

Modelling the Distribution and Habitat Suitability of the European Wildcat (Felis silvestris) in North-Western Spain and Its Conservation Implications
 , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
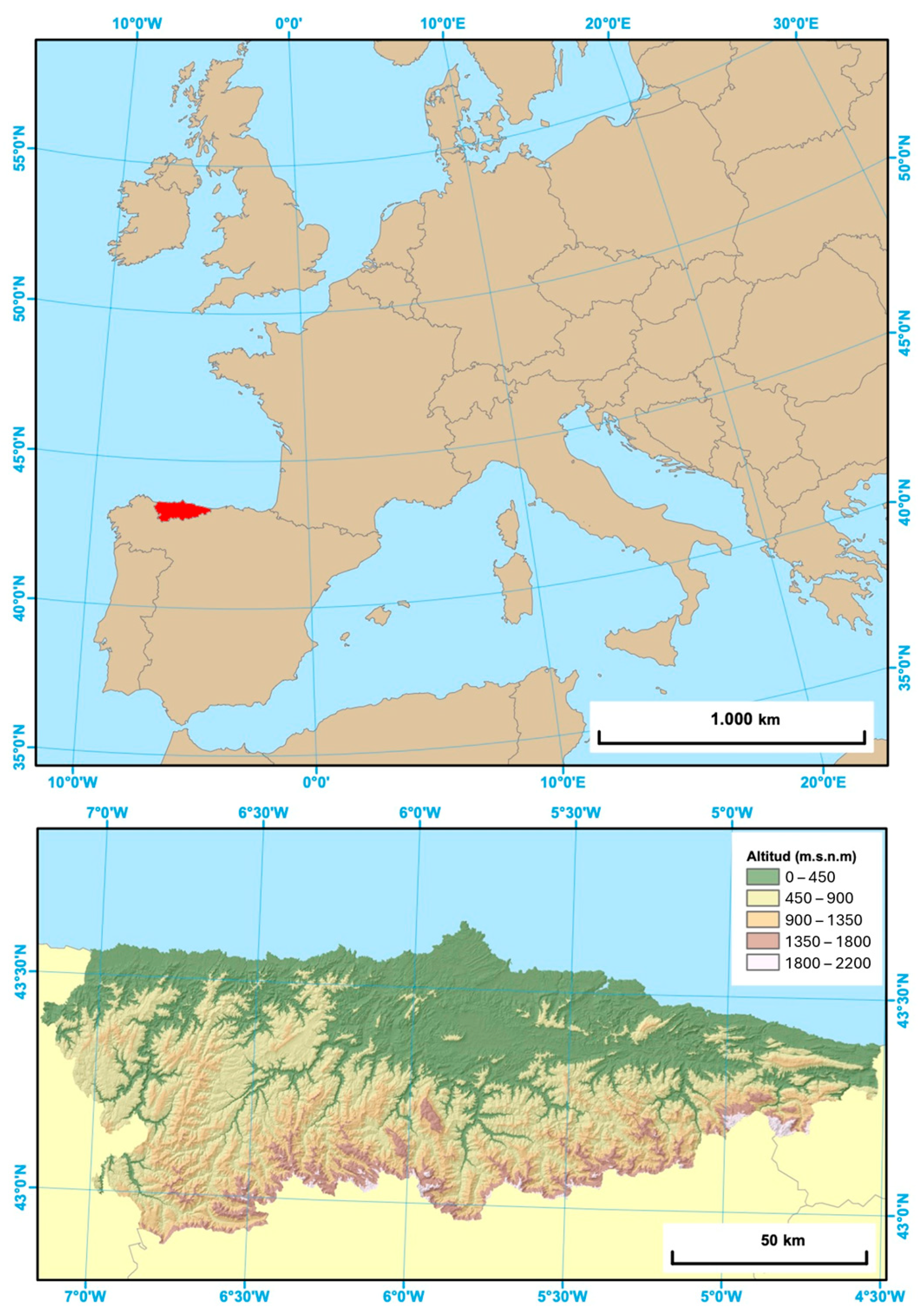
2.1. Study Area
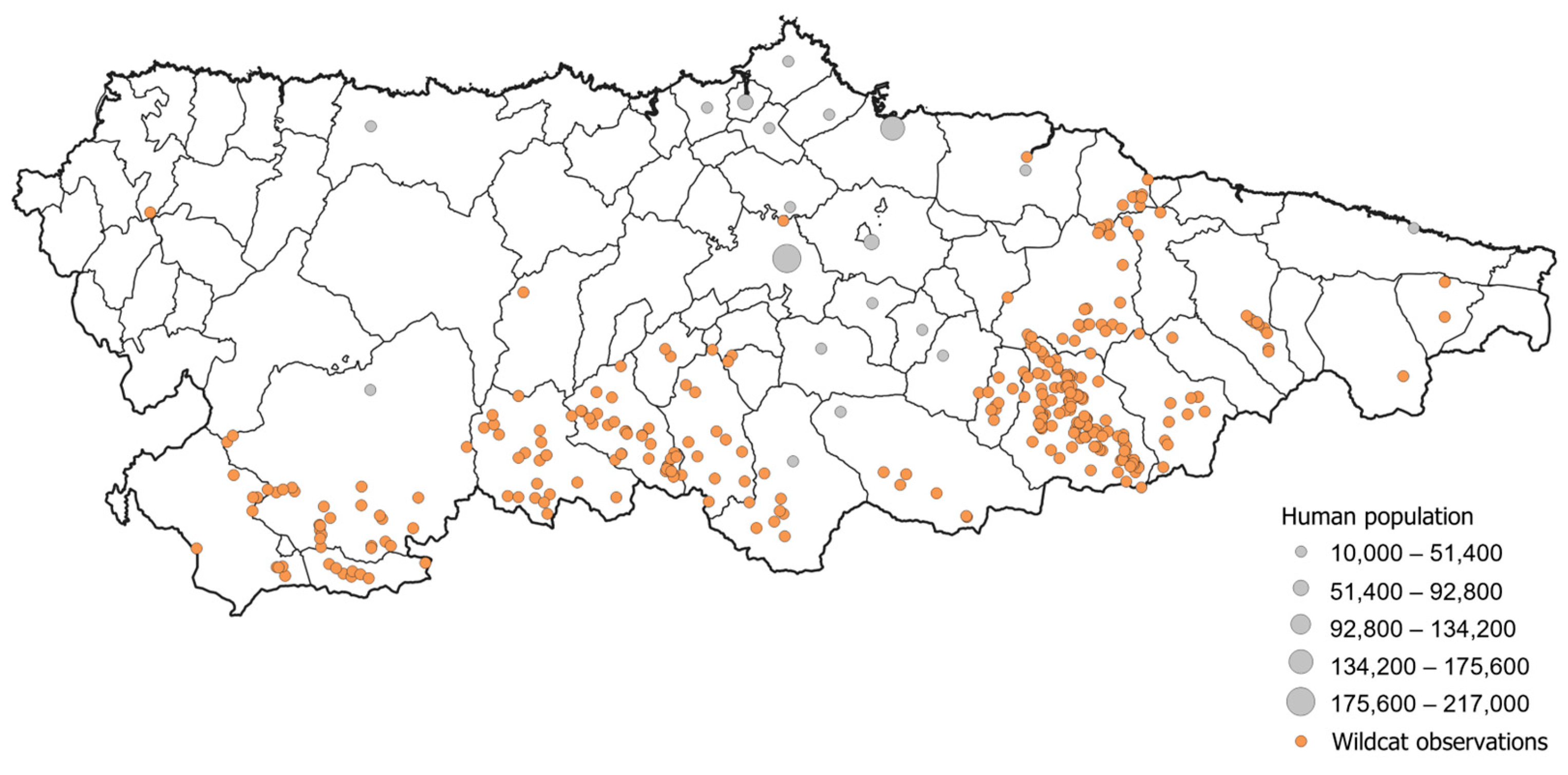
2.2. Wildcat Occurrence Data, Definition of Wildcat Range, and Characterisation of Suitable Habitats Available
2.3. Environmental Variables
2.4. MaxEnt Modelling
2.5. Selecting and Evaluating Models and Variable Contributions
2.6. Model Output
3. Results
3.1. Maxent Model Selection and Evaluation
3.2. Predicted Range Distribution and Potential Expansion Areas
3.3. Environmental Variable Contributions to the Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davoli, M.; Ghoddousi, A.; Sabatini, F.M.; Fabbri, E.; Caniglia, R.; Kuemmerle, T. Changing patterns of conflict between humans, carnivores and crop-raiding prey as large carnivores recolonize human-dominated landscapes. Biol. Conserv. 2022, 269, 109553. [Google Scholar] [CrossRef]
- Bombieri, G.; Penteriani, V.; Almasieh, K.; Ambarlı, H.; Ashrafzadeh, M.R.; Das, C.S.; Dharaiya, N.; Hoogesteijn, R.; Hoogesteijn, A.; Ikanda, D.; et al. A worldwide perspective on large carnivore attacks on humans. PLOS Biol. 2023, 21, e3001946. [Google Scholar] [CrossRef] [PubMed]
- Krofel, M.; Berce, M.; Berce, T.; Kryštufek, B.; Lamut, S.; Tarman, J.; Fležar, U. New mesocarnivore at the doorstep of Central Europe: Historic development of golden jackal (Canis aureus) population in Slovenia. Mammal Res. 2023, 68, 329–339. [Google Scholar] [CrossRef]
- Díaz-Ruiz, F.; Descalzo, E.; Martínez-Jauregui, M.; Soliño, M.; Márquez, A.L.; Farfán, M.Á.; Real, R.; Ferreras, P.; Delibes-Mateos, M. Combining ranger records and biogeographical models to identify the current and potential distribution of an expanding mesocarnivore in southern Europe. Sci. Total Environ. 2024, 946, 174216. [Google Scholar] [CrossRef] [PubMed]
- Benson, J.F.; Mahoney, P.J.; Sikich, J.A.; Serieys, L.E.K.; Pollinger, J.P.; Ernest, H.B.; Riley, S.P.D. Interactions between demography, genetics, and landscape connectivity increase extinction probability for a small population of large carnivores in a major metropolitan area. Proc. R. Soc. B Biol. Sci. 2016, 283, 10. [Google Scholar] [CrossRef]
- Boudreau, M.R.; Gantchoff, M.G.; Ramirez-Reyes, C.; Conlee, L.; Belant, J.L.; Iglay, R.B. Using habitat suitability and landscape connectivity in the spatial prioritization of public outreach and management during carnivore recolonization. J. Appl. Ecol. 2022, 59, 757–767. [Google Scholar] [CrossRef]
- Planillo, A.; Wenzler-Meya, M.; Reinhardt, I.; Kluth, G.; Michler, F.U.; Stier, N.; Louvrier, J.; Steyer, K.; Gillich, B.; Rieger, S.; et al. Understanding habitat selection of range-expanding populations of large carnivores: 20 years of grey wolves (Canis lupus) recolonizing Germany. Divers. Distrib. 2024, 30, 71–86. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Zarzo-Arias, A.; Penteriani, V.; del Mar Delgado, M.; Torre, P.P.; García-González, R.; Mateo-Sánchez, M.C.; García, P.V.; Dalerum, F. Identifying potential areas of expansion for the endangered brown bear (Ursus arctos) population in the Cantabrian Mountains (NW Spain). PLoS ONE 2019, 14, e0209972. [Google Scholar] [CrossRef]
- Su, H.; Bista, M.; Li, M. Mapping habitat suitability for Asiatic black bear and red panda in Makalu Barun National Park of Nepal from Maxent and GARP models. Sci. Rep. 2021, 11, 14135. [Google Scholar] [CrossRef]
- Schmidt, K.; Górny, M.; Jędrzejewski, W. Effect of microhabitat characteristics for predicting habitat suitability for a stalking large carnivore—The Eurasian lynx in middle Europe. Anim. Conserv. 2023, 26, 851–864. [Google Scholar] [CrossRef]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; IUCN Gland: Gland, Switzerland, 1996; Volume 382. [Google Scholar]
- Macdonald, D.W. The Scottish Wildcat: Analyses for Conservation and an Action Plan; Wildlife Conservation Research Unit: Oxford, UK, 2004. [Google Scholar]
- Bastianelli, M.L.; Premier, J.; Herrmann, M.; Anile, S.; Monterroso, P.; Kuemmerle, T.; Dormann, C.F.; Streif, S.; Jerosch, S.; Götz, M.; et al. Survival and cause-specific mortality of European wildcat (Felis silvestris) across Europe. Biol. Conserv. 2021, 261, 109239. [Google Scholar] [CrossRef]
- Gil-Sánchez, J.M.; Barea-Azcón, J.M.; Jaramillo, J.; Herrera-Sánchez, F.J.; Jiménez, J.; Virgós, E. Fragmentation and low density as major conservation challenges for the southernmost populations of the European wildcat. PLoS ONE 2020, 15, e0227708. [Google Scholar] [CrossRef] [PubMed]
- Slotta-Bachmayr, L.; Meikl, M.; Hagenstein, I. Current status of the European wildcat (Felis silvestris silvestris, Schreber, 1777) in Austria. Acta ZooBot Austria 2016, 153, 68–76. [Google Scholar]
- Gerngross, P.; Ambarli, H.; Angelici, F.M.; Anile, S.; Campbell, R.; Ferreras de Andres, P.; Gil-Sanchez, J.M.; Götz, M.; Jerosch, S.; Mengüllüoglu, D.; et al. Felis silvestris (Amended Version of 2022 Assessment). The IUCN Red List of Threatened Species 2023: e.T181049859A224982454. 2023. Available online: https://www.worldclim.org.
- Steyer, K.; Tiesmeyer, A.; Muñoz-Fuentes, V.; Nowak, C. Low rates of hybridization between European wildcats and domestic cats in a human-dominated landscape. Ecol. Evol. 2018, 8, 2290–2304. [Google Scholar] [CrossRef] [PubMed]
- Migli, D.; Astaras, C.; Boutsis, G.; Diakou, A.; Karantanis, N.E.; Youlatos, D. Spatial ecology and diel activity of European Wildcat (Felis silvestris) in a protected lowland area in Northern Greece. Animals 2021, 11, 3030. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Villar, H.; Bastianelli, M.L.; Heurich, M.; Anile, S.; Díaz-Ruiz, F.; Ferreras, P.; Götz, M.; Herrmann, M.; Jerosch, S.; Jubete, F.; et al. Agriculture intensity and landscape configuration influence the spatial use of wildcats across Europe. Biol. Conserv. 2023, 277, 109854. [Google Scholar] [CrossRef]
- Monterroso, P.; Célio Alves, P.; Ferreras, P. Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: Implications for species coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef]
- Martín-Díaz, P.; Gil-Sánchez, J.M.; Balleseros-Duperón, E.; Barea-Azcón, J.M.; Virgós, E.; Pardavila, X.; Moleón, M. Integrating space and time in predator-prey studies: The case of wildcats and rabbits in SE Spain. Mamm. Biol. 2018, 88, 114–122. [Google Scholar] [CrossRef]
- Jiménez-Albarral, J.J.; Urra, F.; Jubete, F.; Roman, J.; Revilla, E.; Palomares, F. Abundance and use pattern of wildcats of ancient human-modified cattle pastures in northern Iberian Peninsula. Eur. J. Wildl. Res. 2021, 67, 94. [Google Scholar] [CrossRef]
- Ruiz-Villar, H.; Jubete, F.; Revilla, E.; Román, J.; Urra, F.; López-Bao, J.V.; Palomares, F. Like cat and fox: Diurnal interactions between two sympatric carnivores in pastoral landscapes of NW Spain. Eur. J. Wildl. Res. 2021, 67, 16. [Google Scholar] [CrossRef]
- Ruiz-Villar, H.; Morales-González, A.; López-Bao, J.V.; Palomares, F. Humans and traffic influence European wildcat behaviour in pastoral landscapes. Anim. Behav. 2024, 207, 131–146. [Google Scholar] [CrossRef]
- Széles, G.L.; Purger, J.J.; Molnár, T.; Lanszki, J. Comparative analysis of the diet of feral and house cats and wildcat in Europe. Mammal Res. 2018, 63, 43–53. [Google Scholar] [CrossRef]
- Soto, C.A.; Palomares, F. Surprising low abundance of European wildcats in a mediterranean protected area of Southwestern Spain. Mammalia 2014, 78, 57–65. [Google Scholar] [CrossRef]
- Lozano, J.; Virgós, E.; Cabezas-Díaz, S. Monitoring European wildcat Felis silvestris populations using scat surveys in central Spain: Are population trends related to wild rabbit dynamics or to landscape features? Zool. Stud. 2013, 52, 16. [Google Scholar] [CrossRef]
- Quevedo, M.; Echegaray, J.; Fernández-Gil, A.; Leonard, J.A.; Naves, J.; Ordiz, A.; Revilla, E.; Vilà, C. Lethal management may hinder population recovery in Iberian wolves. Biodivers. Conserv. 2019, 28, 415–432. [Google Scholar] [CrossRef]
- Martínez Cano, I.; Taboada, F.G.; Naves, J.; Fernández-Gil, A.; Wiegand, T. Decline and recovery of a large carnivore: Environmental change and long-term trends in an endangered brown bear population. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161832. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Lozano, J. Gato montés—Felis silvestris Schreber, 1777. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2017; p. 28. Available online: https://www.vertebradosibericos.org/mamiferos/felsil.html (accessed on 1 September 2024).
- Eriksson, T.; Dalerum, F. Identifying potential areas for an expanding wolf population in Sweden. Biol. Conserv. 2018, 220, 170–181. [Google Scholar] [CrossRef]
- Lozano, J. Habitat use by European wildcats (Felis silvestris) in central Spain: What is the relative importance of forest variables? Anim. Biodivers. Conserv. 2010, 33, 143–150. [Google Scholar] [CrossRef]
- Carreras-Duro, J.; Moleón, M.; Barea-Azcón, J.M.; Ballesteros-Duperón, E.; Virgós, E. Optimization of sampling effort in carnivore surveys based on signs: A regional-scale study in a Mediterranean area. Mamm. Biol.-Z. Saugetierkd. 2016, 81, 205–213. [Google Scholar] [CrossRef]
- Cushman, S.A.; Kilshaw, K.; Kaszta, Z.; Campbell, R.D.; Gaywood, M.; Macdonald, D.W. Variable importance and scale of influence across individual scottish wildcat hybrid habitat models. Ecol. Model. 2024, 491, 110698. [Google Scholar] [CrossRef]
- Garcia-Perea, R. Felis silvestris Schreber, 1777. In Atlas y Libro Rojo de los Mamıferos Terrestres de España; Dirección General para la Biodiversidad–SECEM–SECEMU: Madrid, Spain, 2007; pp. 333–335. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E.; Maxent Software for Modeling Species Niches and Distributions. In (Version 3.4.0). 2017. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent (accessed on 1 September 2024).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.R.; Elith, J. Package ‘dismo’. Circles 2017, 9, 55. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.T.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Young, N.; Carter, L.; Evangelista, P. A MaxEnt Model v3. 3.3 e Tutorial (ArcGIS v10); Natural Resource Ecology Laboratory, Colorado State University and the National Institute of Invasive Species Science: Fort Collins, CO, USA, 2011. [Google Scholar]
- Klar, N.; Fernández, N.; Kramer-Schadt, S.; Herrmann, M.; Trinzen, M.; Büttner, I.; Niemitz, C. Habitat selection models for European wildcat conservation. Biol. Conserv. 2008, 141, 308–319. [Google Scholar] [CrossRef]
- Virgós, E.; Travaini, A. Relationship between small-game hunting and carnivore diversity in Central Spain. Biodivers. Conserv. 2005, 14, 3475–3486. [Google Scholar] [CrossRef]
- Cano, C.; Ayerza, P.; de la Hoz, J.F. El Veneno en España (1990–2005). Análisis del Problema, Incidencia y Causas. Propuestas de WWF/Adena; WWF/Adena: Madrid, Spain, 2006. [Google Scholar]
- Urra, F. El Gato Montés en Navarra: Distribución, Ecología y Conservación; Universidad Autónoma de Madrid: Madrid, Spain, 2003. [Google Scholar]
- Baker, A.D.; Leberg, P.L. Impacts of human recreation on carnivores in protected areas. PLoS ONE 2018, 13, e0195436. [Google Scholar] [CrossRef] [PubMed]
- Bandak, S.; Sarno, R.; Peterson, M.; Farkas, D.; Grigione, M. Active humans, inactive carnivores, and hiking trails within a suburban preserve. Suburb. Sustain. 2020, 6, 1. [Google Scholar] [CrossRef]
- Liberek, M. Eco-ethologie du chat sauvage Felis s. silvestris, Schreber 1777 dans le Jura Vaudois (Suisse): Influence de la Couverture Neigeuse. Ph.D. Thesis, Université de Neuchâtel, Neuchâtel, Switzerland, 1999. [Google Scholar]
- Mermod, C.P.; Liberek, M. The role of snowcover for European wildcat in Switzerland. Z. Jagdwiss. 2002, 48, 17–24. [Google Scholar] [CrossRef]
- Lastrada, E.; Garzón-Roca, J.; Cobos, G.; Torrijo, F.J. A decrease in the regulatory effect of snow-related phenomena in spanish mountain areas due to climate change. Water 2021, 13, 1550. [Google Scholar] [CrossRef]
- Parent, G.H. La migration récente, à caractère invasionnel, du chat sauvage, Felis silvestris silvestris Schreber, en Lorraine Belge. Mammalia 1975, 39, 251–288. [Google Scholar] [CrossRef]
- Piechocki, R. Die Wildkatze: Felis silvestris Wolf, Verlagskg; Ziemsen: Reykjavik, Iceland, 1990. [Google Scholar]
- Stahl, P. Le Chat Forestier d’Europe (Felis silvestris, Schreber 1777): Exploitation des Ressources et Organisation Spatiale. Ph.D. Thesis, Universitè Nancy, Nancy, France, 1986. [Google Scholar]
- Wittmer, H.U. Home range size, movements, and habitat utilization of three male European wildcats (Felis silvestris Schreber, 1777) in Saarland and Rheinland-Pfalz (Germany). Mamm. Biol. 2001, 66, 365–370. [Google Scholar]
- Silva, A.P.; Kilshaw, K.; Johnson, P.J.; MacDonald, D.W.; Rosalino, L.M. Wildcat occurrence in Scotland: Food really matters. Divers. Distrib. 2013, 19, 232–243. [Google Scholar] [CrossRef]
- Osbourne, J.D.; Anderson, J.T.; Spurgeon, A.B. Effects of habitat on small-mammal diversity and abundance in West Virginia. Wildl. Soc. Bull. 2005, 33, 814–822. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Plant and small mammal diversity in orchard versus non-crop habitats. Agric. Ecosyst. Environ. 2006, 116, 235–243. [Google Scholar] [CrossRef]
- López-Martín, J.M.; García, F.J.; Duch, A.; Virgós, E.; Lozano, J.; Duarte, J.; España, A.J. Felis silvestris Schreber, 1777. In Atlas y Libro Rojo de los Mamíferos de España; Dirección General para la Biodiversidad-SECEM -SECEMU: Madrid, Spain, 2007; pp. 336–338. [Google Scholar]
- Vila Subirós, J.; Rodríguez-Carreras, R.; Varga, D.; Ribas, A.; Úbeda, X.; Asperó, F.; Llausàs, A.; Outeiro, L. Stakeholder perceptions of landscape changes in the Mediterranean mountains of the North-Eastern Iberian Peninsula. Land Degrad. Dev. 2016, 27, 1354–1365. [Google Scholar] [CrossRef]
- Regos, A.; Domínguez, J.; Gil-Tena, A.; Brotons, L.; Ninyerola, M.; Pons, X. Rural abandoned landscapes and bird assemblages: Winners and losers in the rewilding of a marginal mountain area (NW Spain). Reg. Environ. Chang. 2016, 16, 199–211. [Google Scholar] [CrossRef]
- Martínez-Abraín, A.; Jiménez, J.; Jiménez, I.; Ferrer, X.; Llaneza, L.; Ferrer, M.; Palomera, G.; Ballesteros, F.; Galán, P.; Oro, D. Ecological consequences of human depopulation of rural areas on wildlife: A unifying perspective. Biol. Conserv. 2020, 252, 108860. [Google Scholar] [CrossRef]
- Oliveira, T.; Urra, F.; López-Martín, J.M.; Ballesteros-Duperón, E.; Barea-Azcón, J.M.; Moléon, M.; Gil-Sánchez, J.M.; Alves, P.C.; Díaz-Ruíz, F.; Ferreras, P.; et al. Females know better: Sex-biased habitat selection by the European wildcat. Ecol. Evol. 2018, 8, 9464–9477. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.; Godinho, R.; Randi, E.; Alves, P.C. Hybridization versus conservation: Are domestic cats threatening the genetic integrity of wildcats (Felis silvestris silvestris) in Iberian Peninsula? Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2953–2961. [Google Scholar] [CrossRef]
- Gil-Sánchez, J.M.; Jaramillo, J.; Barea-Azcón, J.M. Strong spatial segregation between wildcats and domestic cats may explain low hybridization rates on the Iberian Peninsula. Zoology 2015, 118, 377–385. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Name | Description | Source | Format | Year |
|---|---|---|---|---|
| Highways * | Length highways | BCN200 | Vector | 2015 |
| Roads * | Length autonomic and national roads | BCN200 | Vector | 2015 |
| Footpaths * | Length footpaths | BCN200 | Vector | 2015 |
| Rivers * | Length rivers | BCN200 | Vector | 2015 |
| Elevation and slope * | MDT200 | Raster | 2017 | |
| NDVI * | Normalised Difference Vegetation Index | Instituto de Recursos Naturales y Ordenación del Territorio (INDUROT) | Raster | 2011 |
| Land cover * | Percentage of each class per grid: cliffs, farmlands, fern, forests, gorse, heath, human structures, open grounds, pastures, forest plantations, shrublands, water, wetlands; n° classes, and Shannon index | Cartografía Temática Ambiental del Principado de Asturias 1989–1998 (1:50,000) | Vector | 1998 |
| Human density * | SADEI nomenclator | Vector | 2015 | |
| Bio 1 | Annual mean temperature | Worldclim | Raster | 2000 |
| Bio 5 | Max temperature of warmest month | Worldclim | Raster | 2000 |
| Bio 6 | Min temperature of coldest month | Worldclim | Raster | 2000 |
| Bio 7 | Temperature annual range (BIO5–BIO6) | Worldclim | Raster | 2000 |
| Bio 12 | Annual precipitation | Worldclim | Raster | 2000 |
| Bio 15 * | Precipitation seasonality (Coefficient of variation) | Worldclim | Raster | 2000 |
| Feature Types a | Regularisation Multiplier | Full AUC | Mean AUC | AUC Diff | OR Min | AICc | Δ AICc | Nparam |
|---|---|---|---|---|---|---|---|---|
| LQT | 3.5 | 0.840 | 0.750 | 0.093 | 0.128 | 1777.99 | 0 | 26 |
| LQHT | 1.5 | 0.870 | 0.792 | 0.094 | 0.085 | 1779.50 | 1.511 | 35 |
| LQ | 2 | 0.833 | 0.752 | 0.096 | 0.133 | 1781.71 | 3.718 | 25 |
| LQ | 4 | 0.830 | 0.746 | 0.092 | 0.128 | 1784.11 | 6.122 | 23 |
| LQT | 4.5 | 0.832 | 0.745 | 0.091 | 0.128 | 1784.53 | 6.548 | 24 |
| Variable | Percentage Contribution |
|---|---|
| Elevation | 39 |
| Percent of forest | 26.6 |
| Footpaths | 15.9 |
| Percent of forest plantations | 7.6 |
| Slope | 5.2 |
| Precipitation seasonality | 1.5 |
| Percent of fern | 1.4 |
| Percent of shrubland | 0.9 |
| River density | 0.8 |
| Number of landcover classes | 0.8 |
| Percent of heath | 0.2 |
| Percent of pasture | 0.1 |
| Shannon index | 0.1 |
| Roads | 0 |
| Percent of human structures | 0 |
| Percent of gorse | 0 |
| Percent of open ground | 0 |
| NDVI | 0 |
| Human density | 0 |
| Highways | 0 |
| Percent of water | 0 |
| Percent of wetland | 0 |
| Percent of farmlands | 0 |
| Percent of cliffs | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez García, P.; Zarzo-Arias, A.; Vigón Álvarez, E.; Alambiaga, I.; Monrós, J.S. Modelling the Distribution and Habitat Suitability of the European Wildcat (Felis silvestris) in North-Western Spain and Its Conservation Implications. Animals 2024, 14, 2708. https://doi.org/10.3390/ani14182708
Vázquez García P, Zarzo-Arias A, Vigón Álvarez E, Alambiaga I, Monrós JS. Modelling the Distribution and Habitat Suitability of the European Wildcat (Felis silvestris) in North-Western Spain and Its Conservation Implications. Animals. 2024; 14(18):2708. https://doi.org/10.3390/ani14182708
Chicago/Turabian StyleVázquez García, Pablo, Alejandra Zarzo-Arias, Efrén Vigón Álvarez, Iván Alambiaga, and Juan S. Monrós. 2024. "Modelling the Distribution and Habitat Suitability of the European Wildcat (Felis silvestris) in North-Western Spain and Its Conservation Implications" Animals 14, no. 18: 2708. https://doi.org/10.3390/ani14182708
APA StyleVázquez García, P., Zarzo-Arias, A., Vigón Álvarez, E., Alambiaga, I., & Monrós, J. S. (2024). Modelling the Distribution and Habitat Suitability of the European Wildcat (Felis silvestris) in North-Western Spain and Its Conservation Implications. Animals, 14(18), 2708. https://doi.org/10.3390/ani14182708