
Spotting the Pattern: A Review on White Coat Color in the Domestic Horse
by
 , , and
, , and
Aiden McFadden
1,*,
Micaela Vierra
1,
Katie Martin
1,
Samantha A. Brooks
2 ,
,
Robin E. Everts
1 and
and
Christa Lafayette
1 
1
Etalon Inc., Menlo Park, CA 94025, USA
2
Department of Animal Sciences, UF Genetics Institute, University of Florida, Gainesville, FL 32611, USA
*
Author to whom correspondence should be addressed.
Animals 2024, 14(3), 451; https://doi.org/10.3390/ani14030451
Submission received: 25 December 2023
/
Revised: 25 January 2024
/
Accepted: 27 January 2024
/
Published: 30 January 2024
(This article belongs to the Special Issue Genetics of Coat Color in Animals)
Abstract
:Simple Summary
The understanding of coat color genetics in the domestic horse has advanced largely in the last few years. Specifically, many alleles influencing equine white spotting have been described in the last few decades. White spotting phenotypes can range from small white facial spots to an entirely white horse. Although white markings may be the most obvious phenotype, many white-causing alleles also cause adverse health defects, including sterility, deafness, and blindness. Many white spotting alleles are considered embryonic lethal in the homozygous state, causing obstacles in the breeding process. This review aims to concisely summarize recent research related to the genetics of equine white spotting phenotypes.
Abstract
Traits such as shape, size, and color often influence the economic and sentimental value of a horse. Around the world, horses are bred and prized for the colors and markings that make their unique coat patterns stand out from the crowd. The underlying genetic mechanisms determining the color of a horse’s coat can vary greatly in their complexity. For example, only two genetic markers are used to determine a horse’s base coat color, whereas over 50 genetic variations have been discovered to cause white patterning in horses. Some of these white-causing mutations are benign and beautiful, while others have a notable impact on horse health. Negative effects range from slightly more innocuous defects, like deafness, to more pernicious defects, such as the lethal developmental defect incurred when a horse inherits two copies of the Lethal White Overo allele. In this review, we explore, in detail, the etiology of white spotting and its overall effect on the domestic horse to Spot the Pattern of these beautiful (and sometimes dangerous) white mutations.
1. Introduction
A streak of white, a patch of pink skin, a piercing blue eye. These traits add individuality, stunning beauty, and economic value to the domestic horse (Equus caballus). Specific color traits can help a horse qualify for color-specific registries. However, some of these prized alleles can cause detrimental phenotypes, such as an increased risk of deafness or blindness. Other combinations of these sought-after color traits fail to produce viable offspring. Understanding the etiology of white markings is crucial to ensure ethical care and breeding practices.
Recent advancements have decreased the cost of genome sequencing, enabling the elucidation of the characteristics of the genetic mutations causing many depigmentation phenotypes. KIT, MITF, PAX3, HPS5, EDNRB, TRPM1, and RFWD3 represent the collection of genes associated with different white markings in the domestic horse (Table 1). While the genetic mechanisms underlying some of these color traits are not fully understood, the information collected on white spotting mutations can assist breeders in making optimal choices to breed unique and healthy herds. In this review, we explore the mechanisms and phenotypic effects of equine depigmentation and their overall effect on horse health.
2. Dominant White
A “white” horse has held a special place throughout history and mythology, appearing in many great legends and tales. However, the horses captured in the literature and art do not always depict a truly white horse, whose phenotypes are often confused with Grey or Cream. Because horses homozygous for Cream have pink skin and hair diluted to a near-white color, it is difficult to differentiate between true white and homozygous Cream horses [1]. Dominant White horses are born with white markings and display pink skin below these areas, while individuals with Grey do not have pink skin and are not born with white hairs, but develop them with age [1]. The Romans knew of the phenotypic differences between gray and white, although it remains unknown if their terms for these colors correspond to modern designations [2]. Investigations into runs of homozygosity in 1476 horses of European descent revealed positive selection for base coat color on ECA3, but not in the region harboring KIT [3], while selection for white coat color patterns has been identified in ancient horse DNA [4].
The Dominant White locus is the equine locus with the largest number of known variants causing depigmentation (Table 2), and it is located on chromosome 3 within the Proto-Oncogene, Receptor Tyrosine Kinase (KIT) gene [2]. Dominant White uses the capital “W” followed by the integer in the series to indicate the specific dominant variant present in the genotype (e.g., W35). Originally, the W symbol was used for a small number of variants all following a true dominant pattern of inheritance and producing all white horses in the heterozygous state. The earlier dominant mutations were not observed in the homozygous state, leading to the adoption of an alternate term, Lethal Dominant White. Time has not honored this tradition as, to date, six W variants are known to be inherited in an incomplete dominant manner, with genotypes existing in the homozygous state in apparently healthy horses. Recent publications have started to refer to Dominant White as White Spotting to better account for the varied phenotypes at the W locus [5,6,7,8]. Phenotypes at the Dominant White locus are broadly characterized by horses displaying white areas with clear borders, or a completely white horse with pink skin underneath. Thirty-five W variants have been reported, and it seems likely that the number will continue to increase [2,9,10,11,12,13,14,15,16,17,18,19,20].
Many W alleles have been traced to a founding individual and are limited to those descendants, yet others are observed in diverse breeds, including a few that have likely circulated among diverse breeds over centuries, transmitted in cross-breed matings and by shipment of horses around the globe. The introduction of white alleles to new breeds is also promoted by registries opening studbooks to foreign horses with hopes of reducing inbreeding. As an example, researchers identified W13 in American Quarter Horses (AQH) in 2011 [14], but more recently observed this allele in both Shetland ponies and American Miniatures [21], two breeds genetically distant from the AQH. As Shetland ponies are not typically tested for W13, identification of this allele only occurred after genotypes for more common white variants (ex: TO, SB1, W20) failed to explain the depigmentation. Identifying variants outside of their presumed breed of origin is important to ensure the accurate monitoring and reporting of alleles with harmful side effects. Increased awareness of the presence of these alleles in new and existing populations will help prevent the introduction of white alleles into registries that select against white markings and mitigate the potential crossing of lethal pairs.
2.1. Phenotype
Phenotypes associated with the W locus are characterized by either white patterning or an entirely white coat with pink skin underneath. Mild white spotting phenotypes are described as sabino-like, with white legs, facial stripes, and a collection of other white facial markings (called stars or snips depending on the size, shape, and location), and, less commonly, patches of white hair across the abdomen [22]. Strongly deleterious mutations (frameshift, stop-gain, indel) typically result in completely white horses with pink skin (Figure 1). In contrast, W20, W32, W34, and W35 horses can be solid (non-white) in color in the absence of other white alleles but may magnify white markings caused by other alleles. For example, when an individual carries one copy of W22 and an out-of-phase copy of W20, the resulting phenotype is an all-white or almost all-white horse, despite each of these individual alleles typically producing less pronounced depigmentation phenotypes [11,23]. The amplification of the degree of white spotting is also observed with W5/n and W20/n compound heterozygotes, as these horses display an all-white phenotype [24]. Variants W19, W21–W23, W28, and W31–W35 produce a sabino-like phenotype sometimes accompanied by depigmentation on the abdomen with jagged borders. KIT variants also sometimes cause a rare phenotype of blue eyes when the depigmentation covers the entire face, including the eyes.
Deleterious Dominant White alleles result in more extensive white markings and are likely lethal in the homozygous state [25]. W1–W14, W16–W18, W21–28, W30, W31, and W33 have not been observed in the homozygous state, and are predicted to be homozygous lethal due to their similarities to mutations observed in other species [2,25]. Progeny ratios for white alleles causing fully white phenotypes also stray from Mendelian expectations. When heterozygous white horses were crossed, the resulting offspring possessed a 2:1 ratio of white foals to solid foals, supporting the hypothesis that W/W is lethal during early gestation [25]. These two observations suggest homozygous embryos are not viable for certain alleles, but too few births have occurred to conclusively determine the lethality of each variant. W15, originally thought to be embryonic lethal, was later reported in two homozygous individuals [18]. A horse homozygous for W19 was also recently identified, which also boasted two copies of W34 and W35 each, for a total of six white spotting variants [26]. Cases such as this support the hypothesis that other white variants could be viable in the homozygous state but have not yet been observed. Conclusions regarding the lethality of homozygous Dominant White variants will only be elucidated through continued monitoring and expanded genetic testing for these variants.
Many KIT variant haplotypes are reported in horses, and the resulting phenotypes are extremely varied and not fully documented. Phenotypes of horses with multiple white alleles depend on the specific white allele combination but generally result in increased depigmentation when compared to individuals with only one variant. Despite the impressive number of publications on the Dominant White locus, there are few studies focusing on phenotypes of horses with multiple white alleles. There are even fewer studies focusing on the health effects, and specifically, the reproductive effects, of horses with KIT variants, despite reports of KIT variants being associated with health defects in other species [2].
2.2. Mechanisms and Genetics
KIT transmits transmembrane signals critical for survival and plays an important role in melanogenesis [27,28,29]. During development, melanoblasts begin to migrate from the neural crest to populate the rest of the body and eventually develop into melanocytes (pigment-producing cells). Melanocyte development is in part controlled by interactions between KIT, KIT Ligand (KITL), and Melanocyte Inducing Transcription Factor (MITF) [28,29,30,31,32,33,34,35]. After binding with KITL in the extracellular domain, KIT self-dimerizes and phosphorylates MITF, activating the transcription factor and upregulating target genes involved in pigmentation [28,29]. Mutations affecting the function or binding sites of KIT protein disrupt this pathway, resulting in downregulated pigment genes and melanocytes failing to develop in some or all of the tissues.
There are a variety of mutations at the Dominant White locus including deletions, insertions, missense, nonsense, and splice site variants. More impactful mutations alter the protein conformation and function to a greater degree, and cause greater disruptions to KIT pathways, resulting in fewer melanoblasts properly migrating and a more depigmented individual. More tolerated KIT variants such as W20, W32, and W35 have subtle effects on the protein or protein expression and result in milder phenotypes. However, because the failure of a melanoblast to migrate is a chance event, mild variants on their own may still cause extensive depigmentation. The stochastic nature of white spotting events can cause individuals with the same genotype to display very different phenotypes. Commercial genetic testing for all W alleles exists, but assays for W10, W13, W19, W20, and W22 are among the more readily available tests since these alleles are more common.
Up to three KIT variants have recently been found linked together, resulting in complex haplotypes. To date, the W22 allele has only been observed in combination with the W20 allele [11,23]. W19 has been observed by itself and in linkage with W34 and W35. The W19W34W35 haplotype likely occurred by a crossover event because it was only identified in an inbred family, while the W19 allele has been found out of phase of W34 and W35 in multiple families [26]. Sixteen haplotypes, including combinations of W20, W32, W34, and/or W35 with other variants, have been identified, but the genesis of these complex haplotypes is not completely understood. Founder horses have not been reanalyzed for recently discovered alleles to reveal if novel variants occurred on the background of other alleles or if the haplotype occurred via a crossover event. While phenotypic records of all known multilocus genotypes are incomplete, it is likely that more white variants increase the amount of white patterning on a horse.
3. Tobiano, Sabino, and Roan
Tobiano, Sabino, and Roan are three alleles that have adopted the name of the respective phenotype. All three were independently mapped to ECA3 near the KIT gene [9,27,36,37,38,39]. Tobiano and Sabino are two well-characterized variants whose origins trace back to ancient times. Conversely, Roan is poorly understood, and no causative variant has been identified to date, though it has been mapped to the KIT region. All three variants cause varying degrees of white phenotypes in the heterozygous state without disrupting the base coat color. Homozygous Sabino 1 horses are mostly all white, while Tobiano homozygotes display varied amounts of depigmentation. Although roaning can be localized to a region or cover the entire body, the Roan allele is specifically characterized by roaning covering the body of the horse, leaving the head and extremities dark. The genomic coordinates of these three variants are shown in Table 3.
3.1. Tobiano
The nomenclature for Tobiano uses “TO” to represent the presence of the dominant allele and “to” to indicate the wild type allele. The mutation causing the tobiano phenotype, characterized by large white areas with smooth borders on the body and legs, has been mapped just outside of the KIT gene [1,36]. The oldest horse identified with the Tobiano allele dates between 3500 and 3000 years ago in Eastern Europe. The frequency of the TO allele has changed throughout history. During early domestication, the frequency of Tobiano began to rise, but it fell during the Middle Ages, as humans selectively bred in favor of other coats [4,40].
3.1.1. Phenotype
Classic tobiano, minimal tobiano, and cryptic tobiano are terms used to describe the diversity of phenotypes displayed by individuals with the Tobiano allele (Figure 2). Classic tobiano horses display large patches of white spotting, with clear borders [1,24,36,41]. Classic markings often spread over the flank, stretch up the neck, and cross the back. Classic tobiano horses generally have four white legs below the knees, but these marks can extend higher. Minimal tobiano, as the name suggests, is characterized by fewer white markings on the body, similar in pattern to classic tobiano but smaller in distribution. Cryptic tobiano horses display little to no white markings on the body, a phenotype easily confused for solid horses with white socks [41]. This presents a challenge in breeding when genotypes are unknown and has led to inaccurate heredity reporting in studbooks [24]. Cryptic tobiano individuals mistaken for solid horses with socks can also introduce the Tobiano variant into studbooks where the trait should be excluded [41]. Tobiano horses express their base coat color (chestnut, black, bay) in pigmented areas and have black skin beneath; however, the skin beneath white areas is pink. Individuals displaying tobiano characteristics often have a mix of white and base coat-colored tails. This phenotype is unique to Tobiano and is not expected for any other known white spotting trait [1,36]. Homozygous Tobiano horses may display “ink spots,” which are small dark spots within white areas, but the TO/TO genotype does not explain all cases of ink spots [1]. Tobiano horses with other white spotting variants often express an additive phenotypic effect and are more white than horses with Tobiano alone.
3.1.2. Genetics and Mechanism
The mutation causing the tobiano phenotype is a large paracentric inversion of a ~43 Mb region upstream of KIT and is expressed in a dominant manner [1,36]. This region, spanning nearly a third of the chromosome, is thought to harbor genetic machinery regulating KIT and its relocation is believed to reduce KIT expression. The KIT structure is not directly affected by this inversion, allowing for proper protein function, which may explain the possibility of the cryptic tobiano phenotype. Many homozygous TO/TO individuals have been documented and none have been reported to have any health defects due to the TO allele [1,36]. The TO mutation may result in reduced fertility rates, as large chromosomal inversions typically make gametogenesis more difficult, but this has not been studied [36]. The Tobiano allele is found in diverse registries including but not limited to: Paint, Pinto, Shetland, Miniature, Warmblood, Gypsy, and Paso Fino. Conversely, the trait is not commonly found in American Quarter Horses, Thoroughbreds, Standardbreds, or Arabians. Commercial genetic tests for Tobiano are readily available.
3.2. Sabino
The sabino phenotype is the earliest known white spotting phenotype and is one of the best characterized white spotting mutations. The genetic variant responsible for Sabino 1 traces back to sometime between 5500 and 5000 years ago, first appearing during early domestication on the Siberian Steppe and later found in Armenia and Moldavia [4,40]. The sabino trait uses the symbol “SB1” to represent the presence of Sabino and “sb1” or “n” to indicate the absence of Sabino. The phenotype produced by the Sabino mutation manifests in a manner similar to Dominant White mutations like W19 and W31. These visual similarities can cause confusion between Sabino and other white-causing traits. Genetic testing is the best way, and sometimes the only way, to differentiate a true Sabino from a Dominant White horse.
3.2.1. Phenotype
The sabino phenotype is characterized by extensive depigmentation of the legs, face, and abdomen [1,9]. Although the sabino phenotype is attributed to the Sabino 1 allele, Dominant White variants can also produce sabino-like phenotypes, making distinctions between sabino phenotypes and other white spotting phenotypes challenging. Heterozygous Sabino horses display irregular, jagged borders to their white markings, but homozygous horses are fully or nearly fully white (Figure 3). SB1/n individuals may have white speckles near the borders of larger areas but they are typically surrounded by pigmented areas with clear borders [1,9]. Pink skin is found below the white areas and on the face, while skin below pigmented hair is normally pigmented. Homozygous individuals and heterozygotes with extensive white spotting are more sensitive to light and should be monitored closely when exposed to direct sunlight for prolonged periods. Horses with SB1 may express a minimal sabino phenotype with low white socks similar to cryptic tobiano. Horses with SB1/n and other white spotting variants typically express more white than horses with SB1/n alone.
3.2.2. Genetics and Mechanism
The Sabino phenotype is inherited in an incompletely dominant manner and is caused by a splice site SNP in KIT intron 16 [1,9]. This splice site variant produces a unique transcript lacking the 17th exon, which encodes part of the active kinase domain. Without this critical exon, KIT may fold in a less effective manner or be completely inactive. Having less active KIT results in fewer melanoblast signaling events and less developed melanocytes, resulting in the absence of pigment with a white spotting or all white phenotype, depending on zygosity. However, because some copies of the normal transcript are still expressed, heterozygous and homozygous horses develop into healthy individuals with no reported health defects [1,9]. Genetic tests for the sabino mutation are readily available.
SB1 is an old mutation and has had many generations to diffuse through breeds and combine with other white spotting mutations [1,4,9,40]. Haplotypes containing the SB1 allele and the W20 and/or W32 have been identified [26]. The Sabino mutation is found in a diverse set of breeds, many of which are of Spanish origin. Although a sabino-type phenotype is found in Clydesdale horses, the SB1 variant is absent, indicating multiple pathways lead to phenocopies.
3.3. Roan
Although the phenotype caused by the Roan allele is well characterized, the genetic cause remains a mystery. The symbol “RN” is used to indicate the presence of the allele and “rn” or “n” to indicate its absence [1]. The roan phenotype is characterized by a mixture of white hairs diffused throughout the body, while the head, lower limbs, skin, mane, and tail remain solid in color (Figure 4). “Blue roan” is used to describe the roan phenotype for horses with a black base coat color and “red roan” is reserved to describe the roan phenotype on a chestnut coat. The phenotype associated with the Roan allele can often be confused with varnish roan, which is attributed to the LP allele. The Grey phenotype may also be confused for Roan, with the most notable difference being that the Grey trait progressively lightens the coat with age, whereas Roan individuals are born with white ticked coats. So far, only anecdotal evidence exists for homozygous Roan horses, since the trait does not yet have a well-documented mode of inheritance or a known causative variant. Pedigree analysis suggests that Roan is inherited in a dominant manner [37,38,39].
Multiple studies observed the association of Roan to markers within the KIT gene on ECA3, but there is no clear consensus regarding any single SNP most strongly associated with the roaning phenotype. KIT is the gene most likely to harbor the variant causing roan but a causal mutation has not been identified [39]. They excluded an intronic insertion of a LINE as the causative mutation after discovering it was common in all horses. The team also identified a synonymous mutation in exon 19 strongly associated with the roaning phenotype in all breeds except Shetland and Gotland ponies [39]. A genetic marker in the 17th intron of KIT was found to be the most associated with the Roan phenotype [37]. The proposed genetic marker was completely associated with Roan in Noriker horses and is suggestively associated in Quarter horses and various draught breeds [37]. Yet, this marker fails to explain the roaning phenotype in all breeds, as Shetland ponies with the Roan phenotype did not possess the associated marker. The breed segregation of these discoveries and conflicting conclusions regarding lethal Roan suggests there may be allelic heterogeneity for Roan or the presence of multiple phenocopies.
Roaning in canines is caused by a tandem intronic duplication in the USH2A gene [42], and in mice by a deletion in the mouse ortholog of KITL, the Steel Ligand Factor (SLF) gene, encoded by the Steel locus (reviewed in 60). These reports suggest the Roan allele in horses may not be exclusively controlled by mutations in KIT, but neither of these comparative candidate genes lies on ECA3 and would therefore not likely be involved in the Roan phenotypes within breeds where an association with ECA3 was observed. If the roan phenotype is caused by multiple variants, or by epistatic interactions between multiple loci, many of the existing disagreements about Roan could be explained. Top-associated SNPs differ between breeds and studies, bringing into question the validity of currently marketed Roan assays, as these are easily applied inappropriately to populations in which the association of these markers to the Roan phenotype is unknown.
4. Splashed White
The Splashed White collection is the second largest grouping of genetic variants causing depigmentation in horses [15,43,44,45,46,47,48]. The capital letters “SW” followed by the number in the series (e.g., SW10) is used to indicate a mutant allele, with an “n” or “+” used to indicate the wild-type allele [1]. There are ten reported variants in the Splashed White grouping, each affecting one of two transcription factors: PAX3 (Paired-Box 3) or MITF. Although these two genes are completely independent loci, and play different roles in the cell, mutations in either gene causes phenotypes without distinguishable differences, leading both to be included in the Splashed White naming convention (Table 4). The Splashed White trait was named after the resulting phenotype, as affected horses feature patches of white that make it look as if they have been dipped in or “splashed” from below with white paint. White markings typically exist on all four legs extending up to and sometimes covering the abdomen and the face.
4.1. Phenotype
As mentioned above, the phenotype characterizing Splashed White features flashy white legs with additional white markings on the abdomen that can extend all the way up to the body. Some variants produce fully white abdomens, with smooth borders and little to no roaning or speckling, as if the horse dipped its belly in white paint [1]. These horses often also demonstrate all white, “paint dipped” faces: a facial phenotype sometimes referred to as “bald.” In horses with Splashed White, markings can extend beyond the eyes and ears (Figure 5), accompanied by blue or partially blue eye(s). There are no major differences between Splashed White phenotypes caused by mutations in MITF and PAX3, and mutant alleles for either gene are tied to an increased risk of deafness [43,44,45,46,48]. Depigmented individuals may be more sensitive to sunlight and require more care when in direct sunlight for longer periods. Combinations of SW variants are less frequent than combinations of Dominant White alleles, so less is known about their genetic interplay and resulting phenotypes. However, in some observed cases where at least one MITF SW variant and at least one PAX3 SW variant are inherited (most commonly, SW1/n SW2/n), or two MITF SW variants are inherited (SW1/n SW7/n), horses can be born with an all-white or nearly all-white coat and can be deaf depending on the variants inherited [44,48].
4.2. MITF
MITF encodes a transcription factor necessary for proper cell development [29,30,31,32,49]. MITF contains one basic helix-loop-helix and one leucine zipper domain, providing specificity to and affinity for its DNA target sites [35,50]. MITF is phosphorylated by KIT during melanogenesis to direct the gene expression pathways that differentiate melanoblasts into melanocytes and later induce the expression of other pigment genes such as TYR, TYRP1, PMEL, and MLANA [34]. Similar to other mutations found within DNA binding domains, the change in protein structure causes reduced affinity or specificity in binding, causing MITF to stray from its typical targets and pathways.
Seven of the ten Splashed White alleles are MITF mutations on ECA16 [43,44,45,46,47,48]. Most of these alleles are inherited in a dominant manner, meaning individuals with one mutant allele typically demonstrate a Splashed White phenotype. For most of these mutations, homozygous individuals have not yet been observed, and this, in combination with the dominant inheritance pattern, implies that these mutations could be embryonic homozygous lethal. To date, the only MITF allele observed in the homozygous state is SW1, which is inherited in an incomplete dominant manner. Homozygous individuals display typical splashed markings, but heterozygotes display less extensive depigmentation [1]. SW1/SW3, SW1/SW2, SW1/SW5, and SW1/SW7 horses have been observed and display all white phenotypes with signs of deafness [44,48]. No homozygous individuals have been described for MITF Splashed White mutations outside of SW1. Most MITF variants are frameshift indels with significant impact and likely lethal in the homozygous state, which may explain why homozygous individuals are not observed. While SW1 is found in numerous breeds, the remaining MITF alleles are breed specific. SW9 is only found in a single family of Pura Raza Española horses, while SW3, SW5, SW6, and SW7 have only ever been reported in Paint and Quarter horses [44,45,46,48]. SW8 was found in a single Thoroughbred stallion and one of his offspring [43]. Parentage testing confirmed the Thoroughbred proband as the SW8 founder.
There are three MITF mutations associated with depigmentation without an SW designation [44,51]. Macchiato is a variant of MITF (ECA16:21564980T>C) found in one sterile male displaying a light coffee color coat and white spotting similar to phenotypes attributed to Splashed White alleles [44]. The term Macchiato was assigned to this trait because the coat color dilution affects the base coat, while SW does not. A de novo MITF variant predicted to alter protein sequence (ECA16:21556522C>T) was identified in a single white American Standardbred [51]. Because neither mutation was likely to be passed onto future generations, they were not given an SW designation. An intronic variant (ECA16:21608936C>T) was associated with forelimb white markings in Menorca Purebred horses and facial white markings in Pura Raza Española horses [52]. The intronic variant is located 29.9 kb downstream from the transcription start site and is speculated to either affect the regulation of MITF or is linked to another variant responsible for causing white markings [52]. Although this variant was not given an SW designation, it is still a useful marker to select for white coat color in Spanish breeds. The linkage of the ECA16:21608936C>T variant to white markings has not been studied in other breeds.
4.3. PAX3
PAX3 belongs to the Paired Box family of transcription factors characterized by a paired-type homeodomain and paired box domain [50,53,54]. Transcription factors in the PAX family are well understood to regulate transcription during development. PAX proteins contain an octapeptide sequence, which helps maintain DNA–protein interactions and conserve binding domains [29]. The paired box domain, as the name would imply, is the most conserved domain in the PAX family. Mutations in PAX genes are associated with hypopigmentation and deafness in horses and developmental abnormalities in humans and flies [1,54]. In horses, mutations in PAX3 are thought to interrupt the encoded transcription factor’s affinity and/or specificity to bind to MITF. Without proper PAX3 interaction, MITF is under expressed, causing down-regulation of its target pigment proteins and eventual failure of melanoblast proliferation and differentiation [30,49,55]. The regulatory interactions between PAX3 and MITF partly explains how mutations in either gene could result in a splashed white phenotype.
Three of the ten Splashed White alleles are attributed to mutations in PAX3 on ECA6 and cause depigmentation of the hair, skin, and eyes as well as an increased risk for deafness [15,44,47]. Of the Splashed White PAX3 variants, only SW2 has been observed in the homozygous state, resulting in all white phenotypes. SW2 is inherited in an incomplete dominant manner with heterozygous individuals displaying typical Splashed White markings. SW4 is predicted to be nonviable in the homozygous state due to a lack of observed homozygotes. SW10 is also predicted to be embryonic lethal in the homozygous state since it terminates the transcript before a crucial DNA binding domain via an early stop-gain missense mutation. However, too few births have occurred to definitively conclude lethality in both cases. SW2 and SW4 have been observed in Quarter horses and Appaloosas, respectively [15,44]. To date, SW10 has been observed in two Pura Raza Española horses [47]. Interestingly, the families with either SW9 or SW10 have been bred together, making fertilization of a SW9/n SW10/n zygote possible, although no individuals with this genotype have been observed as of yet, if even viable. It is unclear whether combinations of PAX3 mutations are viable, as all alleles exist in low frequency.
5. Eden White and HPS5
Eden White is a recently reported grouping of white spotting variants associated with Hermansky-Pudlak Syndrome 5 (HPS5) on ECA7 [56]. Relatively little is known about the function of this encoding protein, but mutations in HPS5 cause depigmentation in humans, mice, and other organisms [56]. HPS5 encodes a member of Biogenesis of Lysosome Related Organelles Complex 2 (BLOC-2) with members HPS3 and HPS6, but the exact function of the complex is unknown. It is therefore unknown how mutations in HPS5 affect the complex’s function and result in depigmentation. HPS5 has one annotated domain, WD40, thought to control interprotein interactions in BLOC-2 [56]. Although the exact function of the BLOC-2 complex is unknown, it is suspected to play a role in organelle biogenesis associated with melanosomes, lysosomes, and platelet-dense granules [56]. In humans, mutations in HPS5 cause Hermansky–Pudlak syndrome type 5, characterized by depigmentation and a reduction of platelet-dense granules, resulting in prolonged bleeding times [56]. Mutations in HPS5 have also been reported to cause albinism in zebrafish [56].
Three variants within HPS5, termed Eden White 1 through Eden White 3 (EDXW1-EDXW3), have been associated with depigmentation in horses (Table 5). The phenotypes of affected horses are similar to Splashed White phenotypes, featuring sabino-like markings, long blazes, and socks with distinct borders (Figure 6). EDXW variants may also produce white marks on the abdomen and blue eyes (Figure 6A), but these are more rare phenotypes, and are usually only found among homozygous or compound heterozygous individuals. The symbol EDXW followed by the number in the series (e.g., EDXW2) is used for the mutant while the letter “n” is used to indicate the absence of any EDXW variant. Horses of all major breed groups have been found to possess Eden White alleles, but heavy horses and stock-type horses are most commonly found with these variants. To date, no health defects have been reported for horses with any of the Eden White variants, but this has not been thoroughly investigated.
Although it is possible to have combinations of Eden White variants, including compound homozygous, there are no reported trends for the phenotypes of these horses [56]. It has been observed that they display more white spotting but too few individuals have been phenotyped to test this hypothesis statistically. It is likely that the more variants possessed, the more white markings will be displayed. Eden White 1 and Eden White 3 are epistatic to MC1R, which controls the chestnut or black coat color in horses, similar to most mammals. Individuals with a dominant black base coat color (E/e or E/E) display more white markings than individuals with a chestnut coat color (e/e), a trend not yet documented for EDXW2, but likely consistent among all non-tolerated HPS5 variants, similar to findings in other species [56].
6. Lethal White Overo
Lethal White Overo, commonly known as Frame Overo, is the phenotype displayed by heterozygotes possessing the LWO variant. This allele is symbolized by “LWO” or “O” while “n” or “o” are used to indicate the absence of the mutation; however, other texts have used other symbols, including [1]. Horses displaying frame overo can have strong facial markings similar to Splashed White individuals, with their face appearing to be bald or dipped in paint (Figure 7) [57]. LWO horses display irregular framed depigmented patches on the body, but the markings typically do not extend below the body or cross the spine. Horses with LWO may have blue or partially blue eyes and may be predominantly white or their base coat color with minimal or no white markings in the heterozygous state [57]. In one population, 18% of solid breeding stock horses were heterozygous for the LWO allele, indicating that the frame overo phenotype is not always apparent in LWO heterozygotes and other factors might influence the extent of white spotting [57]. Horses born with two copies of LWO are entirely white and with a megacolon, which prompts immediate euthanasia [1,57]. The mutant allele is most common in American Paint Horses and proof of LWO genotype is one of two alleles acceptable for registry in the American Paint Horse Association [57]. Individuals with LWO and other white spotting mutations typically display larger white spots than individuals with only one copy of LWO [1]. The variable phenotypes of LWO and other white spotting mutations has led to the misclassification of heterozygous LWO horses as splashed white, tobiano, sabino, or their phenotypic derivatives [e.g., tovero, calico, and frame blend] [57].
A double nucleotide polymorphism (chr17:50503041-50503042delinsCT) in the first exon of Endothelin Receptor B (EDNRB) on ECA17 is responsible for LWO. EDNRB encodes a g-coupled receptor responsible for the migration of neural crest cells during early development. In embryos, the protein helps neural crest-derived cells develop into melanocytes, as well as nerves within the intestines [1,58]. The dinucleotide polymorphism occurs at codon 118, within a transmembrane domain, and exchanges a nonpolar isoleucine to positively charged lysine [58]. The exchange of the evolutionarily conserved isoleucine is thought to affect EDNRB morphology, affecting how it folds or how it anchors itself in the lipid bilayer [58]. LWO syndrome in horses is synonymous with the Hirschsprung and Waardenburg Syndromes in humans, which are similarly attributed to mutations in human EDNRB [57,58]. These diseases are characterized by developmental issues and depigmented eyes, hair, and skin. Humans, mice, and horses with EDNRB-associated diseases all exhibit similar phenotypes.
The phenotypes of homozygous LWO horses are observable from birth. The most apparent and immediate are the fully white coat and blue eyes, but the adverse health effects are observable soon after. Most homozygous foals show signs of intestinal discomfort within several hours of birth. These foals usually do not pass meconium, and neither surgery nor medication help to bypass the occlusion [1,58]. Affected foals are missing intestinal ganglia that control the peristaltic muscles responsible for peristalsis throughout the intestines, meaning they cannot pass food through their digestive tract. However, not every all-white foal born is homozygous for LWO, nor does every horse heterozygous for LWO display depigmentation phenotypes. For these two reasons, readily available commercial genetic testing for LWO is the most effective method for determining the Frame Overo genotype and limiting instances of breeding that could lead to lethal white syndrome-affected foals.
7. Leopard Complex Spotting
Leopard Complex spotting, commonly referred to by the breed most known for this color, the “Appaloosa,” is an incompletely dominant phenotype caused by a mutation in the Transient Receptor Potential Cation Channel Subfamily Member 1 (TRPM1) gene. The symbol “LP” is used for the incompletely dominant allele and lowercase “lp” is used to indicate the recessive wild type allele. The oldest genetic evidence of leopard spotting could be traced to the Pleistocene, but negative selection for the trait post-domestication reduced its frequency [4]. The protein encoded by TRPM1 mediates the flux of ions across membranes in the brain, heart, and melanocytes to modulate cell polarization and is thought to play a role in melanin synthesis [59]. TRPM1 is partly responsible for vision in low-light settings as visual signals from rod centers trigger the protein channel to close [59]. Leopard Complex spotting is caused by a retroviral insertion found within TRPM1 (Table 6) that interrupts protein function [1,27]. The mutant allele contains a premature poly-adenylation signal, causing the expression of a non-functional TRPM1 isoform. The resulting protein lacks the ability to respond to regulatory signals normally controlled by glutamate. Without this response, TRPM1 cannot fulfill its normal functions, severely impacting pathways including sight and pigment. The hypothesized mechanism by which TRPM1 variants might cause albinism suggests that the mutant transcript causes morphological defects in melanocytes, which causes melanocyte death [27].
Horses with an LP allele typically display a depigmented hind quarter with pigmented spots (Figure 8), but can display more pronounced depigmentation phenotypes extending to beyond the hind quarters onto the body and neck or very minimal phenotypes of roan-like patterning or solid with little to no white markings at all. The LP phenotype of a white hind quarter with pigmented spots is commonly referred to as a blanket, due to its similarity to a polka-dotted blanket draped over a horse. The extent of depigmentation due to the LP allele is highly variable, including other phenotypic characteristics of visible white sclera, striped hooves, mottling (depigmentation/pink skin around the face, genitalia and anus), and varnish roaning (a gradual loss of pigmentation throughout the coat but not on bony surfaces) [1,27]. Horses with LP/LP generally display much more white and fewer spots than heterozygotes. White markings for LP do not generally produce the clear borders observed for other white spotting mutations. White areas can appear in patches with jagged borders and have white hairs dispersed in the surrounding area. The extent of white spotting is influenced by other factors, including sex, the Extension genotype, the PATN1 genotype, and the accumulation of mutations in either KIT or MITF [59]. Although LP is the signature phenotype of the Appaloosa horse breed, the variant is found in several other horse breeds, including but not limited to, Knabbstrupper, Ponies of America, Paint, and Quarter Horses [1].
7.1. Congenital Stationary Night Blindness
The phenotypic health effect of the retroviral insertion responsible for LP is only observed in homozygous individuals, and the LP/LP genotype is the most common cause of Congenital Stationary Night Blindness (CSNB) in horses [1,27]. It is thought that the mutant TRPM1 lacks the necessary machinery to properly respond to ocular signals produced in low light levels [27]. This null response causes homozygous individuals to be born with a lack of vision in the dark that does not progress with age. Although horses can adapt well to CSNB, they are more prone to injury during the night if sufficient light is not provided. CSNB can be difficult to diagnose based on a highly variable phenotype and because horses can be well adapted to the condition. For these reasons, genetic testing is the most recommended method to verify an individual’s LP genotype and a possible diagnosis of CSNB.
7.2. Equine Recurrent Uveitis
Equine recurrent uveitis (ERU) is an ocular disease associated with LP in horses that may cause the development of cataracts, glaucoma, and blindness [60,61]. ERU differs from CSNB in multiple characteristics: 1. ERU usually worsens with age, 2. Horses are not born with symptoms of ERU, and 3. ERU is triggered by environmental and genetic factors as well as age [60,61]. The genetic component of ERU was associated with LP in multiple studies, but the association does not explain all cases of ERU [60,61]. There are likely other undiscovered traits or environmental factors also contributing to ERU risk. Horses with the LP/LP genotype are at a higher risk for developing ERU than heterozygotes. Due to the high frequency of LP in Appaloosa horses, they are eight times more likely to develop ERU than other breeds [60]. It has been predicted that periocular depigmentation might underlie the higher risk for ERU despite findings indicating varnish roan phenotype is not a significant predictor [60]. Commercial genetic testing for LP is available for determining an individual’s ERU risk, although the presence of this allele does not guarantee a horse will develop ERU.
7.3. RFWD3 and PATN1
The LP allele is epistatically influenced by the Pattern-1 allele, located within the Ring Finger and WD Repeat Domain 3 (RFWD3) gene on ECA3 [1,59]. PATN1 is the symbol used to represent the dominant non-reference allele and “n” or “patn1” is used to indicate the absence of the variant. PATN1 is inherited in a dominant fashion as horses express the PATN1 phenotype in the presence of at least one LP allele regardless of PATN1 zygosity (Figure 8). The PATN1 phenotype has only been observed in LP/X horses (where X is either allele) and horses without LP appear the same regardless of the PATN1 genotype. Horses with LP/lp and PATN1/X display more LP-associated white markings while LP/LP PATN/X individuals are generally born nearly all white and with fewer spots. PATN1 is associated with an increased risk of ERU in individuals with at least one LP allele. Individuals with LP/LP PATN1/n are at the greatest risk of developing ERU [60,61]. More studies are needed to explain how PATN1 and ERU are related.
RFWD3 encodes an enzyme responsible for ubiquitin-protein ligase activity and is involved in several pathways including cell cycle regulation and DNA repair. In response to DNA damage, the encoding protein is recruited to form a complex with Mdm2, another ubiquitin ligase. A SNP mutation in the 3′UTR of RFWD3 is strongly associated with the PATN-1 phenotype and is suspected to alter the protein’s expression. RFWD3 is thought to be involved in the removal of deformed melanosomes observed in LP horses, either directly or indirectly. Alterations in its expression could lead to earlier melanocyte death and removal, resulting in more extensive depigmentation in the presence of at least one LP and one PATN1 allele [59].
8. Conclusions and Future Perspective
There have been huge advancements in equine genetics in recent years, and new white variants, coat color dilutions, and health-associated loci are being reported every year. However, there remain many poorly understood pigmentation traits requiring more research to uncover their genetic causes. For example, despite linkage of this phenotype to KIT in some breeds, the mechanisms underlying Roan are still poorly understood. Rabicano is characterized by white ticking on the flank, sometimes extending forward toward the barrel, and white banding on the tailhead, and has been linked to variants in possible regulatory regions affecting KITLG [62], but the genetic cause for Rabicano is still unknown. Few studies have focused on the health or reproduction of horses with multiple KIT variants. It has not been confirmed if homozygosity of multiple Dominant White alleles is truly lethal. More evidence on the deafness of overo and splashed white horses is needed to validate the anecdotal stories of deaf horses and to come to a conclusion about the cause of the hearing loss. It is important to continuously monitor the presence of LP, PATN1, and LWO variants in horse populations to mitigate adverse health effects. Studies are needed to elucidate the function of BLOC-2 and other HPS genes, which will help to improve the prognosis of HPS-related diseases in humans and reveal if horses suffer from the same defects due to HPS mutations. Understanding the etiology of equine coat color is crucial to improve strategies to breed healthier and more beautiful horses while controlling the allele frequencies of variants associated with adverse health effects.
Author Contributions
Conceptualization, A.M. and R.E.E.; Data curation, A.M.; Formal analysis, A.M.; Project administration, K.M., R.E.E. and C.L.; Resources, C.L.; Supervision, K.M., S.A.B. and R.E.E.; Validation, S.A.B. and R.E.E.; Visualization, A.M.; Writing—original draft, A.M.; Writing—review and editing, M.V., S.A.B. and R.E.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable as no experiments were performed during the course of this review.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank all horse owners; you are the true heroes. Without your curiosity and determination, the world of equine genetics would not exist as it is today. We would like to thank all horse owners who submitted photos of their horses for this review and specifically thank Jack Creek Appaloosas for the photographs used in Figure 8B–D.
Conflicts of Interest
Authors A.M., K.M., M.V., R.E.E. and C.L. are affiliated with Etalon, Inc., which offers genetic testing for many pigmentation-related variants, including the variants described in this review.
References
- Bailey, E.; Brooks, S.A. Horse Genetics, 3rd ed.; CABI: Wallingford, UK, 2020. [Google Scholar]
- Haase, B.; Brooks, S.A.; Schlumbaum, A.; Azor, P.J.; Bailey, E.; Alaeddine, F.; Mevissen, M.; Burger, D.; Poncet, P.-A.; Rieder, S.; et al. Allelic Heterogeneity at the Equine KIT Locus in Dominant White (W) Horses. PLoS Genet. 2007, 3, e195. [Google Scholar] [CrossRef]
- Grilz-Seger, G.; Neuditschko, M.; Ricard, A.; Velie, B.; Lindgren, G.; Mesarič, M.; Cotman, M.; Horna, M.; Dobretsberger, M.; Brem, G.; et al. Genome-Wide Homozygosity Patterns and Evidence for Selection in a Set of European and Near Eastern Horse Breeds. Genes 2019, 10, 491. [Google Scholar] [CrossRef]
- He, Z.; Dai, X.; Lyu, W.; Beaumont, M.; Yu, F. Estimating Temporally Variable Selection Intensity from Ancient DNA Data. Mol. Biol. Evol. 2023, 40, msad008. [Google Scholar] [CrossRef]
- Esdaile, E.; Till, B.; Kallenberg, A.; Fremeux, M.; Bickel, L.; Bellone, R.R. A de novo missense mutation in KIT is responsible for dominant white spotting phenotype in a Standardbred horse. Anim. Genet. 2022, 53, 534–537. [Google Scholar] [CrossRef]
- McFadden, A.; Martin, K.; Foster, G.; Vierra, M.; Lundquist, E.W.; Everts, R.E.; Martin, E.; Volz, E.; McLoone, K.; Brooks, S.A.; et al. 5′UTR Variant in KIT Associated With White Spotting in Horses. J. Equine Vet. Sci. 2023, 127, 104563. [Google Scholar] [CrossRef]
- Patterson Rosa, L.; Martin, K.; Vierra, M.; Foster, G.; Lundquist, E.; Brooks, S.A.; Lafayette, C. Two Variants of KIT Causing White Patterning in Stock-Type Horses. J. Hered. 2021, 112, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Patterson Rosa, L.; Martin, K.; Vierra, M.; Lundquist, E.; Foster, G.; Brooks, S.A.; Lafayette, C. A KIT Variant Associated with Increased White Spotting Epistatic to MC1R Genotype in Horses (Equus caballus). Animals 2022, 12, 1958. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Bailey, E. Exon skipping in the KIT gene causes a Sabino spotting pattern in horses. Mamm. Genome 2005, 16, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Capomaccio, S.; Milanesi, M.; Nocelli, C.; Giontella, A.; Verini-Supplizi, A.; Branca, M.; Silvestrelli, M.; Cappelli, K. Splicing site disruption in the KIT gene as strong candidate for white dominant phenotype in an Italian Trotter. Anim. Genet. 2017, 48, 727–728. [Google Scholar] [CrossRef] [PubMed]
- Dürig, N.; Jude, R.; Holl, H.; Brooks, S.A.; Lafayette, C.; Jagannathan, V.; Leeb, T. Whole genome sequencing reveals a novel deletion variant in the KIT gene in horses with white spotted coat colour phenotypes. Anim. Genet. 2017, 48, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Haase, B.; Brooks, S.A.; Tozaki, T.; Burger, D.; Poncet, P.-A.; Rieder, S.; Hasegawa, T.; Penedo, C.; Leeb, T. Seven novel KIT mutations in horses with white coat colour phenotypes. Anim. Genet. 2009, 40, 623–629. [Google Scholar] [CrossRef]
- Haase, B.; Jagannathan, V.; Rieder, S.; Leeb, T. A novel KIT variant in an Icelandic horse with white-spotted coat colour. Anim. Genet. 2015, 46, 466. [Google Scholar] [CrossRef]
- Haase, B.; Rieder, S.; Tozaki, T.; Hasegawa, T.; Penedo, M.C.T.; Jude, R.; Leeb, T. Five novel KIT mutations in horses with white coat colour phenotypes: BRIEF NOTE. Anim. Genet. 2011, 42, 337–339. [Google Scholar] [CrossRef]
- Hauswirth, R.; Jude, R.; Haase, B.; Bellone, R.R.; Archer, S.; Holl, H.; Brooks, S.A.; Tozaki, T.; Penedo, M.C.T.; Rieder, S.; et al. Novel variants in the KIT and PAX3 genes in horses with white-spotted coat colour phenotypes. Anim. Genet. 2013, 44, 763–765. [Google Scholar] [CrossRef]
- Hoban, R.; Castle, K.; Hamilton, N.; Haase, B. Novel KIT variants for dominant white in the Australian horse population. Anim. Genet. 2018, 49, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Holl, H.; Brooks, S.; Bailey, E. De novo mutation of KIT discovered as a result of a non-hereditary white coat colour pattern: De novo mutation of KIT. Anim. Genet. 2010, 41, 196–198. [Google Scholar] [CrossRef]
- Holl, H.M.; Brooks, S.A.; Carpenter, M.L.; Bustamante, C.D.; Lafayette, C. A novel splice mutation within equine KIT and the W15 allele in the homozygous state lead to all white coat color phenotypes. Anim. Genet. 2017, 48, 497–498. [Google Scholar] [CrossRef]
- Hug, P.; Jude, R.; Henkel, J.; Jagannathan, V.; Leeb, T. A novel KIT deletion variant in a German Riding Pony with white-spotting coat colour phenotype. Anim. Genet. 2019, 50, 761–763. [Google Scholar] [CrossRef]
- Martin, K.; Patterson Rosa, L.; Vierra, M.; Foster, G.; Brooks, S.A.; Lafayette, C. De novo mutation of KIT causes extensive coat white patterning in a family of Berber horses. Anim. Genet. 2021, 52, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Esdaile, E.; Kallenberg, A.; Avila, F.; Bellone, R.R. Identification of W13 in the American Miniature Horse and Shetland Pony Populations. Genes 2021, 12, 1985. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, F.; Tammen, I.; Hub, S.I. Online Mendelian Inheritance in Animals (OMIA). 1995. Available online: https://ses.library.usyd.edu.au/handle/2123/31190 (accessed on 17 July 2023).
- Avila, F.; Hughes, S.S.; Magdesian, K.G.; Penedo, M.C.T.; Bellone, R.R. Breed Distribution and Allele Frequencies of Base Coat Color, Dilution, and White Patterning Variants across 28 Horse Breeds. Genes 2022, 13, 1641. [Google Scholar] [CrossRef]
- Pasternak, M.; Krupiński, J.; Gurgul, A.; Bugno-Poniewierska, M. Genetic, historical and breeding aspects of the occurrence of the tobiano pattern and white markings in the Polish population of Hucul horses–A review. J. Appl. Anim. Res. 2020, 48, 21–27. [Google Scholar] [CrossRef]
- Pulos, W.L.; Hutt, F.B. Lethal Dominant White in Horses. J. Hered. 1969, 60, 59–63. [Google Scholar] [CrossRef] [PubMed]
- McFadden, A.; Vierra, M.; Robilliard, H.; Martin, K.; Brooks, S.A.; Everts, R.E.; Lafayette, C. Population Analysis Identifies 15 Multi-Variant Dominant White Haplotypes in Horses. Animals, 2023; Manuscript submitted for review. [Google Scholar]
- Bellone, R.R.; Holl, H.; Setaluri, V.; Devi, S.; Maddodi, N.; Archer, S.; Sandmeyer, L.; Ludwig, A.; Foerster, D.; Pruvost, M.; et al. Evidence for a retroviral insertion in TRPM1 as the cause of congenital stationary night blindness and leopard complex spotting in the horse. PLoS ONE 2013, 8, e78280. [Google Scholar] [CrossRef] [PubMed]
- Phung, B.; Sun, J.; Schepsky, A.; Steingrimsson, E.; Rönnstrand, L. C-KIT Signaling Depends on Microphthalmia-Associated Transcription Factor for Effects on Cell Proliferation. PLoS ONE 2011, 6, e24064. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.J.; Erickson, C.A. The making of a melanocyte: The specification of melanoblasts from the neural crest. Pigment. Cell Melanoma Res. 2008, 21, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Fuse, N.; Yasumoto, K.; Suzuki, H.; Takahashi, K.; Shibahara, S. Identification of a Melanocyte-Type Promoter of the Microphthalmia-Associated Transcription Factor Gene. Biochem. Biophys. Res. Commun. 1996, 219, 702–707. [Google Scholar] [CrossRef]
- Jones, S. An overview of the basic helix-loop-helix proteins. Genome Biol. 2004, 5, 226. [Google Scholar] [CrossRef]
- Kawakami, A.; Fisher, D.E. The master role of microphthalmia-associated transcription factor in melanocyte and melanoma biology. Lab. Investig. 2017, 97, 649–656. [Google Scholar] [CrossRef]
- Williams, D.E.; de Vries, P.; Namen, A.E.; Widmer, M.B.; Lyman, S.D. The Steel factor. Dev. Biol. 1992, 151, 368–376. [Google Scholar] [CrossRef]
- Yasumoto, K.; Yokoyama, K.; Shibata, K.; Tomita, Y.; Shibahara, S. Microphthalmia-associated transcription factor as a regulator for melanocyte-specific transcription of the human tyrosinase gene. Mol. Cell. Biol. 1994, 14, 8058–8070. [Google Scholar] [CrossRef]
- Yutzey, K.E.; Konieczny, S.F. Different E-box regulatory sequences are functionally distinct when placed within the context of the troponin I enhancer. Nucleic Acids Res. 1992, 20, 5105–5113. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Lear, T.L.; Adelson, D.L.; Bailey, E. A chromosome inversion near the KIT gene and the Tobiano spotting pattern in horses. Cytogenet. Genome Res. 2007, 119, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Grilz-Seger, G.; Reiter, S.; Neuditschko, M.; Wallner, B.; Rieder, S.; Leeb, T.; Jagannathan, V.; Mesarič, M.; Cotman, M.; Pausch, H.; et al. A Genome-Wide Association Analysis in Noriker Horses Identifies a SNP Associated With Roan Coat Color. J. Equine Vet. Sci. 2020, 88, 102950. [Google Scholar] [CrossRef] [PubMed]
- Hintz, H.F.; van Vleck, L.D. Lethal dominant roan in horses. J. Hered. 1979, 70, 145–146. [Google Scholar] [CrossRef]
- Marklund, S.; Moller, M.; Sandberg, K.; Andersson, L. Close association between sequence polymorphism in the KIT gene and the roan coat color in horses. Mamm. Genome 1999, 10, 283–288. [Google Scholar] [CrossRef]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.-S.; et al. Coat color variation at the beginning of horse domestication. Science 2009, 324, 485. [Google Scholar] [CrossRef]
- Stachurska, A.; Jansen, P. Crypto-tobiano horses in Hucul breed. Czech J. Anim. Sci. 2015, 60, 1–9. [Google Scholar] [CrossRef]
- Kawakami, T.; Jensen, M.K.; Slavney, A.; Deane, P.E.; Milano, A.; Raghavan, V.; Ford, B.; Chu, E.T.; Sams, A.J.; Boyko, A.R. R-locus for roaned coat is associated with a tandem duplication in an intronic region of USH2A in dogs and also contributes to Dalmatian spotting. PLoS ONE 2021, 16, e0248233. [Google Scholar] [CrossRef]
- Bellone, R.R.; Tanaka, J.; Esdaile, E.; Sutton, R.B.; Payette, F.; Leduc, L.; Till, B.J.; Abdel-Ghaffar, A.K.; Hammond, M.; Magdesian, K.G. A de novo 2.3 kb structural variant in MITF explains a novel splashed white phenotype in a Thoroughbred family. Anim. Genet. 2023, 54, 752–762. [Google Scholar] [CrossRef]
- Hauswirth, R.; Haase, B.; Blatter, M.; Brooks, S.A.; Burger, D.; Drögemüller, C.; Gerber, V.; Henke, D.; Janda, J.; Jude, R.; et al. Mutations in MITF and PAX3 Cause “Splashed White” and Other White Spotting Phenotypes in Horses. PLoS Genet. 2012, 8, e1002653. [Google Scholar] [CrossRef]
- Henkel, J.; Lafayette, C.; Brooks, S.A.; Martin, K.; Patterson-Rosa, L.; Cook, D.; Jagannathan, V.; Leeb, T. Whole-genome sequencing reveals a large deletion in the MITF gene in horses with white spotted coat colour and increased risk of deafness. Anim. Genet. 2019, 50, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Magdesian, K.G.; Tanaka, J.; Bellone, R.R. A De Novo MITF Deletion Explains a Novel Splashed White Phenotype in an American Paint Horse. J. Hered. 2020, 111, 287–293. [Google Scholar] [CrossRef]
- McFadden, A.; Martin, K.; Foster, G.; Vierra, M.; Lundquist, E.W.; Everts, R.E.; Martin, E.; Volz, E.; McLoone, K.; Brooks, S.A.; et al. Two Novel Variants in MITF and PAX3 Associated with Splashed White Phenotypes in Horses. J. Equine Vet. Sci. 2023, 128, 104875. [Google Scholar] [CrossRef]
- Patterson Rosa, L.; Martin, K.; Vierra, M.; Foster, G.; Brooks, S.A.; Lafayette, C. Non-frameshift deletion on MITF is associated with a novel splashed white spotting pattern in horses (Equus caballus). Anim. Genet. 2022, 53, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Galibert, M.-D.; Dexter, T.J.; Goding, C.R.; Yavuzer, U. Pax3 and Regulation of the Melanocyte-specific Tyrosinase-related Protein-1 Promoter. J. Biol. Chem. 1999, 274, 26894–26900. [Google Scholar] [CrossRef]
- Pogenberg, V.; Ögmundsdóttir, M.H.; Bergsteinsdóttir, K.; Schepsky, A.; Phung, B.; Deineko, V.; Milewski, M.; Steingrímsson, E.; Wilmanns, M. Restricted leucine zipper dimerization and specificity of DNA recognition of the melanocyte master regulator MITF. Genes Dev. 2012, 26, 2647–2658. [Google Scholar] [CrossRef] [PubMed]
- Dürig, N.; Jude, R.; Jagannathan, V.; Leeb, T. A novel MITF variant in a white American Standardbred foal. Anim. Genet. 2017, 48, 123–124. [Google Scholar] [CrossRef]
- Negro, S.; Imsland, F.; Valera, M.; Molina, A.; Solé, M.; Andersson, L. Association analysis of KIT, MITF, and PAX3 variants with white markings in Spanish horses. Anim. Genet. 2017, 48, 349–352. [Google Scholar] [CrossRef]
- Scott, M.P.; Tamkun, J.W.; Hartzell, G.W. The structure and function of the homeodomain. Biochim. Biophys. Acta (BBA)-Rev. Cancer 1989, 989, 25–48. [Google Scholar] [CrossRef]
- Thompson, B.; Davidson, E.A.; Liu, W.; Nebert, D.W.; Bruford, E.A.; Zhao, H.; Dermitzakis, E.T.; Thompson, D.C.; Vasiliou, V. Overview of PAX gene family: Analysis of human tissue-specific variant expression and involvement in human disease. Hum. Genet. 2021, 140, 381–400. [Google Scholar] [CrossRef]
- Goulding, M.D.; Chalepakis, G.; Deutsch, U.; Erselius, J.R.; Gruss, P. Pax-3, a novel murine DNA binding protein expressed during early neurogenesis. EMBO J. 1991, 10, 1135–1147. [Google Scholar] [CrossRef]
- McFadden, A.; Martin, K.; Vierra, M.; Robilliard, H.; Lundquist, E.W.; Everts, R.E.; Brooks, S.A.; Lafayette, C. Three HPS5 Mutations Associated with Depigmentation in Diverse Horse Breeds. J. Livest. Sci. 2023; Manuscript submitted for review. [Google Scholar]
- Santschi, E.M.; Vrotsos, P.D.; Purdy, A.K.; Mickelson, J.R. Incidence of the endothelin receptor B mutation that causes lethal white foal syndrome in white-patterned horses. Am. J. Vet. Res. 2001, 62, 97–103. [Google Scholar] [CrossRef]
- Santschi, E.M.; Purdy, A.K.; Valberg, S.J.; Vrotsos, P.D.; Kaese, H.; Mickelson, J.R. Endothelin receptor B polymorphism associated with lethal white foal syndrome in horses. Mamm. Genome 1998, 9, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Holl, H.M.; Brooks, S.A.; Archer, S.; Brown, K.; Malvick, J.; Penedo, M.C.T.; Bellone, R.R. Variant in the RFWD3 gene associated with PATN1, a modifier of leopard complex spotting. Anim. Genet. 2016, 47, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, H.; Mack, M.; Famula, T.; Sandmeyer, L.; Bauer, B.; Dwyer, A.; Lassaline, M.; Beeson, S.; Archer, S.; McCue, M.; et al. Genetic investigation of equine recurrent uveitis in Appaloosa horses. Anim. Genet. 2020, 51, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Sandmeyer, L.S.; Kingsley, N.B.; Walder, C.; Archer, S.; Leis, M.L.; Bellone, R.R.; Bauer, B.S. Risk factors for equine recurrent uveitis in a population of Appaloosa horses in western Canada. Vet. Ophthalmol. 2020, 23, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Esdaile, E.S. Short Tandem Repeat Analysis of Genetic Diversity Metrics in American Standardbreds and an Investigation on the Cause of the Rabicano Coat Color Phenotype Utilizing Short and Long Read Sequencing Methods. [UC Davis]. 2021. Available online: https://escholarship.org/uc/item/95w0q51r (accessed on 17 July 2023).
Figure 1.
Various phenotypes caused by variants at the W locus, including full depigmentation, a sabino-like pattern, and minimal markings. Coat color genotypes and breeds are as follows: (A) a/a E/e (black) W13/n, Friesian-American White Horse cross. (B) A/a E/e Cr/n (Buckskin). (C) Foal—A/A E/E (bay) W15/15, Arabian, Dam—A/A E/e (bay) W15/n, Arabian.
Figure 1.
Various phenotypes caused by variants at the W locus, including full depigmentation, a sabino-like pattern, and minimal markings. Coat color genotypes and breeds are as follows: (A) a/a E/e (black) W13/n, Friesian-American White Horse cross. (B) A/a E/e Cr/n (Buckskin). (C) Foal—A/A E/E (bay) W15/15, Arabian, Dam—A/A E/e (bay) W15/n, Arabian.

Figure 2.
Varying degrees of the tobiano phenotype in the Hucul horse. The horse in (A) displays cryptic tobiano markings that can be confused with a solid horse with white markings despite carrying a TO variant. The horse in (B) shows minimal tobiano markings. The horse in (C) displays classic tobiano. All horses possess at least one TO variant. All horses display their base coat color as well as white markings. Figure adapted from [24]. Photographs by M. Pasternak.
Figure 2.
Varying degrees of the tobiano phenotype in the Hucul horse. The horse in (A) displays cryptic tobiano markings that can be confused with a solid horse with white markings despite carrying a TO variant. The horse in (B) shows minimal tobiano markings. The horse in (C) displays classic tobiano. All horses possess at least one TO variant. All horses display their base coat color as well as white markings. Figure adapted from [24]. Photographs by M. Pasternak.

Figure 3.
Incomplete dominance of the sabino trait. The American Saddlebred horse in (A) genotypes as SB1/sb1 and displays white socks, a blaze, and white spots stretching across the body while the Paint horse foal in (B) genotypes as SB1/SB1 and displays an all-white phenotype. Pink skin is exposed on the faces of both individuals.
Figure 3.
Incomplete dominance of the sabino trait. The American Saddlebred horse in (A) genotypes as SB1/sb1 and displays white socks, a blaze, and white spots stretching across the body while the Paint horse foal in (B) genotypes as SB1/SB1 and displays an all-white phenotype. Pink skin is exposed on the faces of both individuals.
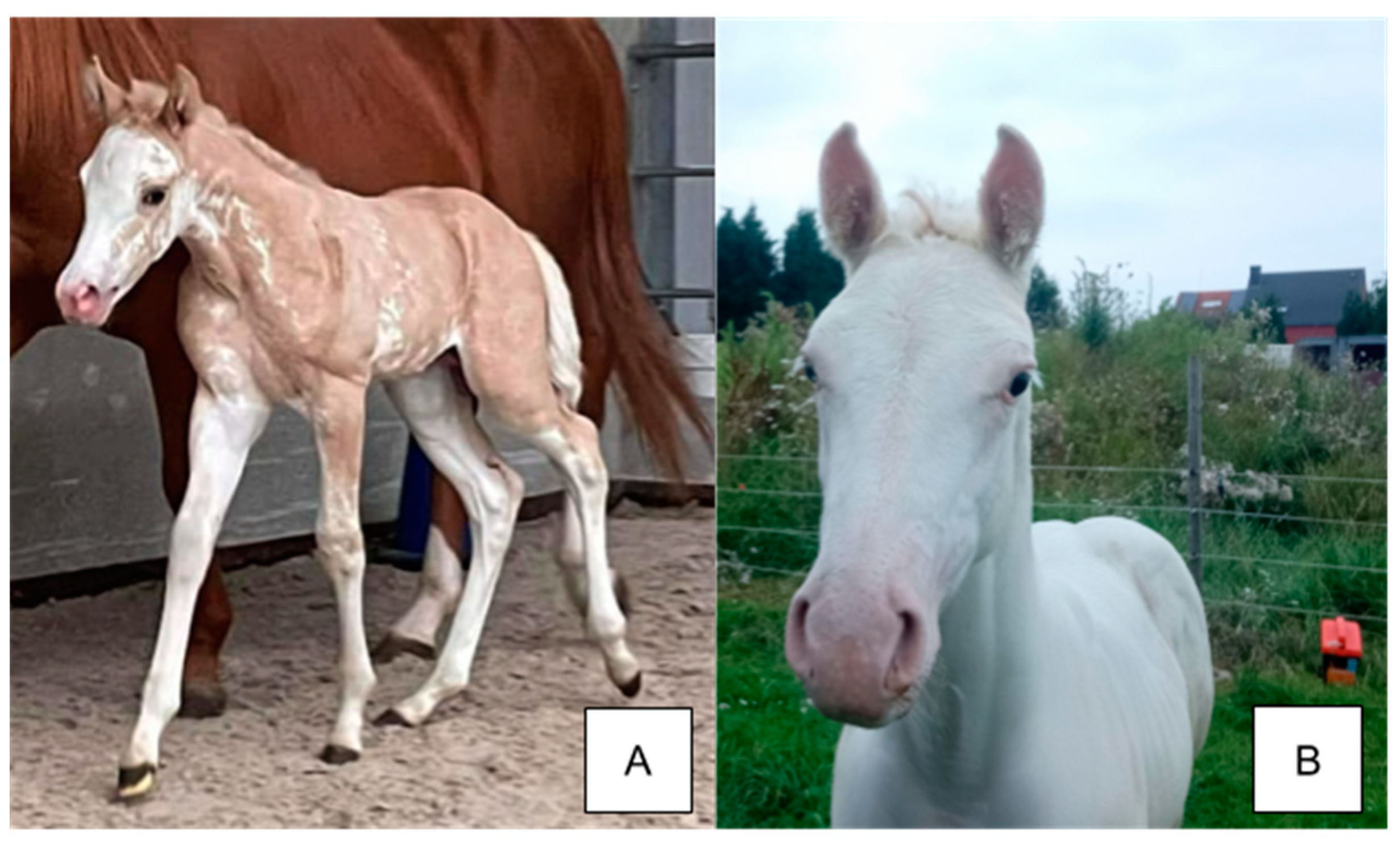
Figure 4.
(A) Chestnut Roan Quarter horse with a mixture of white and chestnut hairs over the body (B) Bay Roan Quarter horse with a mixture of white and dark hairs over the body. The head and lower legs are solid in color for both horses.
Figure 4.
(A) Chestnut Roan Quarter horse with a mixture of white and chestnut hairs over the body (B) Bay Roan Quarter horse with a mixture of white and dark hairs over the body. The head and lower legs are solid in color for both horses.

Figure 5.
Common splashed white markings. (A) Paint horse with SW7 displaying typical splashed markings on the face, legs, and body and blue eyes. This individual was reported as deaf and possesses one Cream variant diluting the coat color. Image adapted from [48]. (B) Pura Raza Española horse with SW9 displaying blue eyes, four white limbs, and a white face. (C) Franches-Montagnes horse with the Macchiato variant, which causes a phenotype similar to splashed white and a coat color dilution similar to Cream. This individual was reported to not carry any coat dilutions other than Macchiato. Image adapted from [44].
Figure 5.
Common splashed white markings. (A) Paint horse with SW7 displaying typical splashed markings on the face, legs, and body and blue eyes. This individual was reported as deaf and possesses one Cream variant diluting the coat color. Image adapted from [48]. (B) Pura Raza Española horse with SW9 displaying blue eyes, four white limbs, and a white face. (C) Franches-Montagnes horse with the Macchiato variant, which causes a phenotype similar to splashed white and a coat color dilution similar to Cream. This individual was reported to not carry any coat dilutions other than Macchiato. Image adapted from [44].

Figure 6.
Phenotypes of horses with EDXW1-3. (A) Shire horse homozygous for both EDXW1 and EDXW3 displaying white spotting with white spots on the abdomen and blue eyes. (B) A Gypsy Vanner horse heterozygous for EDXW2 showing a similar, but less extensive, phenotype as Horse A. There is slight depigmentation below the back quarter. (C) Shire horse heterozygous for EDXW3 displaying white spotting on the abdomen, legs, and face. The horses pictured possess no white-causing variants except for the alleles explicitly mentioned.
Figure 6.
Phenotypes of horses with EDXW1-3. (A) Shire horse homozygous for both EDXW1 and EDXW3 displaying white spotting with white spots on the abdomen and blue eyes. (B) A Gypsy Vanner horse heterozygous for EDXW2 showing a similar, but less extensive, phenotype as Horse A. There is slight depigmentation below the back quarter. (C) Shire horse heterozygous for EDXW3 displaying white spotting on the abdomen, legs, and face. The horses pictured possess no white-causing variants except for the alleles explicitly mentioned.

Figure 7.
Common phenotypes of Paint horses with Lethal White Overo (LWO). (A) LWO/n individual displays a blue eye and white blaze (photograph by M. Simmons). (B) Jagged body marks as a result of one copy of LWO. (C) All white homozygous LWO foal and a heterozygous LWO mare. The photographs in (B,C) were adapted from [57]).
Figure 7.
Common phenotypes of Paint horses with Lethal White Overo (LWO). (A) LWO/n individual displays a blue eye and white blaze (photograph by M. Simmons). (B) Jagged body marks as a result of one copy of LWO. (C) All white homozygous LWO foal and a heterozygous LWO mare. The photographs in (B,C) were adapted from [57]).

Figure 8.
Phenotypes of Appaloosa horses attributed to LP and PATN1. (A) LP/lp patn1/patn1 individual with a depigmented hind quarter. (B) LP/LP patn1/patn1 individual with a depigmented flank and the few spot pattern. (C) LP/lp PATN1/patn1 individual with full leopard spotting. (D) LP/LP PATN1/PATN1 individual that is fully white with no other white spotting alleles.
Figure 8.
Phenotypes of Appaloosa horses attributed to LP and PATN1. (A) LP/lp patn1/patn1 individual with a depigmented hind quarter. (B) LP/LP patn1/patn1 individual with a depigmented flank and the few spot pattern. (C) LP/lp PATN1/patn1 individual with full leopard spotting. (D) LP/LP PATN1/PATN1 individual that is fully white with no other white spotting alleles.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
White coat color traits and their distinguishing characteristics along with the associated genes and encoded proteins.
Table 1.
White coat color traits and their distinguishing characteristics along with the associated genes and encoded proteins.
| Trait | Allele Symbol | Gene/Region | Encoded Protein (Complex) | Distinguishing Characteristics (Phenotype) |
|---|---|---|---|---|
| Dominant White | W | KIT | KIT | White spots or white coat, pink skin (sabino-like, all white) |
| Tobiano | TO | KIT | KIT | Large white spots covering the body and crossing the spine, white legs, pink skin (tobiano) |
| Sabino | SB1 | KIT | KIT | Jagged white markings or white coat, pink skin (sabino, all white) |
| Roan | RN | KIT | KIT | Interspersed white hairs distributed through the coat (roan) |
| Splashed White | SW | MITF, PAX3 | MITF, PAX3 | Deafness, blue eyes, smooth boarded white faces, abdomens, and legs (splashed white) |
| Eden White | EDXW | HPS5 | HPS5 (BLOC-2) | White body spots, legs, and faces (sabino-like) |
| Lethal White Overo | LWO, O | EDNRB | EDNRB | White face, legs, and body spots that do not cross the spine (overo) |
| Leopard Spotting | LP | TRPM1 | TRPM1 | White hind quarter or white coat with colored spots, epistatic to PATN1 (blanket or leopard) |
| Pattern 1 | PATN1 | RFWD3 | RFWD3 | Increases white spotting for individuals with a least one copy of LP |
Table 2.
Genomic location, variant type, and phenotypes of Dominant White KIT variants.
| Allele | Genomic Coordinate EquCab3.0 | Type | Phenotype | Homozygotes | References |
|---|---|---|---|---|---|
| W1 | chr3:79545942G>C | nonsense | All White | Not Observed | [2] |
| W2 | chr3:79549540C>T | missense | All White | Not Observed | [2] |
| W3 | chr3:79578535T>A | nonsense | All White | Not Observed | [2] |
| W4 | chr3:79549780G>A | missense | All White | Not Observed | [2] |
| W5 | chr3:79545900delC | small deletion | Sabino-like | Not Observed | [12] |
| W6 | chr3:79573754C>T | missense | Sabino-like to All White | Not Observed | [12] |
| W7 | chr3:79580000C>G | splice site | All White | Not Observed | [12] |
| W8 | chr3:79545374C>T | splice site | Sabino-like | Not Observed | [12] |
| W9 | chr3:79549797C>T | missense | All White | Not Observed | [12] |
| W10 | chr3:79566925_79566928del | small deletion | Sabino-like to All White | Not Observed | [12] |
| W11 | chr3:79540429C>A | splice site | All White | Not Observed | [12] |
| W12 | chr3:79579755_79579779delAGACG | small deletion | Sabino-like | Not Observed | [17] |
| W13 | chr3:79544066C>G | splice site | All White | Not Observed | [14] |
| W14 | chr3:79544151_79544204del | gross deletion | All White | Not Observed | [14] |
| W15 | chr3:79550351A>G | missense | Sabino-like to All White | Observed | [14,18] |
| W16 | chr3:79540741T>A | missense | All White | Not Observed | [14] |
| W17a | chr3:79548265T>A | missense | All White | Not Observed | [14] |
| W17b | chr3:79548244A>G | missense | All White | Not Observed | [14] |
| W18 | chr3:79553751C>T | splice site | Sabino-like | Not Observed | [15] |
| W19 | chr3:79553776T>C | missense | Sabino-like | Observed | [15] |
| W20 | chr3:7948220T>C | missense | No markings to Sabino-like | Observed | [15] |
| W21 | chr3:79544174delG | small deletion | Sabino-like | Not Observed | [13] |
| W22 | chr3:79548925_79550822del1898 | gross deletion | Sabino-like | Not Observed | [11] |
| W23 | chr3:79578484C>G | splice site | All White | Not Observed | [18] |
| W24 | chr3:79545245C>T | splice site | All White | Not Observed | [10] |
| W25 | chr3:77769878T>C | missense | All White | Not Observed | [16] |
| W26 | chr3:79544150del | small deletion | Sabino-like | Not Observed | [16] |
| W27 | chr3:79552028A>C | missense | All White | Not Observed | [16] |
| W28 | chr3:79579925_79581197del | gross deletion | Sabino-like | Not Observed | [19] |
| W30 | chr3:79548244T>A | missense | All White | Not Observed | [20] |
| W31 | chr3:79618532_79618533insT | fs nonsense | Sabino-like | Not Observed | [7] |
| W32 | chr3:79538738C>T | missense | No markings to Sabino-like | Observed | [7] |
| W33 | chr3:79545248T>A | missense | Sabino-like | Not Observed | [5] |
| W34 | chr3:79566881T>C | missense | No markings to Sabino-like | Observed | [8] |
| W35 | chr3:79618649A>C | UTR variant | No markings to Sabino-like | Observed | [6] |
Table 3.
Genomic coordinates, gene, variant type, and phenotypes of Tobiano and Sabino variants and the Roan-associated SNP.
Table 3.
Genomic coordinates, gene, variant type, and phenotypes of Tobiano and Sabino variants and the Roan-associated SNP.
| Allele | Gene | Genome Coordinate EquCab3.0 | Type | Phenotype | Reference |
|---|---|---|---|---|---|
| SB1 | KIT | chr3:79544206A>T | splice site | White markings with jagged borders and pink skin. Homozygotes are all white with pink skin | [9] |
| TO | Intergenic | chr3:42737476_79471993inv | paracentric inversion | Large white areas across the body, pink skin | [36] |
| RN | KIT | chr3:79543439A>G | associated SNP | White hairs distributed throughout the coat | [37] |
Table 4.
Genomic coordinates, gene, variant type, and phenotypes of Splashed White variants.
| Allele | Gene | Genomic Coordinate EquCab3.0 | Type | Phenotype | Reference |
|---|---|---|---|---|---|
| SW1 | MITF | chr16:21579201delinsATAATAACCTA | small indel | Splashed Markings, Deafness, Blue Eyes | [44] |
| SW2 | PAX3 | chr6:11199026C>T | missense | Minimal Splashed Markings, Deafness, Blue Eyes | [44] |
| SW3 | MITF | chr16:21567245_21567249del | small deletion | Splashed Markings, Deafness, Blue Eyes | [44] |
| SW4 | PAX3 | chr6:11199140G>C | missense | Minimal Splashed Markings, Blue Eyes | [15] |
| SW5 | MITF | chr16:21503211_21566617del | gross deletion | Splashed Markings, Deafness, Blue Eyes | [45] |
| SW6 | MITF | chr16:21551060_21559770del | gross deletion | Splashed Markings, Deafness, Blue Eyes | [46] |
| SW7 | MITF | chr16:21559953_21559955del | small deletion | Splashed Markings, Deafness, Blue Eyes | [48] |
| SW8 | MITF | chr16:21555811_21558139del | gross deletion | Splashed Markings, Deafness, Blue Eyes | [43] |
| SW9 | MITF | chr16:21559940A>T | missense | Splashed Markings, Blue Eyes | [47] |
| SW10 | PAX3 | chr6:11196181C>T | nonsense | Splashed Markings, Blue Eyes | [47] |
Table 5.
Genomic coordinates, variant type, and phenotypes of Eden White variants in HPS5.
| Allele | Genomic Coordinate EquCab3.0 | Type | Phenotype | Reference |
|---|---|---|---|---|
| EDXW1 | chr7:88741616T>A | missense | High socks, belly spots, blazes | [56] |
| EDXW2 | chr7:88751667G>A | missense | High socks, belly spots, blazes | [56] |
| EDXW3 | chr7:88766673T>C | splice site | High socks, belly spots, blazes | [56] |
Table 6.
Genomic coordinates, genes, variant type, and phenotypes of LP and PATN1.
| Allele | Gene | Genomic Coordinate EquCab3.0 | Type | Phenotype | Reference |
|---|---|---|---|---|---|
| LP | TRPM1 | chr1:108297929_108297930insN[1378] | insertion | White hind quarter, all white (hom), leopard spots | [27] |
| PATN1 | RFWD3 | chr3:24352525T>G | UTR variant | Increases white and decreases number of spots only for LP/LP or LP/lp individuals | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McFadden, A.; Vierra, M.; Martin, K.; Brooks, S.A.; Everts, R.E.; Lafayette, C. Spotting the Pattern: A Review on White Coat Color in the Domestic Horse. Animals 2024, 14, 451. https://doi.org/10.3390/ani14030451
AMA Style
McFadden A, Vierra M, Martin K, Brooks SA, Everts RE, Lafayette C. Spotting the Pattern: A Review on White Coat Color in the Domestic Horse. Animals. 2024; 14(3):451. https://doi.org/10.3390/ani14030451
Chicago/Turabian StyleMcFadden, Aiden, Micaela Vierra, Katie Martin, Samantha A. Brooks, Robin E. Everts, and Christa Lafayette. 2024. "Spotting the Pattern: A Review on White Coat Color in the Domestic Horse" Animals 14, no. 3: 451. https://doi.org/10.3390/ani14030451
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.