

Towards Safe African Swine Fever Vaccines: The A137R Gene as a Tool to Reduce Virulence and a Promising Serological DIVA Marker Candidate

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Viruses
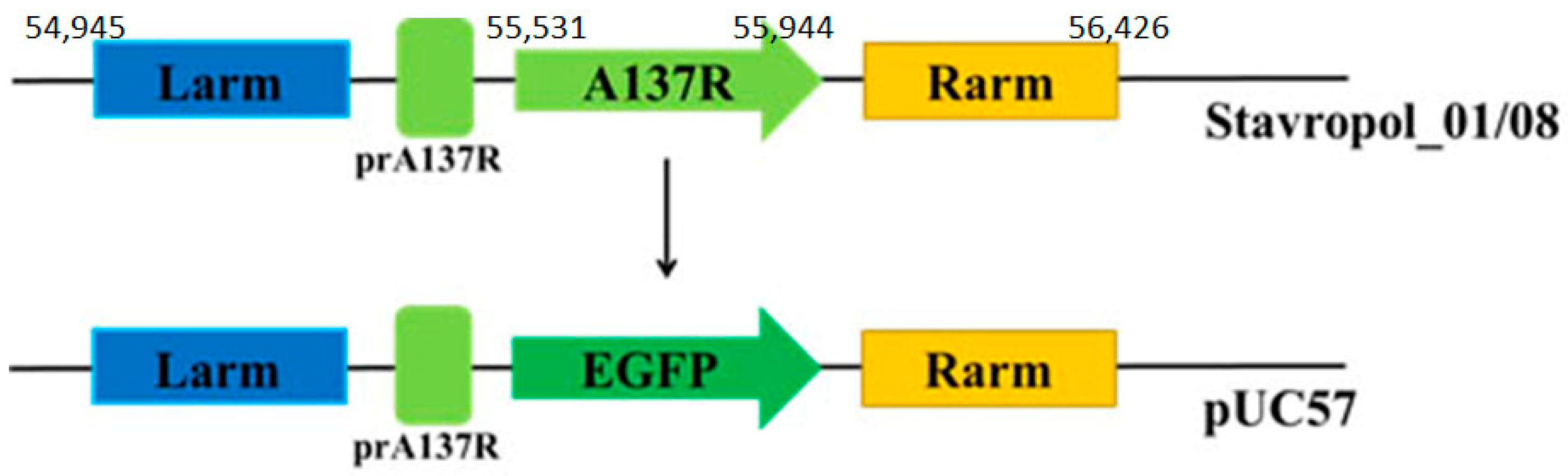
2.2. Construction of A137R-Deletion Mutant
2.3. Animal Experiments and Ethics Statement
2.4. ASFV pA137R-Coding Sequence Amplification, Cloning and Expression
2.5. Testing of Pig Sera by Indirect Immunofluorescence Assays and Western Blot
2.6. Hemadsorption Inhibition (HAI) Assays
2.7. Statistical Analysis
3. Results
3.1. Construction of A137R-deletion Mutant
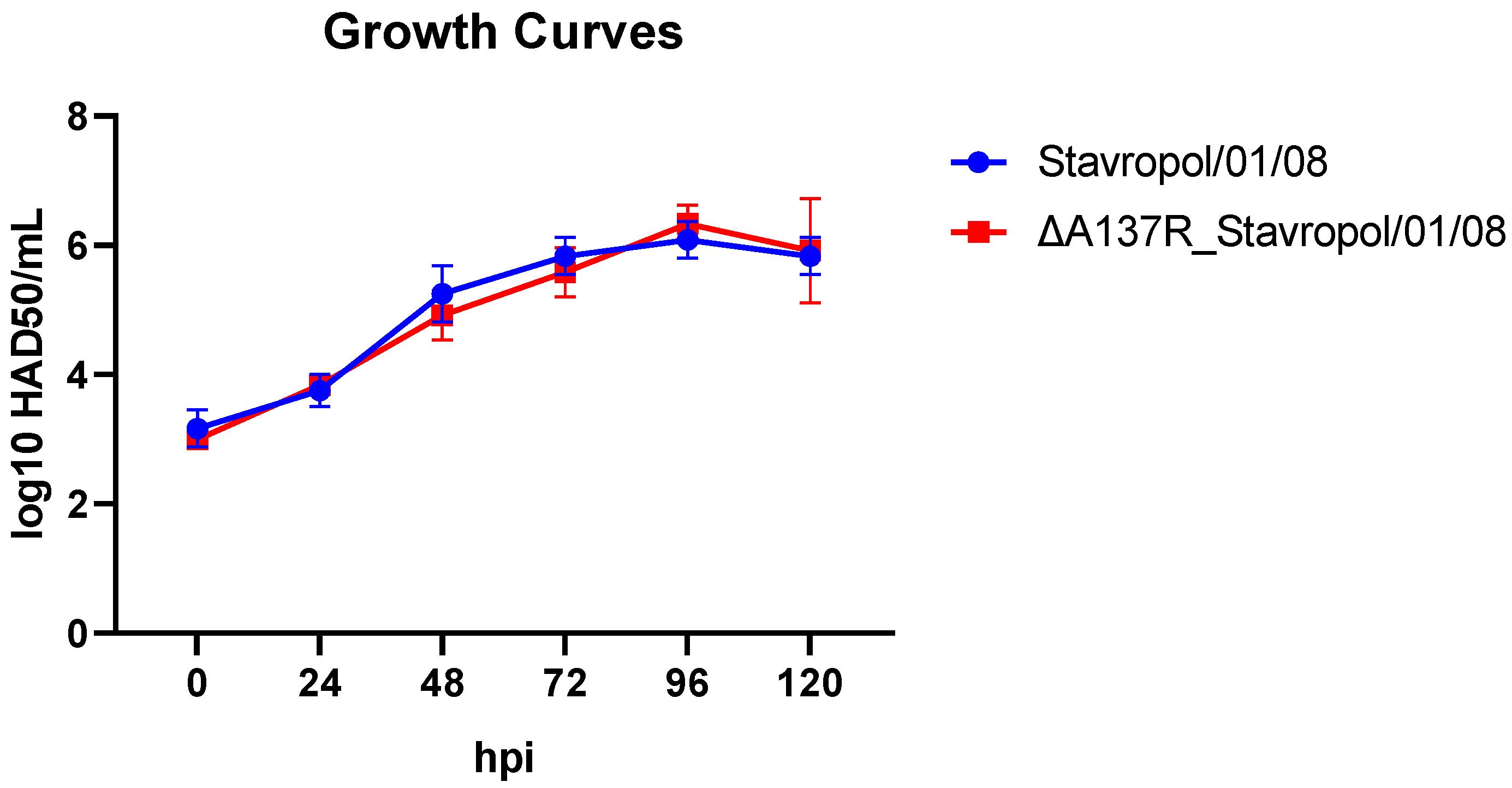
3.2. In Vitro Replication and Growth Kinetics of A137R-Deletion Mutant in Primary Cultures of Porcine Macrophages
3.3. Replication In Vivo and Virulence in Susceptible Animals of A137R-Deletion Mutant
3.4. ASFV p11.5 Protein Expression and Sera Sample Testing by Recombinant p11.5 Based IFA and Western Blot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eustace Montgomery, R. On a Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef]
- Coggins, L. African swine fever virus. Pathogenesis. Prog. Med. Virol. 1974, 18, 48–63. [Google Scholar] [PubMed]
- Mebus, C.A. African swine fever. Adv. Virus Res. 1988, 35, 251–269. [Google Scholar] [CrossRef]
- Dixon, L.K.; Chapman, D.A.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.D.; Penrith, M.L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, G.R. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Spinard, E.; Rai, A.; Osei-Bonsu, J.; O’Donnell, V.; Ababio, P.T.; Tawiah-Yingar, D.; Arthur, D.; Baah, D.; Ramirez-Medina, E.; Espinoza, N.; et al. The 2022 Outbreaks of African Swine Fever Virus Demonstrate the First Report of Genotype II in Ghana. Viruses 2023, 15, 1722. [Google Scholar] [CrossRef] [PubMed]
- Dinhobl, M.; Spinard, E.; Tesler, N.; Birtley, H.; Signore, A.; Ambagala, A.; Masembe, C.; Borca, M.V.; Gladue, D.P. Reclassification of ASFV into 7 Biotypes Using Unsupervised Machine Learning. Viruses 2024, 16, 67. [Google Scholar] [CrossRef]
- Spinard, E.; Dinhobl, M.; Fenster, J.; Davis, C.; Borca, M.V.; Gladue, D.P. Analysis of the Unique Historical Isolate of African Swine Fever Virus Isolate Spencer from Outbreaks in 1951. Viruses 2024, 16, 1175. [Google Scholar] [CrossRef]
- Thomson, G.R. The epidemiology of African swine fever: The role of free-living hosts in Africa. Onderstepoort J. Vet. Res. 1985, 52, 201–209. [Google Scholar] [PubMed]
- Jori, F.; Vial, L.; Penrith, M.L.; Pérez-Sánchez, R.; Etter, E.; Albina, E.; Michaud, V.; Roger, F. Review of the sylvatic cycle of African swine fever in sub-Saharan Africa and the Indian ocean. Virus Res. 2013, 173, 212–227. [Google Scholar] [CrossRef]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef]
- World Organisation for Animal Health: Home—WOAH. Available online: https://www.woah.org/en/disease/african-swine-fever/#ui-id-2 (accessed on 9 August 2024).
- Leitao, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.M.E.; Portugal, R.; Vigario, J.D.; Martins, C.L.V. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. J. Gen. Virol. 2001, 82, 513–523. [Google Scholar] [CrossRef] [PubMed]
- King, K.; Chapman, D.; Argilaguet, J.M.; Fishbourne, E.; Hutet, E.; Cariolet, R.; Hutchings, G.; Oura, C.A.L.; Netherton, C.L.; Moffat, K.; et al. Protection of European domestic pigs from virulent African isolates of African swine fever virus by experimental immunisation. Vaccine 2011, 29, 4593–4600. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Cordon, P.J.; Chapman, D.A.G.; Jabbar, T.; Reis, A.L.; Goatley, L.C.; Netherton, C.L.; Taylor, G.; Montoya, M.; Dixon, L.K. Different routes and doses influence protection in pigs immunised with the naturally attenuated African swine fever virus isolate OURT88/3. Antivir. Res. 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Vlasov, M.; Sindryakova, I.; Kudryashov, D.; Morgunov, S.; Kolbasova, O.; Lyska, V.; Zhivoderov, S.; Pivova, E.; Balyshev, V.; Namsrayn, S.; et al. Administration Routes and Doses of the Attenuated African Swine Fever Virus Strain PSA-1NH Influence Cross-Protection of Pigs against Heterologous Challenge. Animals 2024, 14, 1277. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.W.; Holinka, L.G.; O’Donnell, V.; Reese, B.; Sanford, B.; Fernandez-Sainz, I.; Gladue, D.P.; Arzt, J.; Rodriguez, L.; Risatti, G.R.; et al. The progressive adaptation of a Georgian isolate of African swine fever virus to Vero cells leads to a gradual attenuation of virulence in swine corresponding to major modifications of the viral genome. J. Virol. 2015, 89, 2324–2332. [Google Scholar] [CrossRef] [PubMed]
- Titov, I.; Burmakina, G.; Morgunov, Y.; Morgunov, S.; Koltsov, A.; Malogolovkin, A.; Kolbasov, D. Virulent strain of African swine fever virus eclipses its attenuated derivative after challenge. Arch. Virol. 2017, 162, 3081–3088. [Google Scholar] [CrossRef] [PubMed]
- Sereda, A.D.; Balyshev, V.M.; Kazakova, A.S.; Imatdinov, A.R.; Kolbasov, D.V. Protective Properties of Attenuated Strains of African Swine Fever Virus Belonging to Seroimmunotypes I–VIII. Pathogens 2020, 9, 274. [Google Scholar] [CrossRef]
- Sereda, A.D.; Vlasov, M.E.; Koltsova, G.S.; Morgunov, S.Y.; Kudrjashov, D.A.; Sindryakova, I.P.; Kolbasova, O.L.; Lyska, V.M.; Koltsov, A.Y.; Zhivoderov, S.P.; et al. Immunobiological Characteristics of the Attenuated African Swine Fever Virus Strain Katanga-350. Viruses 2022, 14, 1630. [Google Scholar] [CrossRef] [PubMed]
- Vlasov, M.E.; Sindryakova, I.P.; Kudrjashov, D.A.; Morgunov, S.Y.; Kolbasova, O.L.; Lyska, V.M.; Zhivoderov, S.P.; Pivova, E.Y.; Balyshev, V.M.; Sereda, A.D.; et al. Inoculation with ASFV-Katanga-350 Partially Protects Pigs from Death during Subsequent Infection with Heterologous Type ASFV-Stavropol 01/08. Viruses 2023, 15, 430. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African swine fever virus Georgia isolate harboring deletions of MGF360 and MGF505 genes is attenuated in swine and confers protection against challenge with virulent parental virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.; Risatti, G.R.; Holinka, L.G.; Krug, P.W.; Carlson, J.; Velazquez-Salinas, L.; Azzinaro, P.A.; Gladue, D.P.; Borca, M.V. Simultaneous deletion of the 9GL and UK genes from the African swine fever virus Georgia 2007 isolate offers increased safety and protection against homologous challenge. J. Virol. 2017, 91, e01760-16. [Google Scholar] [CrossRef]
- Monteagudo, P.L.; Lacasta, A.; López, E.; Bosch, L.; Collado, J.; Pina-Pedrero, S.; Correa-Fiz, F.; Accensi, F.; Navas, M.J.; Vidal, E.; et al. BA71DCD2: A new recombinant live attenuated African swine fever virus with cross-protective capabilities. J. Virol. 2017, 91, e01058-17. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a highly effective African swine fever virus vaccine by deletion of the I177L gene results in sterile immunity against the current epidemic Eurasia strain. J. Virol. 2020, 94, 10-1128. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Chen, T.; Yang, J.; Yue, H.; Wang, L.; Zhou, X.; Qi, Y.; Han, X.; Ke, J.; et al. Deletion of the L7L-L11L genes attenuates ASFV and induces protection against homologous challenge. Viruses 2021, 13, 255. [Google Scholar] [CrossRef]
- Balysheva, V.I.; Prudnikova, E.Y.; Galnbek, T.V.; Balyshev, V.M. Immunological properties of attenuated variants of African swine fever virus isolated in the Russian Federation. Russ. Agricult. Sci. 2015, 41, 178–182. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G. A proteomic atlas of the African swine fever virus particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of A137R gene from the pandemic strain of African swine fever virus is attenuated and offers protection against virulent pandemic virus. J. Virol. 2021, 95, e01139-21. [Google Scholar] [CrossRef]
- Kessler, C.; Forth, J.H.; Keil, G.M.; Mettenleiter, T.C.; Blome, S.; Karger, A. The intracellular proteome of African swine fever virus. Sci. Rep. 2018, 8, 14714. [Google Scholar] [CrossRef] [PubMed]
- Cackett, G.; Matelska, D.; Sykora, M.; Portugal, R.; Malecki, M.; Bahler, J.; Dixon, L.; Werner, F. The African swine fever virus transcriptome. J. Virol. 2020, 94, e00119-20. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Medina, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Pina-Pedrero, S.; Zhu, J.; Rodriguez, F.; Borca, M.V.; et al. Deletion of E184L, a putative DIVA target from the pandemic strain of African swine fever virus, produces a reduction in virulence and protection against virulent challenge. J. Virol. 2022, 96, e01419-21. [Google Scholar] [CrossRef] [PubMed]
- Hemmink, J.D.; Khazalwa, E.M.; Abkallo, H.M.; Oduor, B.; Khayumbi, J.; Svitek, N.; Henson, S.P.; Blome, S.; Keil, G.; Bishop, R.P.; et al. Deletion of the CD2v gene from the genome of ASFV-Kenya-IX-1033 partially reduces virulence and induces protection in pigs. Viruses 2022, 14, 1917. [Google Scholar] [CrossRef]
- Deutschmann, P.; Carrau, T.; Sehl-Ewert, J.; Forth, J.H.; Viaplana, E.; Mancera, J.C.; Urniza, A.; Beer, M.; Blome, S. Taking a promising vaccine candidate further: Efficacy of ASFV-G-ΔMGF after intramuscular vaccination of domestic pigs and oral vaccination of wild boar. Pathogens 2022, 11, 996. [Google Scholar] [CrossRef]
- Sun, M.; Yu, S.; Ge, H.; Wang, T.; Li, Y.; Zhou, P.; Pan, L.; Han, Y.; Yang, Y.; Sun, Y.; et al. The A137R protein of African swine fever virus inhibits type I interferon production via the autophagy-mediated lysosomal degradation of TBK1. J. Virol. 2022, 96, e01957-21. [Google Scholar] [CrossRef]
- Li, C.; Jia, M.; Hao, T.; Peng, Q.; Peng, R.; Chai, Y.; Shi, Y.; Song, H.; Gao, G.F. African swine fever virus A137R assembles into a dodecahedron cage. J. Virol. 2024, 98, e01536-23. [Google Scholar] [CrossRef]
- Yin, D.; Geng, R.; Lv, H.; Bao, C.; Shao, H.; Ye, J.; Qian, K.; Qin, A. Development of real-time PCR based on A137R gene for the detection of African swine fever virus. Front. Vet. Sci. 2021, 8, 753967. [Google Scholar] [CrossRef]
- Watanabe, M.; Kitamura, T.; Nagata, K.; Ikezawa, M.; Kameyama, K.; Masujin, K.; Kokuho, T. Development of a novel indirect ELISA for the serological diagnosis of African swine fever using p11.5 protein as a target antigen. Pathogens 2023, 12, 774. [Google Scholar] [CrossRef]
- Embury-Hyatt, C.; Moffat, E.; Zhmendak, D.; Erdelyan, C.N.G.; Collignon, B.; Goonewardene, K.; Ambagala, A.; Yang, M. Generation and characterization of a monoclonal antibody against an African swine fever virus protein encoded by the A137R gene. Front. Vet. Sci. 2023, 10, 1286906. [Google Scholar] [CrossRef]
- Malogolovkin, A.; Burmakina, G.; Tulman, E.R.; Delhon, G.; Diel, D.G.; Salnikov, N.; Kutish, G.F.; Kolbasov, D.; Rock, D.L. African swine fever virus CD2v and C-type lectin gene loci mediate serological specificity. J. Gen. Virol. 2015, 96, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Koltsova, G.; Koltsov, A.; Krutko, S.; Kholod, N.; Tulman, E.R.; Kolbasov, D. Growth kinetics and protective efficacy of attenuated ASFV strain Congo with deletion of the EP402 gene. Viruses 2021, 13, 1259. [Google Scholar] [CrossRef]
- Howey, E.B.; O’Donnell, V.; de Carvalho Ferreira, H.C.; Borca, M.V.; Arzt, J. Pathogenesis of highly virulent African swine fever virus in domestic pigs exposed via intraoropharyngeal, intranasopharyngeal, and intramuscular inoculation, and by direct contact with infected pigs. Virus Res. 2013, 178, 328–339. [Google Scholar] [CrossRef]
- Burmakina, G.; Malogolovkin, A.; Tulman, E.R.; Zsak, L.; Delhon, G.; Diel, D.G.; Shobogorov, N.M.; Morgunov, Y.P.; Morgunov, S.Y.; Kutish, G.F.; et al. African swine fever virus serotype-specific proteins are significant protective antigens for African swine fever. J. Gen. Virol. 2016, 97, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Ruíz-Gonzalvo, F.; Rodríguez, F.; Escribano, J.M. Functional and immunological properties of the baculovirus-expressed hemagglutinin of African swine fever virus. Virology 1996, 218, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Espinoza, N.; Rai, A.; Spinard, E.; Velazquez-Salinas, L.; Valladares, A.; Silva, E.; Burton, L.; Meyers, A.; et al. Deletion of the EP402R gene from the genome of African swine fever vaccine strain ASFV-G-ΔI177L provides the potential capability of differentiating between infected and vaccinated animals. Viruses 2024, 16, 376. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African swine fever virus bearing an I226R gene deletion elicits robust immunity in pigs to African swine fever. J. Virol. 2021, 95, e01199-21. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No of Animals | Mortality | Fever | Result of ELISA | Viral Load in Blood (Max) | Viral Load in Organs (Max) | ||
|---|---|---|---|---|---|---|---|---|
| % | TTD | % | TTF | % | Genome Copies/mL | Genome Copies/mL | ||
| ΔA137R_Stavropol_01/08 | 8 | 87.5 | 10.7 | 100 | 3.875 | 12.5 | 1.64 × 108 | 5.12 × 109 |
| Stavropol_01/08 | 4 | 100 | 7.5 | 100 | 4 | 0 | 1.07 × 109 | 2.56 × 1010 |
| Samples | Positive Results * | ||
|---|---|---|---|
| IFA | WB | ELISA | |
| The reference anti-ASFV hyperimmune sera | 4/4 | 4/4 | 4/4 |
| Positive porcine serum samples collected at 14–28 dpi with the ASFV attenuated strains encoding p11.5 | 18/18 | 10/10 | 18/18 |
| Inconclusive porcine serum samples collected at 14–28 dpi with the ASFV attenuated strains encoding p11.5 | 6/6 | 4/4 | 0/6 |
| Porcine serum samples collected at 14–28 dpi with the ASFV A137R-deleted mutants | 0/6 | 0/6 | 6/6 |
| Porcine negative sera | 0/20 | 0/4 | 0/20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koltsov, A.; Sukher, M.; Krutko, S.; Belov, S.; Korotin, A.; Rudakova, S.; Morgunov, S.; Koltsova, G. Towards Safe African Swine Fever Vaccines: The A137R Gene as a Tool to Reduce Virulence and a Promising Serological DIVA Marker Candidate. Animals 2024, 14, 2469. https://doi.org/10.3390/ani14172469
Koltsov A, Sukher M, Krutko S, Belov S, Korotin A, Rudakova S, Morgunov S, Koltsova G. Towards Safe African Swine Fever Vaccines: The A137R Gene as a Tool to Reduce Virulence and a Promising Serological DIVA Marker Candidate. Animals. 2024; 14(17):2469. https://doi.org/10.3390/ani14172469
Chicago/Turabian StyleKoltsov, Andrey, Mikhail Sukher, Sergey Krutko, Sergey Belov, Alexey Korotin, Sofia Rudakova, Sergey Morgunov, and Galina Koltsova. 2024. "Towards Safe African Swine Fever Vaccines: The A137R Gene as a Tool to Reduce Virulence and a Promising Serological DIVA Marker Candidate" Animals 14, no. 17: 2469. https://doi.org/10.3390/ani14172469
APA StyleKoltsov, A., Sukher, M., Krutko, S., Belov, S., Korotin, A., Rudakova, S., Morgunov, S., & Koltsova, G. (2024). Towards Safe African Swine Fever Vaccines: The A137R Gene as a Tool to Reduce Virulence and a Promising Serological DIVA Marker Candidate. Animals, 14(17), 2469. https://doi.org/10.3390/ani14172469