Prearo, M. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. In Feed and Feeding Practices in Aquaculture; Woodhead Publishing: Cambridge, UK, 2015; p. 432. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. Biotechnol. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Bureau, D. Rendered products in fish aquaculture feeds. In Essential Rendering: All About the Animal By-Products Industry; Meeker, D.L., Ed.; National Renderers Association: Alexandria, VA, USA, 2006; p. 302. [Google Scholar]
- Arru, B.; Furesi, R.; Gasco, L.; Madau, F.A.; Pulina, P. The introduction of insect meal into fish diet: The first economic analysis on European sea bass farming. Sustainability 2019, 11, 1697. [Google Scholar] [CrossRef]
- Freccia, A.; Tubin, J.S.B.; Rombenso, A.N.; Emerenciano, M.G.C. Insects in aquaculture nutrition: An emerging eco-friendly approach or commercial reality? In Emerging Technologies, Environment and Research for Sustainable Aquaculture; IntechOpen: London, UK, 2020; pp. 1–14. [Google Scholar]
- Gasco, L.; Acuti, G.; Bani, P.; Dalle Zotte, A.; Danieli, P.P.; De Angelis, A.; Fortina, R.; Marino, R.; Parisi, G.; Piccolo, G. Insect and fish by-products as sustainable alternatives to conventional animal proteins in animal nutrition. Ital. J. Anim. Sci. 2020, 19, 360–372. [Google Scholar] [CrossRef]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquac. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a feed ingredient for fish culture: Status and trends. Aquac. Fish. 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Nairuti, R.N.; Musyoka, S.N.; Yegon, M.J.; Opiyo, M.A. Utilization of Black Soldier Fly (Hermetia illucens Linnaeus) Larvae as a Protein Source for Fish Feed—A Review. Aquac. Stud. 2022, 22, 697. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Antonopoulou, E.; Nikouli, E.; Piccolo, G.; Gasco, L.; Gai, F.; Chatzifotis, S.; Mente, E.; Kormas, K.A. Reshaping gut bacterial communities after dietary Tenebrio molitor larvae meal supplementation in three fish species. Aquaculture 2019, 503, 628–635. [Google Scholar] [CrossRef]
- Bruni, L.; Pastorelli, R.; Viti, C.; Gasco, L.; Parisi, G. Characterisation of the intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens (black soldier fly) partially defatted larva meal as partial dietary protein source. Aquaculture 2018, 487, 56–63. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 2019, 29, 465–486. [Google Scholar] [CrossRef]
- Henry, M.A.; Gasco, L.; Chatzifotis, S.; Piccolo, G. Does dietary insect meal affect the fish immune system? The case of mealworm, Tenebrio molitor on European sea bass, Dicentrarchus labrax. Dev. Comp. Immunol. 2018, 81, 204–209. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed. 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The Potential Role of Insects as Feed: A Multi-Perspective Review. Animals 2019, 9, 119. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EU) 2021/1925 of 5 November 2021 amending certain Annexes to Regulation (EU) No 142/2011 as regards the requirements for placing on the market of certain insect products and the adaptation of a containment method (Text with EEA relevance). C/2021/7824. Off. J. Eur. Union. 2021, 393, 4–8. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=uriserv%3AOJ.L_.2021.393.01.0004.01.ENG&toc=OJ%3AL%3A2021%3A393%3AFULL (accessed on 15 November 2022).
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Secci, G.; Moniello, G.; Gasco, L.; Bovera, F.; Parisi, G. Barbary partridge meat quality as affected by Hermetia illucens and Tenebrio molitor larva meals in feeds. Food Res. Int. 2018, 112, 291–298. [Google Scholar] [CrossRef]
- Allegretti, G.; Talamini, E.; Schmidt, V.; Bogorni, P.C.; Ortega, E. Insect as feed: An emergy assessment of insect meal as a sustainable protein source for the Brazilian poultry industry. J. Clean. Prod. 2018, 171, 403–412. [Google Scholar] [CrossRef]
- Hussein, M.; Pillai, V.V.; Goddard, J.M.; Park, H.G.; Kothapalli, K.S.; Ross, D.A.; Ketterings, Q.M.; Brenna, J.T.; Milstein, M.B.; Marquis, H. Sustainable production of housefly (Musca domestica) larvae as a protein-rich feed ingredient by utilizing cattle manure. PLoS ONE 2017, 12, e0171708. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; De Marco, M.; Martínez, S.; Dabbou, S.; Renna, M.; Madrid, J.; Hernandez, F.; Rotolo, L.; Costa, P.; Gai, F.; et al. Nutritional value of a partially defatted and a highly defatted black soldier fly larvae (Hermetia illucens L.) meal for broiler chickens: Apparent nutrient digestibility, apparent metabolizable energy and apparent ileal amino acid digestibility. J. Anim. Sci. Biotechnol. 2017, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; Dabbou, S.; De Marco, M.; Cullere, M.; Biasato, I.; Biasibetti, E.; Capucchio, M.T.; Bergagna, S.; Dezzutto, D.; Meneguz, M.; et al. Black soldier fly larva fat inclusion in finisher broiler chicken diet as an alternative fat source. Animal 2018, 12, 2032–2039. [Google Scholar] [CrossRef] [PubMed]
- Oteri, M.; Di Rosa, A.R.; Lo Presti, V.; Giarratana, F.; Toscano, G.; Chiofalo, B. Black Soldier Fly Larvae Meal as Alternative to Fish Meal for Aquaculture Feed. Sustainability 2021, 13, 5447. [Google Scholar] [CrossRef]
- Oteri, M.; Chiofalo, B.; Maricchiolo, G.; Toscano, G.; Nalbone, L.; Lo Presti, V.; Di Rosa, A.R. Black Soldier Fly Larvae Meal in the Diet of Gilthead Sea Bream: Effect on Chemical and Microbiological Quality of Filets. Front. Nutr. 2022, 9, 896552. [Google Scholar] [CrossRef]
- Jucker, C.; Lupi, D.; Moore, C.D.; Leonardi, M.G.; Savoldelli, S. Nutrient recapture from insect farm waste: Bioconversion with Hermetia illucens (L.) (Diptera: Stratiomyidae). Sustainability 2020, 12, 362. [Google Scholar] [CrossRef]
- Smetana, S.; Schmitt, E.; Mathys, A. Sustainable use of Hermetia illucens insect biomass for feed and food: Attributional and consequential life cycle assessment. Resour. Conserv. Recycl. 2019, 144, 285–296. [Google Scholar] [CrossRef]
- Danieli, P.P.; Lussiana, C.; Gasco, L.; Amici, A.; Ronchi, B. The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 2019, 9, 178. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.-G.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Cullere, M.; Tasoniero, G.; Giaccone, V.; Miotti-Scapin, R.; Claeys, E.; De Smet, S.; Dalle Zotte, A. Black soldier fly as dietary protein source for broiler quails: Apparent digestibility, excreta microbial load, feed choice, performance, carcass and meat traits. Animal 2016, 10, 1923–1930. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martinez-Sánchez, A.; Pérez Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422, 193–201. [Google Scholar] [CrossRef]
- Oonincx, D.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Pulido, L.; Secci, G.; Maricchiolo, G.; Gasco, L.; Gai, F.; Serra, A.; Conte, G.; Parisi, G. Effect of dietary black soldier fly larvae meal on fatty acid composition of lipids and sn-2 position of triglycerides of marketable size gilthead sea bream fillets. Aquaculture 2022, 546, 737351. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly Prepupae as a Feedstuff for Rainbow Trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetiaillucens)–Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef]
- Esteban, M.A.; Cuesta, A.; Ortuno, J.; Meseguer, J. Immunomodulatory effects of dietary intake of chitin on gilthead seabream (Sparus aurata L.) innate immune system. Fish Shellfish Immunol. 2001, 11, 303–315. [Google Scholar] [CrossRef]
- Lock, E.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Effects of insect diets on the gastrointestinal tract health and growth performance of Siberian sturgeon (Acipenser baerii Brandt, 1869). BMC Vet. Res. 2019, 15, 348. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.; Daskalopoulou, E.; Vogiatzis, I.; Christos, R.; Eleni, M.; Christos, A. Substitution of Fishmeal by Fly Hermetia illucens Prepupae Meal in the Diet of Gilthead Seabream (Sparus aurata). In Proceedings of the HydroMedit, Volos, Greece, 13–15 November 2014. [Google Scholar]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Olivotto, I. Physiological response of rainbow trout (Oncorhynchus mykiss) to graded levels of Hermetia illucens or poultry by-product meals as single or combined substitute ingredients to dietary plant proteins. Aquaculture 2021, 538, 736550. [Google Scholar] [CrossRef]
- FEAP. European Aquaculture Production Report 2014–2019; FEAP: Brussels, Belgium, 2020; pp. 1–47. Available online: https://feap.info/wp-content/uploads/2020/10/20201007_feap-production-report-2020.pdf (accessed on 20 December 2022).
- European Directive. Legislative Decree No. 26 implementing European Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Ital. Leg. 2014, 61, 1–36. [Google Scholar]
- European Directive. European Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. 2010, 276, 33–79. [Google Scholar]
- National Research Council (NCR). Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011; p. 376. [Google Scholar]
- AOAC. Official Methods of Analysis, 20th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2015. [Google Scholar]
- García-Ortega, A.; Kissinger, K.R.; Trushenski, J.T. Evaluation of fish meal and fish oil replacement by soybean protein and algal meal from Schizochytrium limacinum in diets for giant grouper Epinephelus lanceolatus. Aquaculture 2016, 452, 1–8. [Google Scholar] [CrossRef]
- Penn, M.H.; Bendiksen, E.Å.; Campbell, P.; Krogdahl, Å. High level of dietary pea protein concentrate induces enteropathy in Atlantic salmon (Salmo salar L.). Aquaculture 2011, 310, 267–273. [Google Scholar] [CrossRef]
- Urán, P.A.; Gonçalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.J.; Rombout, J.H.W.M. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef]
- Baeza-Ariño, R.; Martínez-Llorens, S.; Nogales-Mérida, S.; Jover-Cerda, M.; Tomás-Vidal, A. Study of liver and gut alterations in sea bream, Sparus aurata L., fed a mixture of vegetable protein concentrates. Aquac. Res. 2016, 47, 460–471. [Google Scholar] [CrossRef]
- Monge-Ortiz, R.; Martínez-Llorens, S.; Márquez, L.; Moyano, F.J.; Jover-Cerdá, M.; Tomás-Vidal, A. Potential use of high levels of vegetal proteins in diets for market-sized gilthead sea bream (Sparus aurata). Arch. Anim. Nutr. 2016, 70, 155–172. [Google Scholar] [CrossRef]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft Inc: New York, NY, USA, 2022. Available online: https://www.xlstat.com/en/news/2022 (accessed on 20 December 2022).
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.-J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Tippayadara, N.; Dawood, M.A.; Krutmuang, P.; Hoseinifar, S.H.; Doan, H.V.; Paolucci, M. Replacement of fish meal by Black soldier fly (Hermetia illucens) larvae meal: Effects on growth, haematology, and skin mucus immunity of Nile Tilapia, Oreochromis niloticus. Animals 2021, 11, 193. [Google Scholar] [CrossRef]
- Muddasir, J.; Imtiaz, A. Length weight relationship and condition factor of snow trout, Schizothorax plagiostomus (Heckel, 1838) from Lidder River, Kashmir. Int. J. Fish. Aquat. 2016, 4, 131–136. [Google Scholar]
- Palmegiano, G.B.; Daprà, F.; Forneris, G.; Gai, F.; Gasco, L.; Guo, K.; Peiretti, P.G.; Sicuro, B.; Zoccarato, I. Rice protein concentrate meal as a potential ingredient in practical diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 258, 357–367. [Google Scholar] [CrossRef]
- Vargas-Abúndez, A.J.; Randazzo, B.; Foddai, M.; Sanchini, L.; Truzzi, C.; Giorgini, E.; Gasco, L.; Olivotto, I. Insect meal based diets for clownfish: Biometric, histological, spectroscopic, biochemical and molecular implications. Aquaculture 2019, 498, 1–11. [Google Scholar] [CrossRef]
- Vargas, A.; Randazzo, B.; Riolo, P.; Truzzi, C.; Gioacchini, G.; Giorgini, E.; Loreto, N.; Ruschioni, S.; Zarantoniello, M.; Antonucci, M.; et al. Rearing zebrafish on black soldier fly (Hermetia illucens): Biometric, histological, spectroscopic, biochemical, and molecular implications. Zebrafish 2018, 15, 404–419. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial dietary inclusion of Hermetia illucens (black soldier fly) full-fat prepupae in zebrafish feed: Biometric, histological, biochemical, and molecular implications. Zebrafish 2018, 15, 519–532. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Truzzi, C.; Giorgini, E.; Marcellucci, C.; Vargas-Abúndez, J.A.; Zimbelli, A.; Annibaldi, A.; Parisi, G.; Tulli, F.; et al. A six-months study on black soldier fly (Hermetia illucens) based diets in zebrafish. Sci. Rep. 2019, 9, 8598. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black soldier fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518, 734659. [Google Scholar] [CrossRef]
- Bruni, L.; Belghit, I.; Lock, E.-J.; Secci, G.; Taiti, C.; Parisi, G. Total replacement of dietary fish meal with black soldier fly (Hermetia illucens) larvae does not impair physical, chemical or volatile composition of farmed Atlantic salmon (Salmo salar L.). J. Sci. Food Agric. 2020, 100, 1038–1047. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Sadler, J.; Wells, R.M.G.; Pankhurst, P.M.; Pankhurst, N.W. Blood oxygen transport, rheology and hematological responses to confinement stress in diploid and triploid Atlantic salmon, Salmo salar. Aquaculture 2000, 184, 349–361. [Google Scholar] [CrossRef]
- Campbell, T.W. Blood biochemistry in lower vertebrates. In Proceedings of the 55th Annual Meeting of the American College of Veterinary Pathologists (ACVP) & 39th Annual Meeting of the American Society of Clinical Pathology (ASVCP), ACVP and ASVCP, Orlando, FL, USA, 13–17 November 2004. [Google Scholar]
- Wagner, T.; Congleton, J.L. Blood chemistry correlates of nutritional condition, tissue damage, and stress in migrating juvenile chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. 2004, 61, 1066–1074. [Google Scholar] [CrossRef]
- Pottinger, T.G.; Mosuwe, E. The corticosteroidogenic response of brown and rainbow trout alevins and fry to environmental stress during a critical period. Gen. Comp. Endocrinol. 1994, 95, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar-Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Pottinger, T.G.; Rand-Weaver, M.; Sumpter, J.P. Overwinter fasting and re-feeding in rainbow trout: Plasma growth hormone and cortisol levels in relation to energy mobilization. Comp. Biochem. Physiol. B 2003, 136, 403–417. [Google Scholar] [CrossRef]
- Haukenes, A.H.; Barton, B.A.; Bolligs, H. Cortisol responses of pallid sturgeon and yellow perch folowing challenge with lipopolysaccharide. J. Fish Biol. 2008, 72, 780–784. [Google Scholar] [CrossRef]
- Silbergeld, E.K. Blood glucose: A sensitive indicator of environmental stress in fish. Bull. Environ. Contam. Toxicol. 1974, 11, 20–25. [Google Scholar] [CrossRef]
- Wedemeyer, G.A.; Yasutake, W.T. Clinical methods for the assessment of the effects of environmental stress on fish health. Technical Paper no 89. In United States Geological Survey; U.S. Fish and Wildlife Service: Washington, DC, USA, 1977. [Google Scholar]
- David, M.; Shivakumar, R.; Mushigeri, S.B.; Kuri, R.C. Blood glucose and glycogen levels as indicators of stress in the freshwater fish, Labeo rohita under fenvalerate intoxication. J. Ecotoxicol. Environ. Monit. 2005, 15, 1–5. [Google Scholar]
- Martinez-Porchas, M.; Martinez-Córdova, L.R.; Ramos-Enriquez, R. Cortisol and Glucose: Reliable indicators of fish stress? Pan-Am. J. Aquat. Sci. 2009, 4, 158–178. [Google Scholar]
- Roche, H.; Bogé, G. Fish blood parameters as a potential tool for identification of stress caused by environmental factors and chemical intoxication. Mar. Environ. Res. 1996, 41, 27–43. [Google Scholar] [CrossRef]
- Soivio, A.; Oikari, A. Haematological effects of stress on a teleost, Esox lucius L. J. Fish Biol. 1976, 8, 397–411. [Google Scholar] [CrossRef]
- Zhou, J.S.; Liu, S.S.; Ji, H.; Yu, H.B. Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2018, 24, 424–433. [Google Scholar] [CrossRef]
- Yildirim-Aksoy, M.; Eljack, R.; Schrimsher, C.; Beck, B.H. Use of dietary frass from black soldier fly larvae, Hermetia illucens, in hybrid tilapia (Nile x Mozambique, Oreocromis niloticus x O. mozambique) diets improves growth and resistance to bacterial diseases. Aquac. Rep. 2020, 17, 100373. [Google Scholar] [CrossRef]
- Raskovic, B.; Stankovic, M.; Markovic, Z.; Poleksic, V. Histological methods in the assessment of different feed effects on liver and intestine of fish. J. Agric. Sci. 2011, 56, 87–100. [Google Scholar] [CrossRef]
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal alternative protein sources for aquaculture feeds. In Feeds for the Aquaculture Sector; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–28. [Google Scholar]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [Google Scholar] [CrossRef]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Belghit, I.; Lock, E.-J.; Krogdahl, Å. Total replacement of fish meal with black soldier fly (Hermetia illucens) larvae meal does not compromise the gut health of Atlantic salmon (Salmo salar). Aquaculture 2020, 520, 734967. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Secci, G.; Notarstefano, V.; Giorgini, E.; Lock, E.J.; Parisi, G.; Olivotto, I. Application of laboratory methods for understanding fish responses to Black Soldier Fly (Hermetia illucens) based diets. J. Insects Food Feed 2021, 8, 1173–1195. [Google Scholar] [CrossRef]
- Santigosa, E.; Sánchez, J.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J.; Gallardo, M.A. Modifications of digestive enzymes in trout (Oncorhynchus mykiss) and sea bream (Sparus aurata) in response to dietary fish meal replacement by plant protein sources. Aquaculture 2008, 282, 68–74. [Google Scholar] [CrossRef]
- Wagner, C.E.; McIntyre, P.B.; Buels, K.S.; Gilbert, D.M.; Michel, E. Diet predicts intestine length in Lake Tanganyika’s cichlid fishes. Funct. Ecol. 2009, 23, 1122–1131. [Google Scholar] [CrossRef]
- Melenchón, F.; de Mercado, E.; Pula, H.J.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Tomás-Almenar, C. Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals 2022, 12, 179. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Lu, R.; Chen, Y.; Yu, W.; Lin, M.; Yang, G.; Qin, C.; Meng, X.; Zhang, Y.; Ji, H.; Nie, G. Defatted black soldier fly (Hermetia illucens) larvae meal can replace soybean meal in juvenile grass carp (Ctenopharyngodon idellus) diets. Aquac. Rep. 2020, 18, 100520. [Google Scholar] [CrossRef]
- Hender, A.; Siddik, M.A.; Howieson, J.; Fotedar, R. Black Soldier Fly, Hermetia illucens as an Alternative to Fishmeal Protein and Fish Oil: Impact on Growth, Immune Response, Mucosal Barrier Status, and Flesh Quality of Juvenile Barramundi, Lates calcarifer (Bloch, 1790). Biology 2021, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Fronte, B.; Licitra, R.; Bibbiani, C.; Casini, L.; De Zoysa, M.; Miragliotta, V.; Sagona, S.; Coppola, F.; Brogi, L.; Abramo, F. Fishmeal replacement with hermetia illucens meal in aquafeeds: Effects on zebrafish growth performances, intestinal morphometry, and enzymology. Fishes 2021, 6, 28. [Google Scholar] [CrossRef]
- Machado, M.R.F.; Souza, H.O.; de Souza, V.L.; de Azevedo, A.; Goitein, R.; Nobre, A.D. Morphological and anatomical characteriza- tion of the digestive tract of Centropomus parallelus and C. un- decimalis. Acta Sci. Biol. Sci. 2013, 35, 467–474. [Google Scholar] [CrossRef]
- Rossi, W., Jr.; Allen, K.M.; Habte-Tsion, H.M.; Meesala, K.M. Supplementation of glycine, prebiotic, and nucleotides in soybean meal-based diets for largemouth bass (Micropterus salmoides): Effects on production performance, whole-body nutrient composition and retention, and intestinal histopathology. Aquaculture 2021, 532, 736031. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Tibaldi, E.; Cardinaletti, G.; Giorgini, E.; Lunelli, F.; Olivotto, I. A multidisciplinary approach to investigate biological effects on intestine phisiology and appetite stimulus in rainbow trout Onchorhincus mykiss fed diets with graded levels of insect meal and poultry by-product meal. In Aquaculture America 2020; World Aquaculture Society: Honolulu, HI, USA, 2020; Available online: https://openpub.fmach.it/bitstream/10449/67298/1/2020%20AA%20552%20Lunelli.pdf (accessed on 20 December 2022).
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) physiological and behavioural responses to insect-based diets: A multidisciplinary approach. Sci. Rep. 2020, 10, 10648. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Yu, H.; Zhou, J. Influence of dietary black soldier fly (Hermetia illucens Linnaeus) pulp on growth performance, antioxidant capacity and intestinal health of juvenile mirror carp (Cyprinus carpio var. specularis). Aquac. Nutr. 2020, 26, 432–443. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]


{kind=link}
| HIM0 | HIM25 | HIM35 | HIM50 | |
|---|---|---|---|---|
| Ingredients, % as fed | ||||
| Fish meal | 25.00 | 18.75 | 16.25 | 12.50 |
| Hermetia illucens meal | 0 | 7.90 | 11.00 | 15.70 |
| Soy protein concentrate | 5.00 | 5.00 | 5.00 | 5.00 |
| Wheat gluten | 5.00 | 5.00 | 5.00 | 5.00 |
| Corn gluten | 5.00 | 5.00 | 5.00 | 5.00 |
| Soybean meal 48 | 15.00 | 15.00 | 15.00 | 15.00 |
| Rapeseed meal | 5.00 | 5.00 | 5.00 | 5.00 |
| Wheat meal | 17.45 | 15.17 | 14.21 | 12.88 |
| Whole peas | 4.00 | 4.00 | 4.00 | 4.00 |
| Fish oil | 5.00 | 5.00 | 5.00 | 5.00 |
| Rapeseed oil | 10.00 | 9.80 | 9.80 | 9.80 |
| Vitamin and mineral premix | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin C35 | 0.03 | 0.03 | 0.03 | 0.03 |
| Vitamin E50 | 0.02 | 0.02 | 0.02 | 0.02 |
| Antioxidant | 0.30 | 0.30 | 0.30 | 0.30 |
| Sodium propionate | 0.10 | 0.10 | 0.10 | 0.10 |
| MCP, monocalcium phosphate | 1.50 | 2.20 | 2.50 | 2.80 |
| L-Lysine | 0.30 | 0.35 | 0.37 | 0.40 |
| L-Tryptophan | - | 0.03 | 0.04 | 0.05 |
| DL-Methionine | 0.10 | 0.15 | 0.18 | 0.22 |
| L-Taurine | 0.20 | 0.20 | 0.20 | 0.20 |
| Chemical composition, % as fed | ||||
| Dry matter | 92.33 | 92.78 | 92.90 | 92.64 |
| Crude protein | 42.7 | 42.7 | 42.7 | 42.7 |
| Ether extract | 18.6 | 18.6 | 18.6 | 18.7 |
| Crude fiber | 2.3 | 2.2 | 2.2 | 2.1 |
| Ash | 9.3 | 9.3 | 9.4 | 9.3 |
| NFE * | 19.43 | 19.98 | 20.00 | 19.84 |
| Gross Energy, MJ/kg feed # | 22.0 | 21.9 | 21.9 | 21.9 |
| GROUP | p-Value | SEM | |||||
|---|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM35 | HIM50 | D | IBW | ||
| Number of fish | 78 | 78 | 78 | 78 | |||
| IBW, g | 144 | 144 | 144 | 144 | 2.883 | ||
| FBW, g | 387 | 386 | 396 | 394 | 0.473 | 0.762 | 5.516 |
| FCR | 1.42 | 1.43 | 1.41 | 1.42 | 0.979 | 0.866 | 0.030 |
| SGR, %/d | 0.76 | 0.76 | 0.76 | 0.74 | 0.774 | 0.758 | 0.014 |
| PER | 1.82 | 1.80 | 1.77 | 1.76 | 0.746 | 0.829 | 0.041 |
| DIR, % | 18.30 | 18.43 | 17.77 | 17.56 | 0.137 | 0.261 | 0.269 |
| K | 1.908 a | 1.867 ab | 1.828 b | 1.852 ab | 0.008 | <0.0001 | 0.016 |
| GROUP | p-Value | SEM | |||||
|---|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM35 | HIM50 | D | FBW | ||
| Number of fish | 18 | 18 | 18 | 18 | |||
| Body weight, g | 394 | 407 | 406 | 410 | 0.114 | 5.163 | |
| VSI, % | 0.251 a | 0.212 c | 0.234 b | 0.239 ab | <0.0001 | 0.544 | 0.004 |
| HSI, % | 0.065 ab | 0.104 a | 0.062 b | 0.090 ab | 0.018 | <0.0001 | 0.011 |
| GROUP | p-Value | SEM | |||||
|---|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM35 | HIM50 | D | FBW | ||
| Number of fish | 18 | 18 | 18 | 18 | |||
| Cortisol, ng/mL | 21.693 | 19.972 | 19.166 | 18.492 | 0.541 | 0.261 | 1.623 |
| Glucose, mg/DL | 63.371 | 66.715 | 63.277 | 65.317 | 0.799 | 0.754 | 2.829 |
| Lactate, mg/DL | 19.010 | 20.555 | 19.082 | 18.782 | 0.867 | 0.323 | 1.631 |
| Total Proteins, mg/mL | 33.495 | 33.197 | 30.134 | 31.118 | 0.448 | 0.936 | 1.693 |
| ACTH, pg/mL | 141.272 | 145.767 | 147.839 | 146.571 | 0.991 | 0.997 | 14.942 |
| GROUP | p-Value | SEM | |||||
|---|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM35 | HIM50 | D | FBW | ||
| Number of fish | 18 | 18 | 18 | 18 | |||
| Vl Anterior gut, μm | 1593.798 ab | 1430.240 b | 1638.236 a | 1487.868 ab | 0.024 | 0.110 | 53.511 |
| Vl Posterior gut, μm | 1106.586 | 1107.966 | 1086.043 | 1174.447 | 0.613 | 0.022 | 49.475 |
| Vw Anterior gut, μm | 306.016 ab | 287.608 b | 341.124 a | 352.785 a | 0.002 | 0.939 | 13.237 |
| Vw Posterior gut, μm | 153.452 b | 159.187 ab | 165.904 ab | 177.309 a | 0.033 | 0.513 | 5.945 |
| Gc Anterior gut, nr. | 139.722 b | 173.763 a | 171.066 a | 134.569 b | <0.0001 | 0.321 | 6.906 |
| Gc Posterior gut, nr. | 128.119 | 135.355 | 124.173 | 119.603 | 0.554 | 0.053 | 7.98 |
| GROUP | p-Value | SEM | |||||
|---|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM35 | HIM50 | D | FBW | ||
| Number of fish | 18 | 18 | 18 | 18 | |||
| ED Anterior gut | 3.495 | 3.041 | 3.216 | 3.582 | 0.459 | 0.059 | 0.267 |
| ED Posterior gut | 2.323 | 3.098 | 3.050 | 3.363 | 0.144 | 0.164 | 0.325 |
| mVC Anterior gut | 1.737 | 2.051 | 1.665 | 2.213 | 0.444 | 0.571 | 0.272 |
| mVC Posterior gut | 1.521 | 2.510 | 2.060 | 2.020 | 0.122 | 0.219 | 0.285 |
| NP Anterior gut | 2.963 b | 3.772 ab | 3.220 b | 4.433 a | 0.002 | 0.491 | 0.278 |
| NP Posterior gut | 3.086 | 3.047 | 2.996 | 2.927 | 0.987 | 0.334 | 0.323 |
| SNV Anterior gut | 2.670 b | 3.260 ab | 2.659 b | 3.966 a | 0.004 | 0.036 | 0.282 |
| SNV Posterior gut | 2.408 | 3.050 | 2.886 | 3.433 | 0.199 | 0.557 | 0.335 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Rosa, A.R.; Caccamo, L.; Pansera, L.; Oteri, M.; Chiofalo, B.; Maricchiolo, G. Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata. Animals 2023, 13, 339. https://doi.org/10.3390/ani13030339
Di Rosa AR, Caccamo L, Pansera L, Oteri M, Chiofalo B, Maricchiolo G. Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata. Animals. 2023; 13(3):339. https://doi.org/10.3390/ani13030339
Chicago/Turabian StyleDi Rosa, Ambra Rita, Letteria Caccamo, Lidia Pansera, Marianna Oteri, Biagina Chiofalo, and Giulia Maricchiolo. 2023. "Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata" Animals 13, no. 3: 339. https://doi.org/10.3390/ani13030339
APA StyleDi Rosa, A. R., Caccamo, L., Pansera, L., Oteri, M., Chiofalo, B., & Maricchiolo, G. (2023). Influence of Hermetia illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata. Animals, 13(3), 339. https://doi.org/10.3390/ani13030339