
Muscle Proteins, Technological Properties, and Free Amino Acids of Epaxial Muscle Collected from Asian Seabass (Lates calcarifer) at Different Postmortem Durations

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Sample Collection
2.2. Myofibril Fragmentation Index
2.3. Muscle Protein Composition
2.4. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.5. Thawing and Cooking Loss
2.6. Surface Color
2.7. pH
2.8. Expressible Water
2.9. Texture Profile Analysis (TPA)
2.10. Free Amino Acids
2.11. Statistical Analysis
3. Results
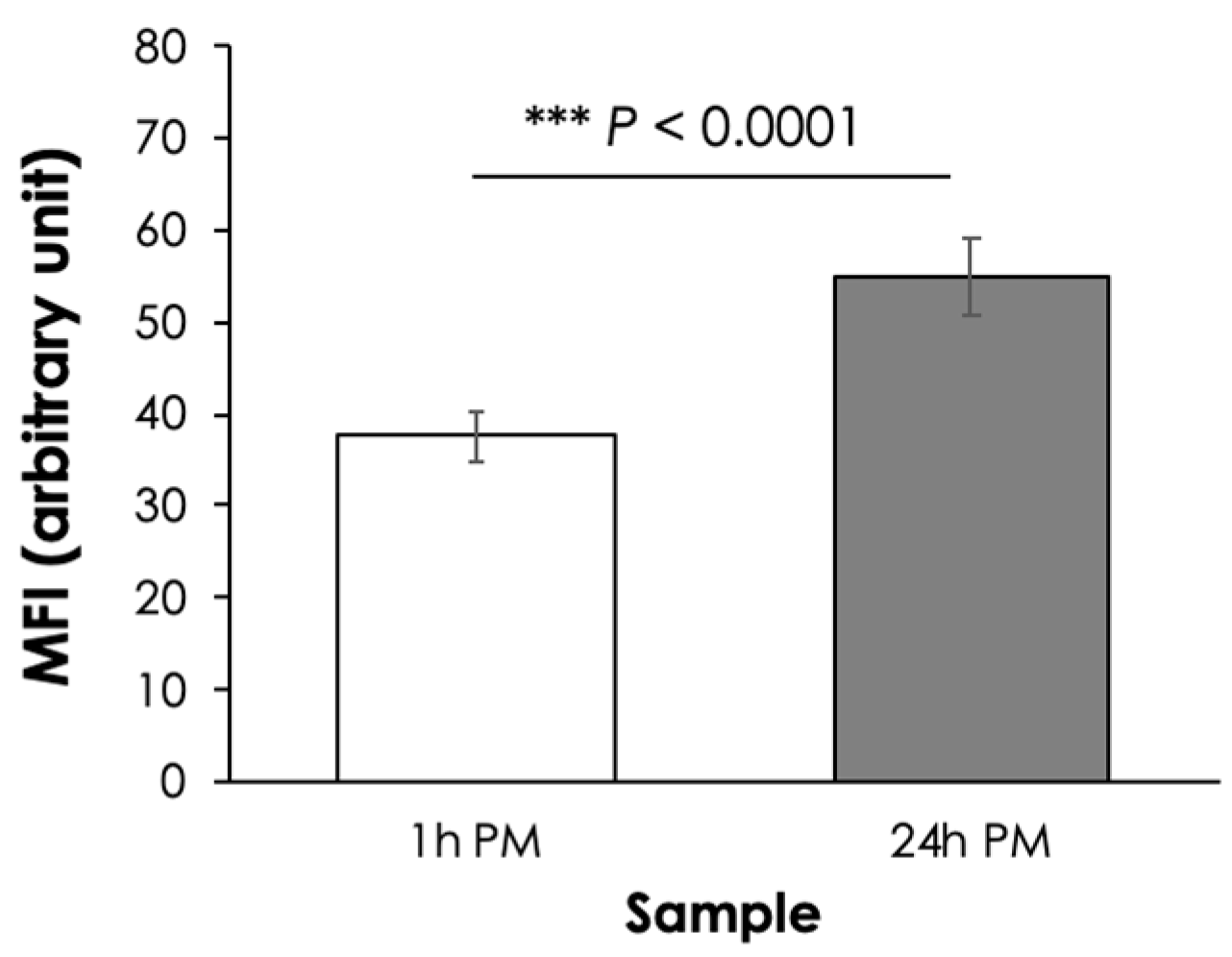
3.1. Myofibril Fragmentation Index (MFI)
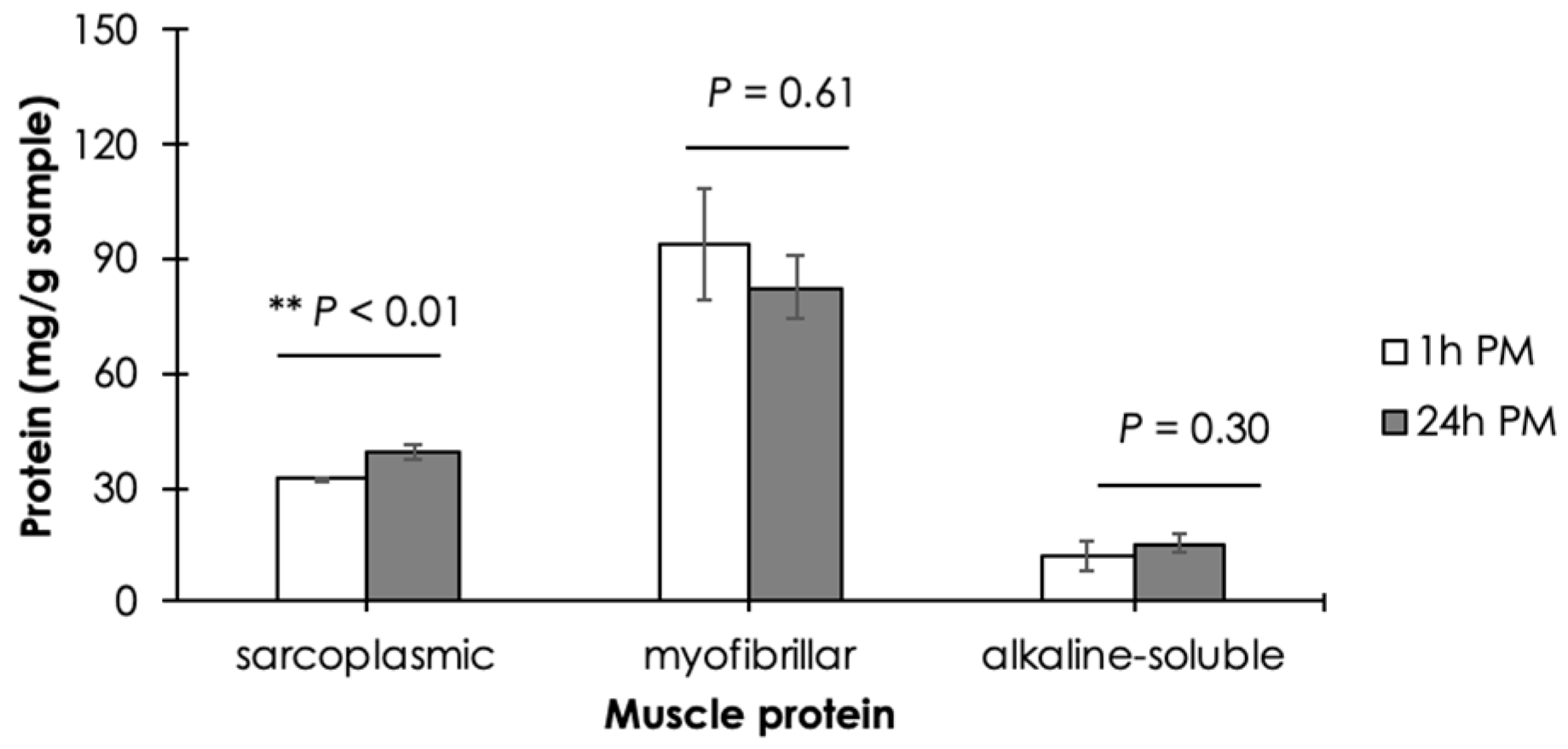
3.2. Muscle Protein Composition
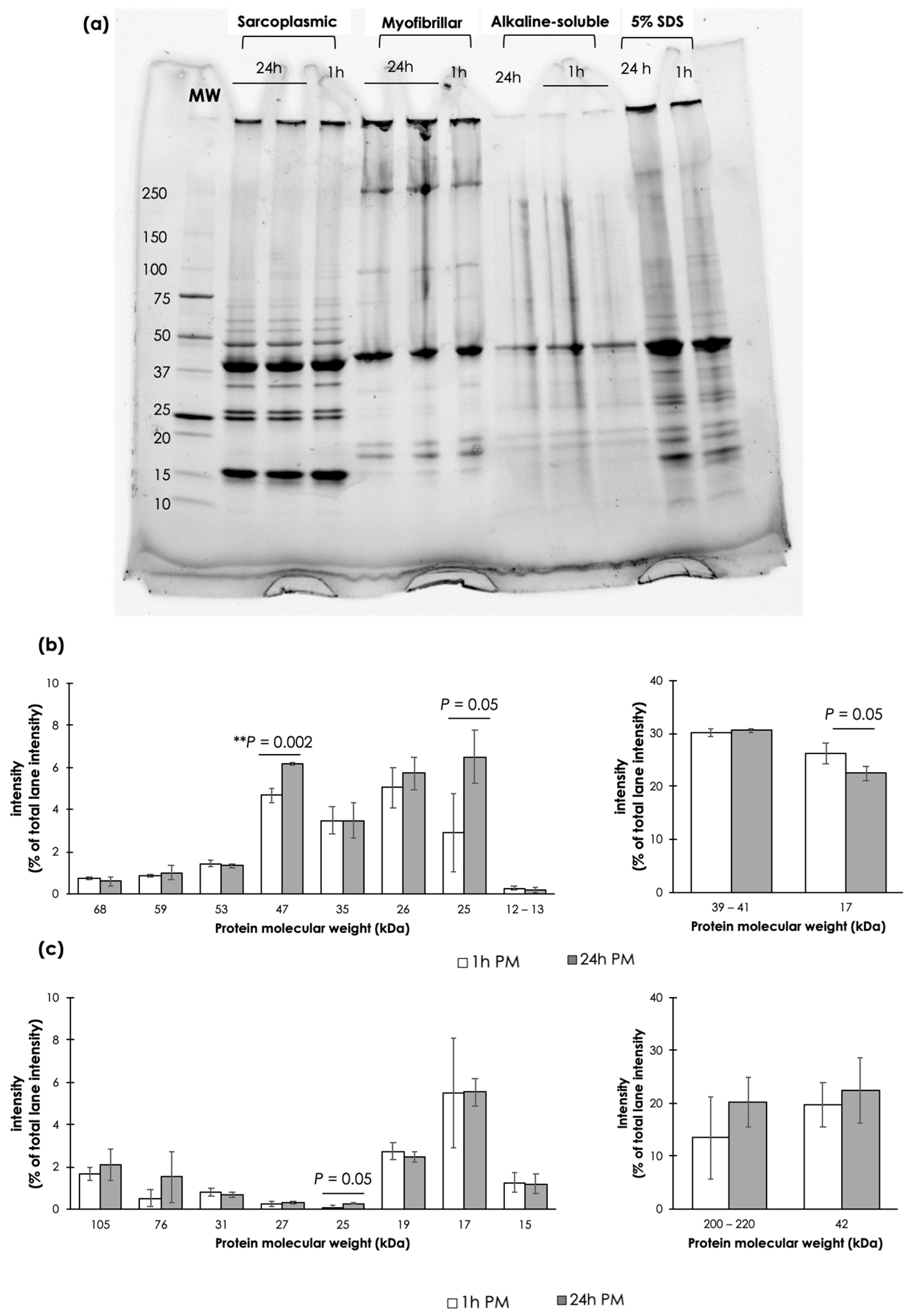
3.3. Protein Electrophoresis
3.4. Technological Properties
3.5. Free Amino Acids in Cooked Sample
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathew, G. Taxonomy, Identification and Biology of Seabass (Lates calcarifer). In Proceedings of the National Training on “Cage Culture of Seabass” Held at CMFRI (Kochi), Kochi, India, 14–23 December 2009. [Google Scholar]
- TAS 7016-2015; Asian Seabass. National Bureau of Agricultural Commodity and Food Standards Ministry of Agriculture and Cooperatives Thai Agricultural Standard: Bangkok, Thailand, 2015; Volume 132, pp. 1–11.
- Panebianco, F.; Nobile, M.; Pasinetti, G.; Pattono, D.; Panseri, S.; Civera, T. Cured or Fresh? Between Fish Maturation Trends in Restaurants and Food Safety: The Case of Dry-Aged Rainbow Trout. Food Control 2024, 165, 110612. [Google Scholar] [CrossRef]
- Verrez-Bagnis, V.; Noel, J.; Sautereau, C.; Fleurence, J. Desmin Degradation in Postmortem Fish Muscle. J. Food Sci. 1999, 64, 240–242. [Google Scholar] [CrossRef]
- Ladrat, C.; Verrez-Bagnis, V.; Noël, J.; Fleurence, J. In Vitro Proteolysis of Myofibrillar and Sarcoplasmic Proteins of White Muscle of Sea Bass (Dicentrarchus labrax L.): Effects of Cathepsins B, D and L. Food Chem. 2003, 81, 517–525. [Google Scholar] [CrossRef]
- Braitenbach Cavali, J.; Vargas Baldi, S.C.; Coutinho Marques Rocha, A.S.; da Silva, E.; Taveira Nunes, C.; Soares, E.C.; de Vargas Schons, S.; Zanella, R.; Bianchini Pontuschka, R.; Vieira Dantas Filho, J. Pre-Slaughter Stunning Methods Influence the Meat Quality of Arapaima Gigas Fillets. Animals 2024, 14, 1155. [Google Scholar] [CrossRef]
- Rucinque, D.S.; Souza, A.P.O.; Molento, C.F.M. Perception of Fish Sentience, Welfare and Humane Slaughter by Highly Educated Citizens of Bogotá, Colombia and Curitiba, Brazil. PLoS ONE 2017, 12, e0168197. [Google Scholar] [CrossRef]
- Ahmed, Z.; Donkor, O.; Street, W.A.; Vasiljevic, T. Calpains- and Cathepsins-Induced Myofibrillar Changes in Post-Mortem Fish: Impact on Structural Softening and Release of Bioactive Peptides. Trends Food Sci. Technol. 2015, 45, 130–146. [Google Scholar] [CrossRef]
- Minami, S.; Takatori, M.; Shirayama, A.; Okita, A.; Nakamura, Y.; Takahashi, K. Taste Components and Texture of Long-Term Aged Fish and Shellfish Sashimi. Nippon. Suisan Gakkaishi 2020, 86, 418–426. [Google Scholar] [CrossRef]
- Poli, B.M.; Parisi, G.; Scappini, F.; Zampacavallo, G. Fish Welfare and Quality as Affected by Pre-Slaughter and Slaughter Management. Aquac. Int. 2005, 13, 29–49. [Google Scholar] [CrossRef]
- Strasburg, G.; Xiong, Y.L. Physiology and Chemistry of Edible Muscle Tissues. In Fennema’s Food Chemistry; CRC Press: Boca Raton, FL, USA, 2017; pp. 955–1015. [Google Scholar]
- England, E.M.; Scheffler, T.L.; Kasten, S.C.; Matarneh, S.K.; Gerrard, D.E. Exploring the Unknowns Involved in the Transformation of Muscle to Meat. Meat Sci. 2013, 95, 837–843. [Google Scholar] [CrossRef]
- Kaur, L.; Hui, S.X.; Morton, J.D.; Kaur, R.; Chian, F.M.; Boland, M. Endogenous Proteolytic Systems and Meat Tenderness: Influence of Post-Mortem Storage and Processing. Food Sci. Anim. Resour. 2021, 41, 589–607. [Google Scholar] [CrossRef]
- Takahashi, K.; Narisawa, Y.; Morii, T.; Morii, S.; Nakamura, Y. Aging of Greater Amberjack Seriola Dumerili Meat Promoted by Low-Temperature Heating. Food Bioproc. Technol. 2023, 16, 2961–2970. [Google Scholar] [CrossRef]
- Terlouw, C.; Bourguet, C. Chapter 1: Quantifying Animal Welfare Preslaughter Using Behavioural, Physiological and Carcass and Meat Quality Measures; Wageningen Academic: Leiden, The Netherlands, 2022; pp. 13–61. ISBN 9789086869244. [Google Scholar]
- Hematyar, N.; Rahimnejad, S.; Gorakh Waghmare, S.; Malinovskyi, O.; Policar, T. Effects of Stocking Density and Pre-Slaughter Handling on the Fillet Quality of Largemouth Bass (Micropterus salmoides): Implications for Fish Welfare. Foods 2024, 13, 1477. [Google Scholar] [CrossRef] [PubMed]
- De Castro, P.L.; Lewandowski, V.; de Souza, M.L.R.; Coradini, M.F.; Alexandre, A.A.D.C.; Sary, C.; Ribeiro, R.P. Effect of Different Periods of Pre-Slaughter Stress on the Quality of the Nile Tilapia Meat. Food Sci. Technol. 2017, 37, 52–58. [Google Scholar] [CrossRef]
- Lippe, G.; Prandi, B.; Bongiorno, T.; Mancuso, F.; Tibaldi, E.; Faccini, A.; Sforza, S.; Stecchini, M.L. The Effect of Pre-Slaughter Starvation on Muscle Protein Degradation in Sea Bream (Sparus aurata): Formation of ACE Inhibitory Peptides and Increased Digestibility of Fillet. Eur. Food Res. Technol. 2021, 247, 259–271. [Google Scholar] [CrossRef]
- Papa, I.; Taylor, R.G.; Astier, C.; Ventre, F.; Lebart, M.C.; Roustan, C.; Ouali, A.; Benyamin, Y. Dystrophin Cleavage and Sarcolemma Detachment Are Early Post Mortem Changes on Bass (Dicentrarchus labrax) White Muscle. J. Food Sci. 1997, 62, 917–921. [Google Scholar] [CrossRef]
- Delbarre-Ladrat, C.; Verrez-Bagnis, V.; Noël, J.; Fleurence, J. Proteolytic Potential in White Muscle of Sea Bass (Dicentrarchus labrax L.) during Post Mortem Storage on Ice: Time-Dependent Changes in the Activity of the Components of the Calpain System. Food Chem. 2004, 84, 441–446. [Google Scholar] [CrossRef]
- Trithavisup, T.; Krobthong, S.; Yingchutrakul, Y.; Sanpinit, P.; Malila, Y. Impact of Wooden Breast Myopathy on in Vitro Protein Digestibility, Metabolomic Profile, and Cell Cytotoxicity of Cooked Chicken Breast Meat. Poult. Sci. 2024, 103, 103261. [Google Scholar] [CrossRef]
- U-chupaj, J.; Malila, Y.; Gamonpilas, C.; Kijroongrojana, K.; Petracci, M.; Benjakul, S.; Visessanguan, W. Differences in Textural Properties of Cooked Caponized and Broiler Chicken Breast Meat. Poult. Sci. 2017, 96, 2491–2500. [Google Scholar] [CrossRef]
- Trithavisup, T.; Sanpinit, P.; Sakulwech, S.; Klamchuen, A.; Malila, Y. In Vitro Protein Digestion of Cooked Spent Commercial Laying Hen and Commercial Broilers Breast Meat. Foods 2022, 11, 1853. [Google Scholar] [CrossRef]
- Caballero, M.J.; Betancor, M.; Escrig, J.C.; Montero, D.; Espinosa de los Monteros, A.; Castro, P.; Ginés, R.; Izquierdo, M. Post Mortem Changes Produced in the Muscle of Sea Bream (Sparus aurata) during Ice Storage. Aquaculture 2009, 291, 210–216. [Google Scholar] [CrossRef]
- Ayala, M.D.; Abdel, I.; Santaella, M.; Martínez, C.; Periago, M.J.; Gil, F.; Blanco, A.; Albors, O.L. Muscle Tissue Structural Changes and Texture Development in Sea Bream, Sparus Aurata L., during Post-Mortem Storage. LWT-Food Sci. Technol. 2010, 43, 465–475. [Google Scholar] [CrossRef]
- Vieira, S.; Franco, A.R.; Fernandes, E.M.; Amorim, S.; Ferreira, H.; Pires, R.A.; Reis, R.L.; Martins, A.; Neves, N.M. Fish Sarcoplasmic Proteins as a High Value Marine Material for Wound Dressing Applications. Colloids Surf. B Biointerfaces 2018, 167, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Mudalal, S.; Babini, E.; Cavani, C.; Petracci, M. Quantity and Functionality of Protein Fractions in Chicken Breast Fillets Affected by White Striping. Poult. Sci. 2014, 93, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.D.; Martínez, T.F.; Sáez, M.I.; Alférez, B.P.; García-Gallego, M. Changes in Muscle Properties during Postmortem Storage of Farmed Sea Bream (Sparus aurata). J. Food Process. Eng. 2011, 34, 922–946. [Google Scholar] [CrossRef]
- Monti, G.; De Napoli, L.; Mainolfi, P.; Barone, R.; Guida, M.; Marino, G.; Amoresano, A. Monitoring Food Quality by Microfluidic Electrophoresis, Gas Chromatography, and Mass Spectrometry Techniques: Effects of Aquaculture on the Sea Bass (Dicentrarchus labrax). Anal. Chem. 2005, 77, 2587–2594. [Google Scholar] [CrossRef]
- Chang-Cheng Zhao, S.B.; Eun, J.-B. Changes in Protein Compositions and Textural Properties of the Muscle of Skate Fermented at 10 °C. Int. J. Food Prop. 2019, 22, 173–185. [Google Scholar] [CrossRef]
- Rotzoll, N.; Dunkel, A.; Hofmann, T. Activity-Guided Identification of (S)-Malic Acid 1-O-d-Glucopyranoside (Morelid) and γ-Aminobutyric Acid as Contributors to Umami Taste and Mouth-Drying Oral Sensation of Morel Mushrooms (Morchella Deliciosa Fr.). J. Agric. Food Chem. 2005, 53, 4149–4156. [Google Scholar] [CrossRef]
- Alam, A.N. Post-Mortem Changes and Assessment of Fish Quality. In Participatory Training of Trainers: A New Approach Applied in Fish Processing; 2007; p. 329, ISBN 984-32-2717-4.
- Poli, B.M.; Parisi, G.; Zampacavallo, G.; Mecatti, M.; Lupi, P.; Gualtieri, M.; Franci, O. Quality Outline of European Sea Bass (Dicentrarchus labrax) Reared in Italy: Shelf Life, Edible Yield, Nutritional and Dietetic Traits. Aquaculture 2001, 202, 303–315. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Littlefield, P.J.; Thompson, J.M. A Research Note on Factors Affecting the Determination of Myofibrillar Fragmentation. Meat Sci. 2000, 56, 19–22. [Google Scholar] [CrossRef]
- Onopiuk, A.; Półtorak, A.; Wierzbicka, A. The Impact of Muscle and Aging Time on Meat Tenderness in the Carcasses of Limousin × Holstein-Friesian Crossbred Bulls. J. Food Process. Preserv. 2018, 42, e13619. [Google Scholar] [CrossRef]
- Li, L.; Zhu, H.; Yi, X.; Nie, Z.; Zheng, Y.; Yang, X.; Xu, P.; Yu, Y.; Xu, G. Study of the Quality and Nutritional Value of Alosa Sapidissima in the Postmortem Process. Fishes 2022, 7, 302. [Google Scholar] [CrossRef]
- Furnesvik, L.; Erkinharju, T.; Hansen, M.; Yousaf, M.N.; Seternes, T. Evaluation of Histological Post-Mortem Changes in Farmed Atlantic Salmon (Salmo salar L.) at Different Time Intervals and Storage Temperatures. J. Fish Dis. 2022, 45, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Verrez-Bagnis, V.; Ladrat, C.; Morzel, M.; Noël, J.; Fleurence, J. Protein Changes in Sea Bass (Dicentrarchus Labrax) Muscle Monitored by One- and Two-Dimensional Gel Electrophoresis. Electrophoresis 2001, 22, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of Taste-Active Amino Acids, Amino Acid Derivatives and Peptides in Food Fermentations—A Review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 1 h PM | 24 h PM | p-Value |
|---|---|---|---|
| Raw sample | |||
| pH | 6.51 ± 0.05 b | 6.63 ± 0.16 a | 0.011 |
| Surface color | |||
| L*-value | 39.31 ± 2.44 b | 43.82 ± 4.06 a | <0.001 |
| a*-value | 1.57 ± 0.85 | 2.00 ± 1.19 | 0.088 |
| b*-value | −3.38 ± 1.15 b | −2.01 ± 1.33 a | 0.001 |
| Thawing loss (%) | 1.16 ± 0.21 | 1.62 ± 0.77 | 0.238 |
| Cooked sample | |||
| pH | 6.81 ± 0.05 | 6.77 ± 0.11 | 0.242 |
| Surface color | |||
| L*-value | 75.55 ± 2.42 | 74.24 ± 2.47 | 0.061 |
| a*-value | −0.40 ± 1.21 b | 0.56 ± 0.86 a | 0.047 |
| b*-value | 7.62 ± 1.36 | 7.09 ± 1.21 | 0.153 |
| Cooking loss (%) | 15.88 ± 5.48 | 10.56 ± 4.67 | 0.149 |
| Expressible water (%) | 13.83 ± 2.12 a | 9.93 ± 2.65 b | 0.004 |
| Texture | |||
| Hardness (g) | 132.60 ± 30.90 a | 104.70 ± 35.56 b | 0.047 |
| Springiness | 0.52 ± 0.05 | 0.51 ± 0.12 | 0.526 |
| Cohesiveness | 0.30 ± 0.03 | 0.27 ± 0.08 | 0.145 |
| Chewiness (g) | 20.85 ± 5.77 | 15.74 ± 9.30 | 0.065 |
| Amino Acid | MW 2 | Taste Threshold 3 | 1 h PM | 24 h PM | p-Value | |
|---|---|---|---|---|---|---|
| (g/mol) | (mg/100 g) | |||||
| Sweet-tasting amino acids | ||||||
| L-alanine | Ala | 89.00 | 106.80 | 15.92 ± 0.38 b | 16.86 ± 0.29 a | 0.026 |
| L-glycine | Gly | 75.07 | 187.68 | 65.55 ± 2.34 b | 73.69 ± 0.41 a | 0.004 |
| L-proline | Pro | 115.13 | 172.70 | 40.44 ± 1.48 a | 15.82 ± 0.27 b | <0.0001 |
| L-serine | Ser | 105.09 | 262.73 | 12.73 ± 0.78 a | 8.81 ± 0.04 b | 0.001 |
| L-threonine | Thr | 119.12 | 416.92 | 7.56 ± 0.47 | 7.84 ± 0.09 | 0.369 |
| Umami-tasting amino acids | ||||||
| L-aspartic acid | Asp | 133.10 | 53.24 | 1.42 ± 0.06 b | 3.28 ± 0.05 a | <0.0001 |
| L-glutamic acid | Glu | 147.13 | 16.18 | 19.37 ± 1.00 b | 23.40 ± 0.33 a | 0.003 |
| Bitter-tasting amino acids | ||||||
| L-leucine | Leu | 131.17 | 144.29 | 3.96 ± 0.14 b | 8.89 ± 0.10 a | <0.0001 |
| L-isoleucine | Ile | 131.17 | 131.17 | 3.38 ± 0.05 b | 5.71 ± 0.14 a | <0.0001 |
| L-arginine | Arg | 174.20 | 1306.50 | 17.91 ± 1.25 b | 23.98 ± 1.36 a | 0.005 |
| L-histidine | His | 155.15 | 698.18 | 11.96 ± 0.86 a | 7.04 ± 0.30 b | 0.001 |
| L-valine | Val | 117.15 | 351.45 | 2.97 ± 0.12 b | 5.63 ± 0.05 a | <0.0001 |
| L-phenylalanine | Phe | 165.19 | 743.36 | 1.74 ± 0.07 b | 3.97 ± 0.06 a | <0.0001 |
| L-lysine | Lys | 146.19 | 1169.52 | 3.79 ± 0.23 b | 13.28 ± 0.04 a | <0.0001 |
| L-tyrosine | Tyr | 181.19 | 72.48 | 1.05 ± 0.09 b | 2.95 ± 0.08 a | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malila, Y.; Charoensuk, D.; Srimarut, Y.; Saensa-ard, S.; Petpiroon, N.; Kunyanee, C.; Rattanawongsa, W.; Saenmuangchin, R.; Klamchuen, A.; Kangwansupamonkon, W.; et al. Muscle Proteins, Technological Properties, and Free Amino Acids of Epaxial Muscle Collected from Asian Seabass (Lates calcarifer) at Different Postmortem Durations. Animals 2024, 14, 2837. https://doi.org/10.3390/ani14192837
Malila Y, Charoensuk D, Srimarut Y, Saensa-ard S, Petpiroon N, Kunyanee C, Rattanawongsa W, Saenmuangchin R, Klamchuen A, Kangwansupamonkon W, et al. Muscle Proteins, Technological Properties, and Free Amino Acids of Epaxial Muscle Collected from Asian Seabass (Lates calcarifer) at Different Postmortem Durations. Animals. 2024; 14(19):2837. https://doi.org/10.3390/ani14192837
Chicago/Turabian StyleMalila, Yuwares, Danai Charoensuk, Yanee Srimarut, Sunitta Saensa-ard, Nalinrat Petpiroon, Chanikarn Kunyanee, Wachiraya Rattanawongsa, Rattaporn Saenmuangchin, Annop Klamchuen, Wiyong Kangwansupamonkon, and et al. 2024. "Muscle Proteins, Technological Properties, and Free Amino Acids of Epaxial Muscle Collected from Asian Seabass (Lates calcarifer) at Different Postmortem Durations" Animals 14, no. 19: 2837. https://doi.org/10.3390/ani14192837