
Lactobacillus salivarius and Berberine Alleviated Yak Calves’ Diarrhea via Accommodating Oxidation Resistance, Inflammatory Factors, and Intestinal Microbiota
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Detection of Oxidation Resistance and Inflammatory Factors in Yaks
2.3. Intestinal Microbiota Sequencing of Yak Calves
2.4. Sequencing Data Analyzing of Yak Calves
2.5. Statistical Analysis
3. Results
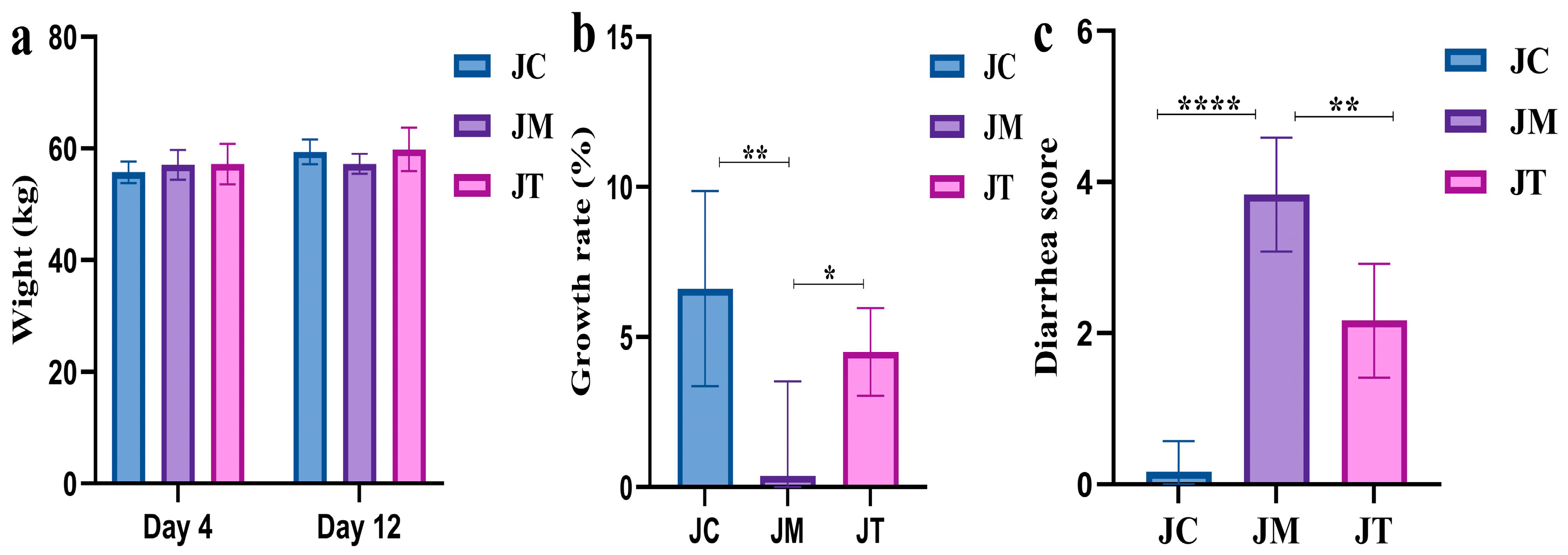
3.1. Lactobacillus salivarius and Berberine Reduced Diarrhea in Yak Calves
3.2. Lactobacillus salivarius and Berberine Increased Oxidation Resistance and Dropped Inflammation Levels in Ruminants
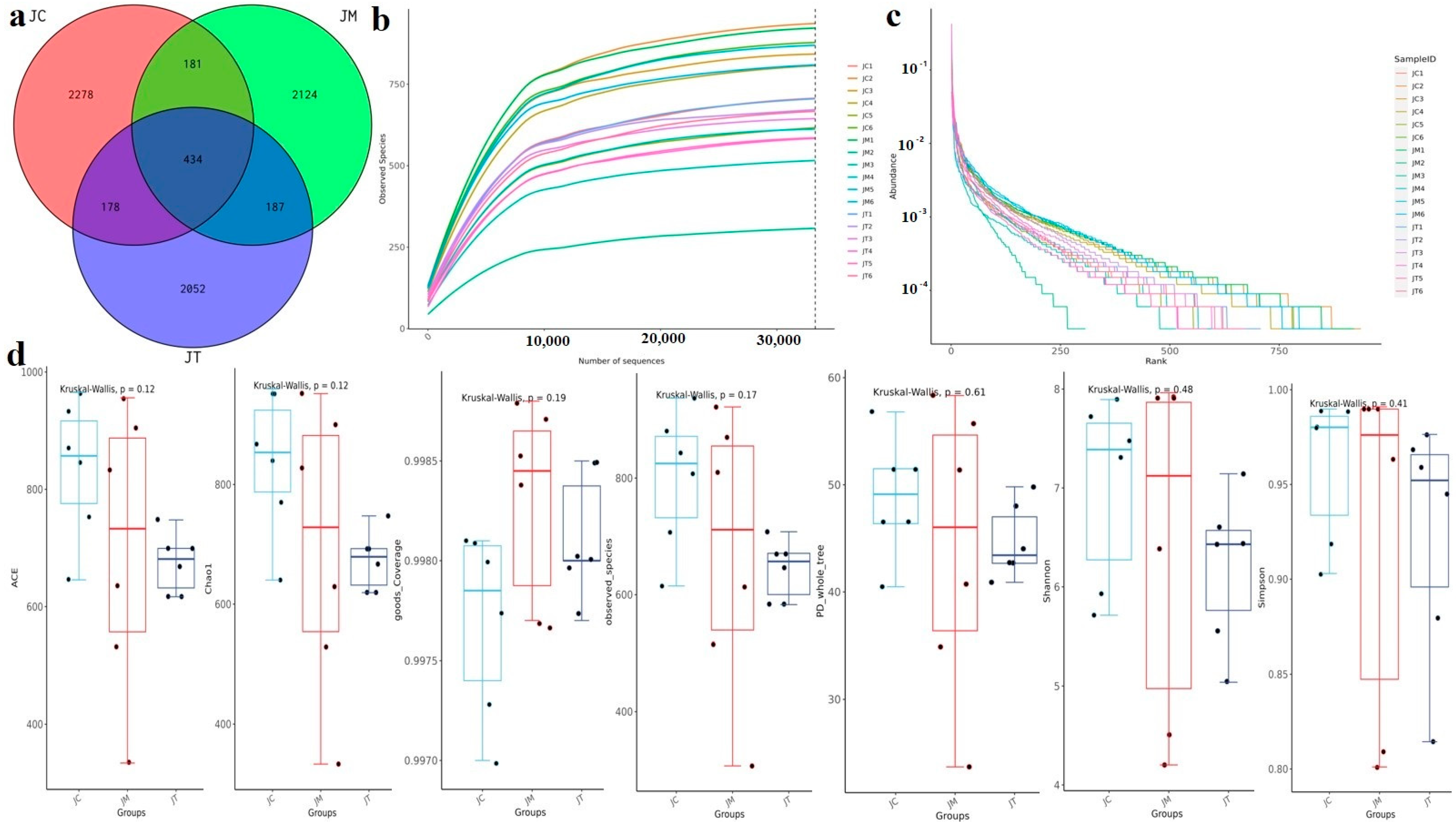
3.3. Lactobacillus salivarius and Berberine Mediated the Microbial Community in Yaks in Different Taxa
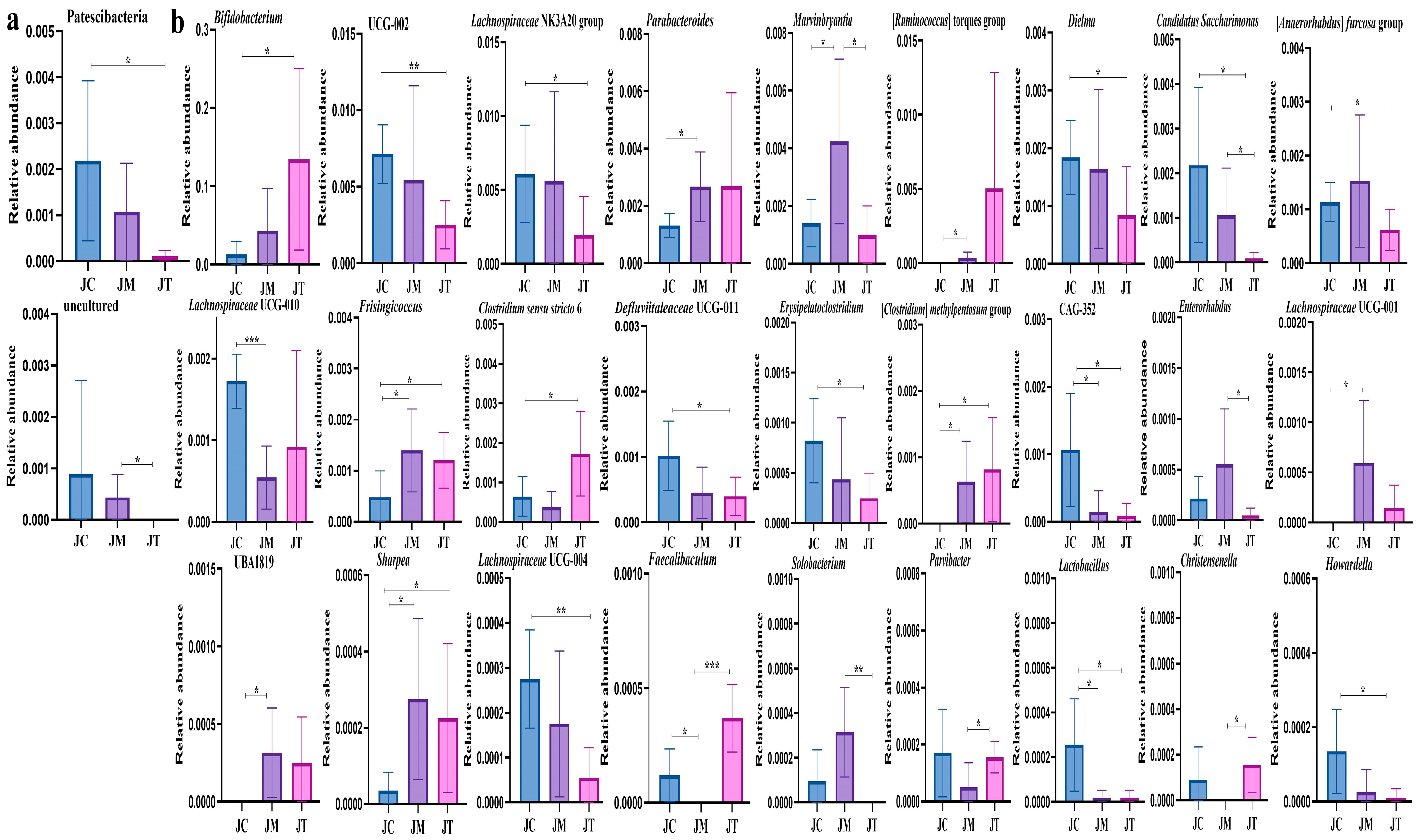
3.4. Distinguished Species among Yak Calves in Different Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, J.; Xin, Z.; Huang, Y.; Yu, J. Climate suitability assessment on the Qinghai-Tibet Plateau. Sci. Total Environ. 2022, 816, 151653. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Wang, H.; Degen, A.A.; Zhou, J.; Guo, N.; Mudassar, S.; Long, R. Rumen parameters of yaks (Bos grunniens) and indigenous cattle (Bos taurus) grazing on the Qinghai-Tibetan Plateau. J. Anim. Physiol. Anim. Nutr. 2019, 103, 969–976. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, H.; Zhang, R.; Chen, X.; Yu, H.; Zu, Y.; Tan, S.; Wang, X.; Wang, Q.; Xu, W. Comparative analysis of nasal microbial community between Tibetan sheep with different ages. Pak. Vet. J. 2023, 43, 449–457. [Google Scholar]
- Li, K.; Mehmood, K.; Zhang, H.; Jiang, X.; Shahzad, M.; Dong, X.; Li, J. Characterization of fungus microbial diversity in healthy and diarrheal yaks in Gannan region of Tibet Autonomous Prefecture. Acta Trop. 2018, 182, 14–26. [Google Scholar] [CrossRef]
- Fan, Q.; Wanapat, M.; Hou, F. Mineral Nutritional Status of Yaks (Bos Grunniens) Grazing on the Qinghai-Tibetan Plateau. Animals 2019, 9, 468. [Google Scholar] [CrossRef]
- García-Valle, J.; Ruiz-Ramírez, J.; Macedo-Barragán, R.; Lizama-Munguía, T.; Gutiérrez-Chávez, A.; García-Valle, M.; García-Márquez, L. Seroprevalence and risk factors associated with sheep neosporosis in Colima, Mexico. Pak. Vet. J. 2023, 44, 195–199. [Google Scholar]
- Shehata, A.A.; El-Emam, A.; Mohamed, M.; Gouda, H.; El-Said, B.M.; Salman, M.B.; Abd-Elfatah, E.B. Molecular characterization of cryptosporidium infection and analysis of hematological and biochemical changes in diarrheic pre-weaned calves in Egypt. Pak. Vet. J. 2024, 44, 135–140. [Google Scholar]
- Schinwald, M.; Creutzinger, K.; Keunen, A.; Winder, C.B.; Haley, D.; Renaud, D.L. Predictors of diarrhea, mortality, and weight gain in male dairy calves. J. Dairy Sci. 2022, 105, 5296–5309. [Google Scholar] [CrossRef]
- Tras, B.; Ok, M.; Parlak, T.M.; Ider, M.; Yildiz, R.; Eser Faki, H.; Ozdemir Kutahya, Z.; Uney, K. Can diarrhea affect the pharmacokinetics of racecadotril in neonatal calves? J. Vet. Pharmacol. Ther. 2022, 45, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Sulehria, M.U.; Ahmad, S.S.; Ijaz, M.; Mushtaq, M.H.; Ahmad, S.F. Molecular detection and hematological changes associated with the canine diarrheic viruses in Pakistan. Pak. Vet. J. 2023, 43, 732–738. [Google Scholar]
- Wang, D.; Du, Y.; Wang, S.; You, Z.; Liu, Y. Effects of sodium humate and glutamine combined supplementation on growth performance, diarrhea incidence, blood parameters, and intestinal microflora of weaned calves. Anim. Sci. J. 2021, 92, e13584. [Google Scholar] [CrossRef]
- Algammal, A.M.; Hetta, H.F.; Batiha, G.E.; Hozzein, W.N.; El, K.W.; Hashem, H.R.; Tawfik, A.M.; El-Tarabili, R.M. Virulence-determinants and antibiotic-resistance genes of MDR-E. coli isolated from secondary infections following FMD outbreak in cattle. Sci. Rep. 2020, 10, 19779. [Google Scholar] [CrossRef] [PubMed]
- Soon, J.M.; Chadd, S.A.; Baines, R.N. Escherichia coli O157:H7 in beef cattle: On farm contamination and pre-slaughter control methods. Anim. Health Res. Rev. 2011, 12, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, T.; Nimmo, G.R. Antibiotic resistance is an emerging threat to public health: An urgent call to action at the Antimicrobial Resistance Summit 2011. Med. J. Aust. 2011, 194, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Tao, M.; Liu, J.; Zhang, X.; Hu, J.; Ma, B.; Chen, P. In vitro evaluation of antibiotic-Matrine combinations against clinically relevant bacteria. Pak. Vet. J. 2024, 44, 162–168. [Google Scholar]
- Astorga, F.; Navarrete-Talloni, M.J.; Miró, M.P.; Bravo, V.; Toro, M.; Blondel, C.J.; Hervé-Claude, L.P. Antimicrobial resistance in E. coli isolated from dairy calves and bedding material. Heliyon 2019, 5, e02773. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, Y.; Cerino, B.; Moreno, M.; Yañez, E.; Heredia, N.; Dávila-Aviña, J.; Quezada, T.; Calle, A.; García, S. Diarrheagenic Escherichia coli with Multidrug Resistance in Cattle from Mexico. J. Food Prot. 2024, 87, 100257. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Mehmood, A.; Nawaz, M.; Rabbani, M.; Mushtaq, M.H. Probiotic effect of Limosilactobacillus fermentum on growth performance and competitive exclusion of Salmonella gallinarum in poultry. Pak. Vet. J. 2023, 43, 659–664. [Google Scholar]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Zhuge, A.; Li, B.; Yuan, Y.; Lv, L.; Li, Y.; Wu, J.; Yang, L.; Bian, X.; Wang, K.; Wang, Q.; et al. Lactobacillus salivarius LI01 encapsulated in alginate-pectin microgels ameliorates D-galactosamine-induced acute liver injury in rats. Appl. Microbiol. Biotechnol. 2020, 104, 7437–7455. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, Y.; Luo, A.; Heng, X.; Liu, J.; Wang, H.; Chu, W. Lactobacillus salivarius CPU-01 Ameliorates Temozolomide-Induced Intestinal Mucositis by Modulating Gut Microbiota, Maintaining Intestinal Barrier, and Blocking Pro-inflammatory Cytokines. Probiotics Antimicrob. Proteins 2023, 15, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; He, Z.; Wang, T.; Wang, X.; Wang, T.; Long, M. Lactobacillus salivarius WZ1 Inhibits the Inflammatory Injury of Mouse Jejunum Caused by Enterotoxigenic Escherichia coli K88 by Regulating the TLR4/NF-κB/MyD88 Inflammatory Pathway and Gut Microbiota. Microorganisms 2023, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, X.; Yang, R.; Chen, F.; Liao, Y.; Zhu, Z.; Wu, Z.; Sun, X.; Wang, L. Effects of Berberine on the Gastrointestinal Microbiota. Front. Cell. Infect. Microbiol. 2021, 10, 588517. [Google Scholar] [CrossRef]
- Song, D.; Hao, J.; Fan, D. Biological properties and clinical applications of berberine. Front. Med. 2020, 14, 564–582. [Google Scholar] [CrossRef]
- Kumar, A.; Ekavali, C.K.; Mukherjee, M.; Pottabathini, R.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef]
- Li, L.; Chang, L.; Zhang, X.; Ning, Z.; Mayne, J.; Ye, Y.; Stintzi, A.; Liu, J.; Figeys, D. Berberine and its structural analogs have differing effects on functional profiles of individual gut microbiomes. Gut Microbes 2020, 11, 1348–1361. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Khadka, D.B.; Cho, W.J. Pharmacological effects of berberine and its derivatives: A patent update. Expert. Opin. Ther. Patents 2016, 26, 229–243. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Xia, Y.; Wang, Y.; Han, D.; Liu, Y.; Li, J.; Fu, J.; Wang, L.; Gan, Z.; Liu, B.; et al. Gut microbiota bridges dietary nutrients and host immunity. Sci. China Life Sci. 2023, 66, 2466–2514. [Google Scholar] [CrossRef]
- Yu, S.; Sun, Y.; Shao, X.; Zhou, Y.; Yu, Y.; Kuai, X.; Zhou, C. Leaky Gut in IBD: Intestinal Barrier-Gut Microbiota Interaction. J. Microbiol. Biotechnol. 2022, 32, 825–834. [Google Scholar] [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Feng, S.; Li, Y.; Zhang, C.; Chao, G.; Zhang, S. Gut microbiota and intestinal immunity—A crosstalk in irritable bowel syndrome. Immunology 2024, 172, 1–20. [Google Scholar] [CrossRef]
- Li, K.; Zeng, Z.; Liu, J.; Pei, L.; Wang, Y.; Li, A.; Kulyar, M.F.; Shahzad, M.; Mehmood, K.; Li, J.; et al. Effects of Short-Chain Fatty Acid Modulation on Potentially Diarrhea-Causing Pathogens in Yaks Through Metagenomic Sequencing. Front. Cell. Infect. Microbiol. 2022, 12, 805481. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, M.; Aldridge, B.; Lowe, J. Dysbiosis of the fecal microbiota in feedlot cattle with hemorrhagic diarrhea. Microb. Pathog. 2018, 115, 123–130. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Zhang, W.; Kulyar, M.F.; Ullah, K.; Han, Z.; Qin, J.; Bi, C.; Wang, Y.; Li, K. Comparative analysis of gut microbiota in healthy and diarrheic yaks. Microb. Cell. Fact. 2022, 21, 111. [Google Scholar] [CrossRef]
- Li, W.; Zhang, L.; Xu, Q.; Yang, W.; Zhao, J.; Ren, Y.; Yu, Z.; Ma, L. Taxifolin Alleviates DSS-Induced Ulcerative Colitis by Acting on Gut Microbiome to Produce Butyric Acid. Nutrients 2022, 14, 1069. [Google Scholar] [CrossRef]
- Chen, X.; Kong, Q.; Zhao, X.; Zhao, C.; Hao, P.; Irshad, I.; Lei, H.; Kulyar, M.F.; Bhutta, Z.A.; Ashfaq, H.; et al. Sodium acetate/sodium butyrate alleviates lipopolysaccharide-induced diarrhea in mice via regulating the gut microbiota, inflammatory cytokines, antioxidant levels, and NLRP3/Caspase-1 signaling. Front. Microbiol. 2022, 13, 1036042. [Google Scholar] [CrossRef]
- Parker, A.; Romano, S.; Ansorge, R.; Aboelnour, A.; Gall, G.L.; Savva, G.M.; Pontifex, M.G.; Telatin, A.; Baker, D.; Jones, E.; et al. Fecal microbiota transfer between young and aged mice reverses hallmarks of the aging gut, eye, and brain. Microbiome 2022, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Abellan-Schneyder, I.; Matchado, M.S.; Reitmeier, S.; Sommer, A.; Sewald, Z.; Baumbach, J.; List, M.; Neuhaus, K. Primer, Pipelines, Parameters: Issues in 16S rRNA Gene Sequencing. mSphere 2021, 6, 01202–01220. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Dubois, B.; Debode, F.; Hautier, L.; Hulin, J.; Martin, G.S.; Delvaux, A.; Janssen, E.; Mingeot, D. A detailed workflow to develop QIIME2-formatted reference databases for taxonomic analysis of DNA metabarcoding data. BMC Genom. Data 2022, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Xu, R.; Chen, X.; Yang, C.; Jiang, F.; Shen, Y.; Li, Q.; Fang, F.; Li, Y.; Shen, X. Characterization of gut microbiota in adults with coronary atherosclerosis. PeerJ 2023, 11, e15245. [Google Scholar] [CrossRef]
- Li, B.; Du, P.; Du, Y.; Zhao, D.; Cai, Y.; Yang, Q.; Guo, Z. Luteolin alleviates inflammation and modulates gut microbiota in ulcerative colitis rats. Life Sci. 2021, 269, 119008. [Google Scholar] [CrossRef]
- Li, L.; Wang, F.; Liu, Y.; Gu, F. Intestinal microbiota dysbiosis in children with recurrent respiratory tract infections. Microb. Pathog. 2019, 136, 103709. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; He, C.; Li, Y.; Yu, Z.; Chen, Y.; Wang, Y.; Ni, D. Changes of fungal community and non-volatile metabolites during pile-fermentation of dark green tea. Food Res. Int. 2021, 147, 110472. [Google Scholar] [CrossRef]
- Kim, B.; Kang, N.; Kim, S.; Kim, D.H.; Kim, H.; Kwon, O.J.; Huh, H.J.; Lee, N.Y.; Jhun, B.W. The lung microbiota in nontuberculous mycobacterial pulmonary disease. PLoS ONE 2023, 18, e0285143. [Google Scholar] [CrossRef]
- Mahmoudiandehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider Paisley, A.; Moseley, M.A.; Thompson, J.W.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease—An emerging role for gut microbiome. Alzheimer’s Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, Q.; Wei, Y.; Wang, H.; Ga, Y.; Zhang, Y.; Hao, Z. Qingjie decoction attenuated E. coli-induced diarrhea by regulating energy metabolism and alleviating inflammation based on network analysis and metabolomics. J. Ethnopharmacol. 2024, 318, 116806. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, R.; Burdukiewicz, M.; Schierack, P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. coli. Front. Cell. Infect. Microbiol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.; Barros, B.; Castro-Tardón, D.; Martin, C.; Santos, F.; Knöbl, T.; Santarosa, B.; Padilha, L.; Hurley, D. The role of anti-E. coli antibody from maternal colostrum on the colonization of newborn dairy calves gut with Escherichia coli and the development of clinical diarrhea. Anim.—Open Space 2023, 2, 100037. [Google Scholar] [CrossRef]
- Vanjari, H.; Mohan, M.; Saudagar, P.; Guthale, G.; Dalvi, M. Protective effect of SKB_Gutbiotic against castor oil and E. coli induced diarrhea in laboratory animals. Microb. Pathog. 2020, 143, 104078. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, D.F.; Mahendra, M.Y.N.; Kamaludeen, J.; Pertiwi, H. Lycopene in Feed as Antioxidant and Immuno-Modulator Improves Broiler Chicken’s Performance under Heat-Stress Conditions. Vet. Med. Int. 2023, 2023, 5418081. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Li, H.; Wan, S.; Tang, S. Bacillus pumilus TS1 alleviates Salmonella Enteritidis-induced intestinal injury in broilers. BMC Vet. Res. 2023, 19, 41. [Google Scholar] [CrossRef]
- Kwon, M.J.; Kim, J.H.; Kim, K.J.; Ko, E.J.; Lee, J.Y.; Ryu, C.S.; Ha, Y.H.; Kim, Y.R.; Kim, N.K. Genetic Association between Inflammatory-Related Polymorphism in STAT3, IL-1beta, IL-6, TNF-alpha and Idiopathic Recurrent Implantation Failure. Genes 2023, 14, 1588. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and Functions of the IL-10 Family of Cytokines in Inflammation and Disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Arbaga, A.; Hassan, H.; Anis, A.; Othman, N.; Kamr, A. Clinicopathological and electrophoretic pattern of serum protein alterations in acute pneumonic sheep. Pak. Vet. J. 2023, 43, 303–308. [Google Scholar]
- Bhatia, R.; Sharma, S.; Bhadada, S.K.; Bishnoi, M.; Kondepudi, K.K. Lactic Acid Bacterial Supplementation Ameliorated the Lipopolysaccharide-Induced Gut Inflammation and Dysbiosis in Mice. Front. Microbiol. 2022, 13, 930928. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Elshimy, R. Therapeutic effects of Stemmoside C against Salmonella enterica serotype typhimurium infected BALB/c mice. Steroids 2023, 199, 109296. [Google Scholar] [CrossRef]
- Xu, X.; Duarte, M.E.; Kim, S.W. Postbiotic effects of Lactobacillus fermentate on intestinal health, mucosa-associated microbiota, and growth efficiency of nursery pigs challenged with F18+ Escherichia coli. J. Anim. Sci. 2022, 100, skac210. [Google Scholar] [CrossRef]
- Li, Q.; Peng, X.; Burrough, E.R.; Sahin, O.; Gould, S.A.; Gabler, N.K.; Loving, C.L.; Dorman, K.S.; Patience, J.F. Dietary Soluble and Insoluble Fiber with or without Enzymes Altered the Intestinal Microbiota in Weaned Pigs Challenged with Enterotoxigenic E. coli F18. Front. Microbiol. 2020, 11, 1110. [Google Scholar] [CrossRef]
- Wang, D.; Zeng, J.; Wujin, C.; Ullah, Q.; Su, Z. Lactobacillus reuteri derived from horse alleviates Escherichia coli-induced diarrhea by modulating gut microbiota. Microb. Pathog. 2024, 188, 106541. [Google Scholar] [CrossRef]
- Peng, W.; He, C.; Li, R.; Qian, D.; Wang, L.; Chen, W.; Zhang, Q.; Wu, C. Zanthoxylum bungeanum amides ameliorates nonalcoholic fatty liver via regulating gut microbiota and activating AMPK/Nrf2 signaling. J. Ethnopharmacol. 2024, 318, 116848. [Google Scholar] [CrossRef]
- Tung, T.; Tung, Y.; Lin, I.; Shih, C.; Nguyen, N.T.K.; Shabrina, A.; Huang, S. Fish Oil, but Not Olive Oil, Ameliorates Depressive-Like Behavior and Gut Microbiota Dysbiosis in Rats under Chronic Mild Stress. Biomolecules 2019, 9, 516. [Google Scholar] [CrossRef]
- Zhang, J.S.; Li, S.; Cheng, X.; Tan, X.C.; Huang, Y.L.; Dong, H.J.; Xue, R.; Zhang, Y.; Li, J.C.; Feng, X.X.; et al. Far-Infrared Therapy Based on Graphene Ameliorates High-Fat Diet-Induced Anxiety-Like Behavior in Obese Mice via Alleviating Intestinal Barrier Damage and Neuroinflammation. Neurochem. Res. 2024, 49, 1735–1750. [Google Scholar] [CrossRef]
- Li, X.H.F.; Xin, T.C.J.; Hui, C.W.; Li, Y.X.; Zhi, F.D.; Rui, L.Y.; Xu, H.L. Irisin attenuates inflammation in a mouse model of ulcerative colitis by altering the intestinal microbiota. Exp. Ther. Med. 2021, 22, 1433. [Google Scholar]
- Zhang, J.; Zhao, Q.; Qin, Y.; Si, W.; Zhang, H.; Zhang, J. The Effect of Epimedium Isopentenyl Flavonoids on the Broiler Gut Health Using Microbiomic and Metabolomic Analyses. Int. J. Mol. Sci. 2023, 24, 7646. [Google Scholar] [CrossRef]
- Liu, W.; Xiao, M.; Yi, J.; Li, Y.; Kudinha, T.; Xu, Y. First case report of bacteremia caused by Solobacterium moorei in China, and literature review. BMC Infect. Dis. 2019, 19, 730. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Zhao, C.; Ashfaq, H.; Fakhar-E-Alam Kulyar, M.; Bhutta, Z.A.; Ali, M.M.; Mansoor, M.K.; Li, K. Cryptosporidium infection induced the dropping of SCFAS and dysbiosis in intestinal microbiome of Tibetan pigs. Microb. Pathog. 2023, 174, 105922. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Li, K.; Wang, P.; Huang, H.; Chen, M. Alleviating chronic kidney disease progression through modulating the critical genus of gut microbiota in a cisplatin-induced Lanyu pig model. J. Food Drug Anal. 2020, 28, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Dejean, G.; Tudela, H.; Bruno, L.; Kissi, D.; Rawadi, G.; Claus, S.P. Identifying a Novel Bile Salt Hydrolase from the Keystone Gut Bacterium Christensenella minuta. Microorganisms 2021, 9, 1252. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zuo, X.; Wang, X.; Shi, M.; Zhu, H.; Cao, C.; Liu, X.; Liang, W.; Yao, Y.; Wang, L. The probiotic Lactobacillus casei Zhang-mediated correction of gut dysbiosis ameliorates peritoneal fibrosis by suppressing macrophage-related inflammation via the butyrate/PPAR-γ/NF-κB pathway. Food Funct. 2023, 14, 6840–6852. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Q.; Lu, S.; Wang, J.; Xu, C.; Qu, W.; Nawaz, S.; Ataya, F.S.; Wu, Y.; Li, K. Lactobacillus salivarius and Berberine Alleviated Yak Calves’ Diarrhea via Accommodating Oxidation Resistance, Inflammatory Factors, and Intestinal Microbiota. Animals 2024, 14, 2419. https://doi.org/10.3390/ani14162419
He Q, Lu S, Wang J, Xu C, Qu W, Nawaz S, Ataya FS, Wu Y, Li K. Lactobacillus salivarius and Berberine Alleviated Yak Calves’ Diarrhea via Accommodating Oxidation Resistance, Inflammatory Factors, and Intestinal Microbiota. Animals. 2024; 14(16):2419. https://doi.org/10.3390/ani14162419
Chicago/Turabian StyleHe, Qing, Sijia Lu, Jia Wang, Chang Xu, Weijie Qu, Shah Nawaz, Farid Shokry Ataya, Yi Wu, and Kun Li. 2024. "Lactobacillus salivarius and Berberine Alleviated Yak Calves’ Diarrhea via Accommodating Oxidation Resistance, Inflammatory Factors, and Intestinal Microbiota" Animals 14, no. 16: 2419. https://doi.org/10.3390/ani14162419