

Environmental Drivers of Local Demography and Size Plasticity in Fire Salamanders (Salamandra salamandra)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Skeletochronological Age Estimation
2.4. Potential Drivers of Local Mortality and Size Variation
2.5. Statistical Analyses
3. Results
3.1. Size at Metamorphosis and Water Quality
3.2. Size and Age Variation in Adults
3.3. Environmental Drivers of Local Demography
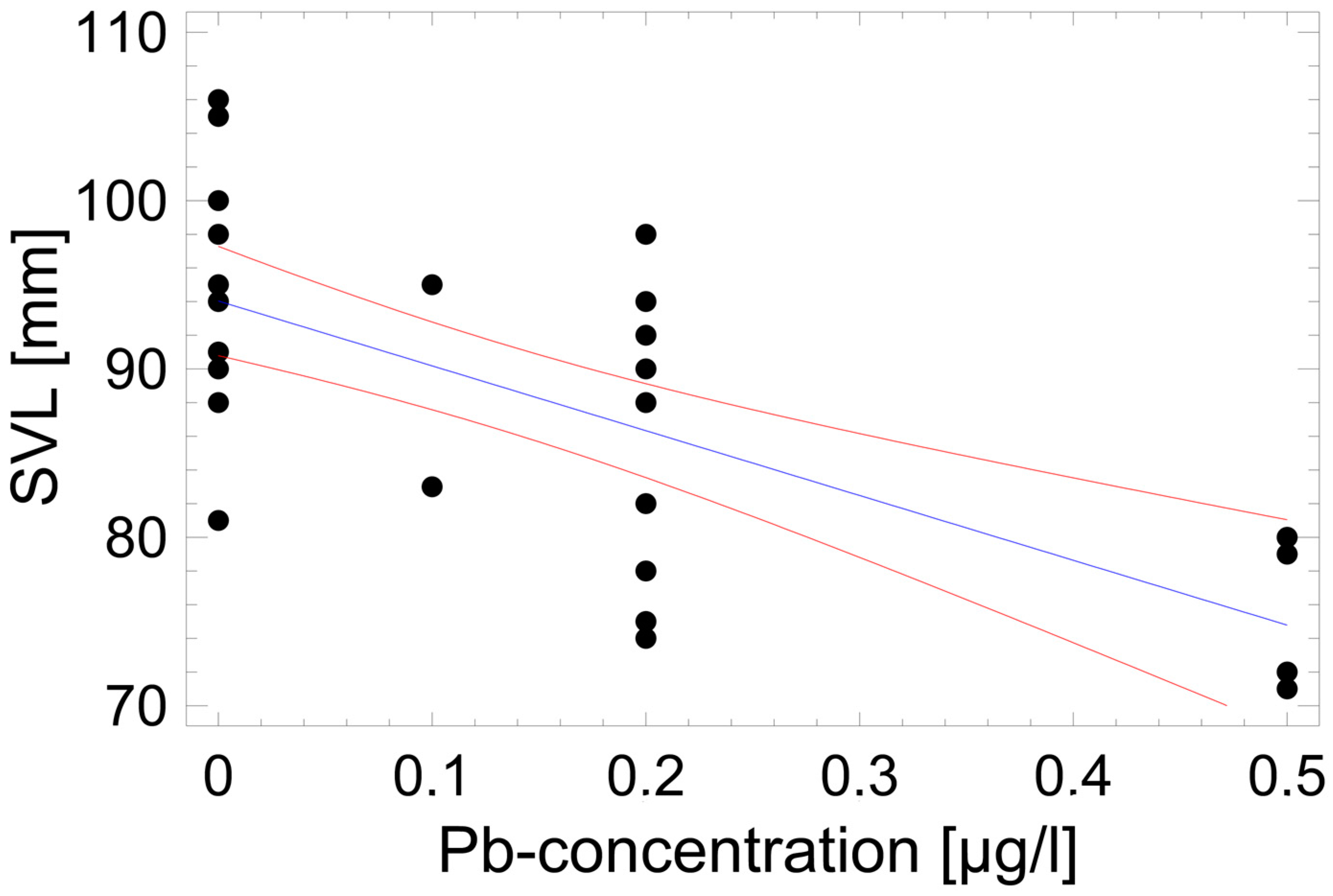
3.4. Environmental Drivers of Local SVL Distribution
4. Discussion
4.1. Demography
4.2. Size Plasticity
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crispo, E. The Baldwin effect and genetic assimilation: Revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity. Evolution 2007, 61, 2469–2479. [Google Scholar] [CrossRef] [PubMed]
- Pfennig, D.W. (Ed.) Phenotypic Plasticity & Evolution: Causes, Consequences, Controversies, 1st ed.; CRC Press & Taylor & Francis: Boca Raton, FL, USA, 2021; p. 437. [Google Scholar] [CrossRef]
- Reed, T.E.; Waples, R.S.; Schindler, D.E.; Hard, J.J.; Kinnison, M.T. Phenotypic plasticity and population viability: The importance of environmental predictability. Proc. R. Soc. B Biol. Sci. 2010, 277, 3391–3400. [Google Scholar] [CrossRef] [PubMed]
- Bylino, O.V.; Ogienko, A.A.; Batin, M.A.; Georgiev, P.G.; Omelina, E.S. Genetic, Environmental, and Stochastic Components of Lifespan Variability: The Drosophila Paradigm. Int. J. Mol. Sci. 2024, 25, 4482. [Google Scholar] [CrossRef]
- Flatt, T.; Amdam, G.V.; Kirkwood, T.B.; Omholt, S.W. Life-history evolution and the polyphenic regulation of somatic maintenance and survival. Q Rev. Biol. 2013, 88, 185–218. [Google Scholar] [CrossRef] [PubMed]
- Miaud, C.; Guyetant, R.; Elmberg, J. Variations in life-history traits in the common frog Rana temporaria (Amphibia: Anura): A literature review and new data from the French Alps. J. Zool. 1999, 249, 61–73. [Google Scholar] [CrossRef]
- Moran, N.A. Adaptation and Constraint in the Complex Life Cycles of Animals. Annu. Rev. Ecol. Syst. 1994, 25, 573–600. [Google Scholar] [CrossRef]
- Denver, R.J.; Middlemis-Maher, J. Lessons from evolution: Developmental plasticity in vertebrates with complex life cycles. J. Dev. Orig. Health Dis. 2010, 1, 282–291. [Google Scholar] [CrossRef]
- Moore, M.P.; Martin, R.A. On the evolution of carry-over effects. J. Anim. Ecol. 2019, 88, 1832–1844. [Google Scholar] [CrossRef]
- Freidenburg, L.K. Environmental drivers of carry-over effects in a pond-breeding amphibian, the Wood Frog (Rana sylvatica). Can. J. Zool. 2017, 95, 255–262. [Google Scholar] [CrossRef]
- Sinsch, U.; Leus, F.; Sonntag, M.; Hantzschmann, A.M. Carry-over effects of the larval environment on the post-metamorphic performance of Bombina variegata (Amphibia, Anura). Herpetol. J. 2020, 30, 126–134. [Google Scholar] [CrossRef]
- Burraco, P.; Valdés, A.E.; Orizaola, G. Metabolic costs of altered growth trajectories across life transitions in amphibians. J. Anim. Ecol. 2020, 89, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Charbonnier, J.F.; Pearlmutter, J.; Vonesh, J.R.; Gabor, C.R.; Forsburg, Z.R.; Grayson, K.L. Cross-Life Stage Effects of Aquatic Larval Density and Terrestrial Moisture on Growth and Corticosterone in the Spotted Salamander. Diversity 2018, 10, 68. [Google Scholar] [CrossRef]
- Klewen, R. Untersuchungen zur Ökologie und Populationsbiologie des Feuersalamanders (Salamandra salamandra terrestris Lacepede 1788) an einer isolierten Population im Kreise Paderborn. Abh. Aus Dem Mus. Für Naturkunde 1985, 47, 1–51. [Google Scholar]
- Schulte, U.; Küsters, D.; Steinfartz, S. A PIT tag-based analysis of annual movement patterns of adult fire salamanders (Salamandra salamandra) in a Middle European habitat. Amphib.-Reptil. 2007, 28, 531–536. [Google Scholar] [CrossRef]
- Bani, L.; Pisa, G.; Luppi, M.; Spilotros, G.; Fabbri, E.; Randi, E.; Orioli, V. Ecological connectivity assessment in a strongly structured fire salamander (Salamandra salamandra) population. Ecol. Evol. 2015, 5, 3472–3485. [Google Scholar] [CrossRef]
- Najbar, A.; Konowalik, A.; Halupka, K.; Najbar, B.; Ogielska, M. Body size and life history traits of the fire salamander Salamandra salamandra from Poland. Amphib. Reptil. 2020, 41, 63–74. [Google Scholar] [CrossRef]
- Kalezic, M.L.; Dzukic, G.; Djorovic, A.; Aleksic, I. Body size, age and sexual size dimorphism in the genus Salamandra. A study of Balkan species. Spixiana 2000, 23, 283–293. [Google Scholar]
- Rebelo, R.; Caetano, M.H. Body size differences among populations of Salamandra salamandra from Portugal: A skeletochronological study. In Proceedings of the 8th Ordinary General Meeting of the Societas Europea Herpetologica, Bonn, Germany, 23–27 August 1995; pp. 307–312. [Google Scholar]
- Alcobendas, M.; Castanet, J. Bone growth plasticity among populations of Salamandra salamandra: Interactions between internal and external factors. Herpetologica 2000, 56, 14–26. [Google Scholar]
- Reinhard, S.; Renner, S.; Kupfer, A. Age and fecundity in Salamandra algira (Caudata: Salamandridae). Salamandra 2015, 51, 19–24. [Google Scholar]
- Reinhard, S.; Renner, S.; Kupfer, A. Sexual dimorphism and age of mediterranean salamanders. Zoology 2015, 118, 19–26. [Google Scholar] [CrossRef]
- Bouzid, S.; Konecny, L.; Grolet, O.; Douady, C.J.; Joly, P.; Bouslama, Z. Phylogeny, age structure, growth dynamics and colour pattern of the Salamandra algira algira population in the Edough Massif, northeastern Algeria. Amphib. Reptil. 2017, 38, 461–471. [Google Scholar] [CrossRef]
- Altunişik, A. Age, Survivorship and Life Expectancy in Near Eastern Fire Salamander, Salamandra infraimmaculata (Caudata: Salamandridae). Russ. J. Ecol. 2018, 49, 166–171. [Google Scholar] [CrossRef]
- Miaud, C.; Andreone, F.; Riberon, A.; De Michelis, S.; Clima, V.; Castanet, J.; Francillon-Vieillot, H.; Guyetant, R. Variations in age, size at maturity and gestation duration among two neighbouring populations of the alpine salamander (Salamandra lanzai). J. Zool. 2001, 254, 251–260. [Google Scholar] [CrossRef]
- Andreone, F.; Miaud, C.; Bergo, P.; Doglio, S.; Stocco, P.; Riberon, A.; Gautier, P. Living at high altitude: Testing the effects of life history traits upon the conservation of Salamandra lanzai (Amphibia, Salamandridae). Ital. J. Zool. 2004, 71, 35–43. [Google Scholar] [CrossRef]
- Fachbach, G. Röhrenknochenentwicklung und Altersbestimmung bei Salamandra atra Laur. (Urodela, Salamandridae). Zool. Anz. 1988, 221, 188–200. [Google Scholar]
- Böhme, W. Zum Höchstalter des Feuersalamanders (Salamandra salamandra (Linnaeus, 1758): Ein wiederentdecktes Dokument aus der Frühzeit der Terraristik (Amphibia: Caudata: Salamandridae). Salamandra 1979, 15, 176–179. [Google Scholar]
- Lüddecke, T.; Schulz, S.; Steinfartz, S.; Vences, M. A salamander’s toxic arsenal: Review of skin poison diversity and function in true salamanders, genus. Salamandra Sci. Nat. 2018, 105, 56. [Google Scholar] [CrossRef]
- Caspers, B.A.; Krause, E.T.; Hermanski, I.; Wiesbrock, C.; Kastrup, F.-W.; Steinfartz, S. Developmental costs of yellow colouration in fire salamanders and experiments to test the efficiency of yellow as a warning colouration. Amphib. Reptil. 2020, 41, 373–385. [Google Scholar] [CrossRef]
- De Meester, G.; Šunje, E.; Prinsen, E.; Verbruggen, E.; Van Damme, R. Toxin variation among salamander populations: Discussing potential causes and future directions. Integr. Zool. 2021, 16, 336–353. [Google Scholar] [CrossRef]
- Kabisch, K.; Belter, H. Das Verzehren von Amphibien und Reptilien durch Vögel. Zool. Abh. Aus Dem Staatl. Mus. Für Tierkd. Dresd. 1968, 29, 191–227. [Google Scholar]
- Sauer, H.; Weisbecker, H. Einheimische Schlangen als gelegentliche Verfolger des Feuersalamanders (Salamandra salamandra)—Zwei Feldbeobachtungen. Nat. Mus. 1994, 124, 349–350. [Google Scholar]
- Pezaro, N.; Rovelli, V.; Segev, O.; Templeton, A.R.; Blaustein, L. Suspected rat predation on the Near Eastern Fire Salamander (Salamandra infraimmaculata) by selective consumption of non-toxic tissue. Zool. Middle East 2018, 64, 91–93. [Google Scholar] [CrossRef]
- Koynova, T.; Tsvetkov, R.; Natchev, N. Do it like the otter—Data on the amphibian skinning behaviour in the Eurasian badger (Meles meles L., 1758). Acta Ethologica 2023, 26, 225–229. [Google Scholar] [CrossRef]
- Velo-Antón, G. When aposematism is not enough: Exotic Rattus rattus shows no mercy for carcasses of Salamandra salamandra in insular populations. Ecol. Evol. 2024, 14, e11229. [Google Scholar] [CrossRef] [PubMed]
- Thiesmeier, B.; Grossenbacher, K. Salamandra salamandra (Linnaeus, 1758)—Feuersalamander. In Handbuch der Reptilien und Amphibien Europas; Thiesmeier, B., Grossenbacher, K., Eds.; Aula: Wiebelsheim, Germany, 2004; Volume 4/IIB, pp. 1059–1132. [Google Scholar]
- Thiesmeier, B. Der Feuersalamander. Z. Feldherpetologie 2004, 4, 192. [Google Scholar]
- Manenti, R.; Ficetola, G.F.; De Bernardi, F. Water, stream morphology and landscape: Complex habitat determinants for the fire salamander Salamandra salamandra. Amphib. Reptil. 2009, 30, 7–15. [Google Scholar] [CrossRef]
- Börder, M.; Karlsson, A.; Sinsch, U. Bestandsdichte, Arealnutzung und Gefährdung einer Feuersalamander-Population (Salamandra salamandra) im Stadtgebiet von Koblenz (Rheinland-Pfalz). Z. Feldherpetologie 2011, 18, 99–116. [Google Scholar]
- Altmayer, D.B.; Eder, D.J.; Eichhorn, D.J.; Fader, B.; Hetterling, U.; Huth, D.C.; Kameke, D.; Kortekamp, D.A.; Ipach, R.; Ipach, D.U.; et al. Lehrgangsbegleitheft Sachkunde im Pflanzenschutz Weinbau, 12th ed.; DLR (Dienstleistungszentrum Ländlicher Raum Rheinpfalz): Neustadt an der Weinstraße, Germany, 2023; p. 176. [Google Scholar]
- Bereswill, R.; Golla, B.; Streloke, M.; Schulz, R. Entry and toxicity of organic pesticides and copper in vineyard streams: Erosion rills jeopardise the efficiency of riparian buffer strips. Agric. Ecosyst. Environ. 2012, 146, 81–92. [Google Scholar] [CrossRef]
- Herwig, N.; Hommel, B.; Felgentreu, D. Copper distribution in German vineyards and its impact on soil organisms—Dataset of a field survey from 2010 to 2014. Data Brief 2022, 48, 109088. [Google Scholar] [CrossRef]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J. 2015, 25, 5–13. [Google Scholar]
- Sinsch, U.; Oromi, N.; Sanuy, D. Growth marks in natterjack toad (Bufo calamita) bones: Histological correlates of hibernation and aestivation periods. Herpetol. J. 2007, 17, 129–137. [Google Scholar]
- Burgstaller, S.; Leeb, C.; Ringler, M.; Gollmann, G. Demography and spatial activity of fire salamanders, Salamandra salamandra (Linnaeus,1758), in two contrasting habitats in the Vienna Woods. Herpetozoa 2021, 34, 23–34. [Google Scholar] [CrossRef]
- DWD Climate Data Center (CDC). Vieljährige Stationsmittelwerte für die Klimareferenzperiode 1981–2010. 2024. Available online: https://cdc.dwd.de (accessed on 19 September 2024).
- Pröfrock, D.; Prange, A. Inductively coupled plasma–mass spectrometry (ICP-MS) for quantitative analysis in environmental and life sciences: A review of challenges, solutions, and trends. Appl. Spectrosc. 2012, 66, 843–868. [Google Scholar] [CrossRef] [PubMed]
- Chardon, J.P.; Adriaensen, F.; Matthysen, E. Incorporating landscape elements into a connectivity measure: A case study for the Speckled wood butterfly (Pararge aegeria L.). Landsc. Ecol. 2003, 18, 561–573. [Google Scholar] [CrossRef]
- von Bertalanffy, L. A quantitative theory of organic growth (Inquires on growth laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Brizzi, R.; Calloni, C. Male cloacal region of the spotted salamander, Salamandra salamandra gigliolii (Amphibia, Salamandridae). Ital. J. Zool. 1992, 59, 377–385. [Google Scholar] [CrossRef]
- Labus, N.; Cvijanović, M.; Vukov, T. Sexual size and shape dimorphism in Salamandra salamandra (Amphibia, Caudata, Salamandridae) from the central Balkans. Arch. Biol. Sci. 2013, 65, 969–975. [Google Scholar] [CrossRef]
- Lunghi, E.; Corti, C.; Biaggini, M.; Zhao, Y.; Cianferoni, F. The Trophic Niche of Two Sympatric Species of Salamanders (Plethodontidae and Salamandridae) from Italy. Animals 2022, 12, 2221. [Google Scholar] [CrossRef]
- Hantzschmann, A.M.; Gollmann, B.; Gollmann, G.; Sinsch, U. The fast–slow continuum of longevity among yellow-bellied toad populations (Bombina variegata): Intrinsic and extrinsic drivers of variation. PeerJ 2019, 7, e8233. [Google Scholar] [CrossRef]
- Cayuela, H.; Griffiths, R.A.; Zakaria, N.; Arntzen, J.W.; Priol, P.; Lena, J.P.; Besnard, A.; Joly, P. Drivers of amphibian population dynamics and asynchrony at local and regional scales. J. Anim. Ecol. 2020, 89, 1350–1364. [Google Scholar] [CrossRef]
- Blanco, M.A.; Sherman, P.W. Maximum longevities of chemically protected and non-protected fishes, reptiles, and amphibians support evolutionary hypotheses of aging. Mech. Ageing Dev. 2005, 126, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Mebs, D.; Kwet, A.; Schneider, M. Tetrodotoxin and its analogue 6-epitetrodotoxin in newts (Triturus spp.; Urodela, Salamandridae) from southern Germany. Toxicon 2007, 50, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Łaciak, M.; Adamski, P.; Bielański, W.; Ćmiel, A.; Lipińska, A.; Łaciak, T.; Zając, T. Tastier than thought? Telemetric research reveals predation on the poisonous yellow-bellied toad Bombina variegata. Ecology 2023, 104, e3996. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.R.; Feldmann, R.; Schaub, M. Demographic processes underlying population growth and decline in Salamandra salamandra. Conserv. Biol. 2005, 19, 1149–1156. [Google Scholar] [CrossRef]
- Angelini, C.; Tiberi, A.; Cari, B.; Giachi, F. Survival and recruitment in the population ecology of the endangered Bombina pachypus (Amphibia: Anura). Amphib. Reptil. 2018, 39, 265–274. [Google Scholar] [CrossRef]
- Homan, R.N.; Holgerson, M.A.; Biga, L.M. A Long-term Demographic Study of a Spotted Salamander (Ambystoma maculatum) Population in Central Ohio. Herpetologica 2018, 74, 109–116. [Google Scholar] [CrossRef]
- Adlassnig, W.; Schmidt, B.; Jirsa, F.; Gradwohl, A.; Ivesic, C.; Koller-Peroutka, M. The Arsenic–Antimony Creek at Sauerbrunn/Burgenland, Austria: A Toxic Habitat for Amphibians. Int. J. Environ. Res. Public Health 2022, 19, 6010. [Google Scholar] [CrossRef]
- Bodinof-Jachowski, C.M.; Hopkins, W.A. Loss of catchment-wide riparian forest cover is associated with reduced recruitment in a long-lived amphibian. Biol. Conserv. 2018, 220, 215–227. [Google Scholar] [CrossRef]
- Adlassnig, W.; Sassmann, S.; Grawunder, A.; Puschenreiter, M.; Horvath, A.; Koller-Peroutka, M. Amphibians in metal-contaminated habitats. Salamandra 2013, 49, 149–158. [Google Scholar]
- Brodie, E.D., Jr. Salamander Antipredator Postures. Copeia 1977, 1977, 523–535. [Google Scholar] [CrossRef]
- Krause, E.T.; Caspers, B.A. Long-term consequences of early nutritional conditions on the behaviour and growth of fire salamanders. Amphib.-Reptil. 2016, 37, 69–77. [Google Scholar] [CrossRef]
- Manenti, R.; Kristensen, N.; Cogliati, P.; Barzaghi, B.; Melotto, A.; Ficetola, G.F. Larval development and poor trophic resource availability: Local adaptations and plasticity in a widespread amphibian species. J. Evol. Biol. 2023, 36, 529–541. [Google Scholar] [CrossRef]
- Băncilă, R.I.; Stănescu, F.; Plăiaşu, R.; Nae, I.; Székely, D.; Vlad, S.E.; Cogălniceanu, D. Food and light availability induce plastic responses in fire salamander larvae from contrasting environments. PeerJ 2023, 11, e16046. [Google Scholar] [CrossRef]
- Nebeker, A.; Schuytema, G.; Ott, S. Effects of cadmium on growth and bioaccumulation in the Northwestern salamander Ambystoma gracile. Arch. Environ. Contam. Toxicol. 1995, 29, 492–499. [Google Scholar] [CrossRef]
- Lefcort, H.; Meguire, R.A.; Wilson, L.H.; Ettinger, W.F. Heavy Metals Alter the Survival, Growth, Metamorphosis, and Antipredatory Behavior of Columbia Spotted Frog (Rana luteiventris) Tadpoles. Arch. Environ. Contam. Toxicol. 1998, 35, 447–456. [Google Scholar] [CrossRef]
- Gardner, S.; Cline, G.; Mwebi, N.; Rayburn, J. Natural tissue concentrations in adult Ambystoma maculatum and larval DNA damage from exposure to arsenic and chromium. J. Toxicol. Environ. Health Part A Curr. Issues 2018, 81, 512–524. [Google Scholar] [CrossRef]
- Arrieta, M.A.; Apartin, C.; Rosenberg, C.E.; Fink, N.E.; Salibian, A. Blood lead content in a peri-urban population of the South American toad Bufo arenarum. Sci. Total Environ. 2001, 271, 99–105. [Google Scholar] [CrossRef]
- Lenhardt, P.P.; Brühl, C.A.; Leeb, C.; Theissinger, K. Amphibian population genetics in agricultural landscapes: Does viniculture drive the population structuring of the European common frog (Rana temporaria)? PeerJ 2017, 5, e3520. [Google Scholar] [CrossRef]
- Leeb, C.; Brühl, C.; Theissinger, K. Potential pesticide exposure during the post-breeding migration of the common toad (Bufo bufo) in a vineyard dominated landscape. Sci. Total Environ. 2020, 706, 134430. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, R.; Ai, S.; Yang, Y.; Ding, J.; Zhang, Y. Long-term heavy metal pollution varied female reproduction investment in free-living anura, Bufo raddei. Ecotoxicol. Environ. Saf. 2018, 159, 136–142. [Google Scholar] [CrossRef]
- Kiss, I.; Vörös, J.; Hamer, A.J. Movement patterns within an urban population of fire salamanders highlight the importance of conserving small habitat patches. J. Zool. 2022, 316, 240–249. [Google Scholar] [CrossRef]
- Bruno, S. Anfibi d’Italia: Caudata. Natura 1973, 64, 209–450. [Google Scholar]
- Catenazzi, A. Ecologie d’une Population de Salamandre Tachetée au sud Des Alpes. Master´s Thesis, Université de Neuchâtel, Neuchâtel, Switzerland, 1998. [Google Scholar]
- Manenti, R.; Conti, A.; Pennati, R. Fire salamander (Salamandra salamandra) males’ activity during breeding season: Effects of microhabitat features and body size. Acta Herpetol. 2017, 12, 29–36. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinsch, U. Environmental Drivers of Local Demography and Size Plasticity in Fire Salamanders (Salamandra salamandra). Animals 2024, 14, 2869. https://doi.org/10.3390/ani14192869
Sinsch U. Environmental Drivers of Local Demography and Size Plasticity in Fire Salamanders (Salamandra salamandra). Animals. 2024; 14(19):2869. https://doi.org/10.3390/ani14192869
Chicago/Turabian StyleSinsch, Ulrich. 2024. "Environmental Drivers of Local Demography and Size Plasticity in Fire Salamanders (Salamandra salamandra)" Animals 14, no. 19: 2869. https://doi.org/10.3390/ani14192869